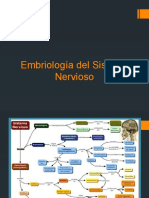
También podría gustarte
- Histología y Embrología del Sitema NerviosoDe EverandHistología y Embrología del Sitema NerviosoCalificación: 4 de 5 estrellas4/5 (4)
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- Tarea 1 de PsicofisiologiaDocumento16 páginasTarea 1 de Psicofisiologialaura michelleAún no hay calificaciones
- Guía de Embriología Del Sistema NerviosoDocumento23 páginasGuía de Embriología Del Sistema Nerviosoprotoprueba123Aún no hay calificaciones
- Resumen NeuroDocumento91 páginasResumen NeuroNa ParritaAún no hay calificaciones
- Sistema nervioso: Definición, estructura y desarrollo embriológicoDocumento15 páginasSistema nervioso: Definición, estructura y desarrollo embriológicoElexnat IdiomasAún no hay calificaciones
- Embriología Del Sistema NerviosoDocumento9 páginasEmbriología Del Sistema NerviosoBETSY ANDREA CHUQUIN GARAYAún no hay calificaciones
- Desarrollo Embrionario y Neurobiologia de La Neurona y NeurogliaDocumento6 páginasDesarrollo Embrionario y Neurobiologia de La Neurona y NeurogliaLisbeth Alejandra Martínez GonzálezAún no hay calificaciones
- MonofrafíaDocumento12 páginasMonofrafíaLucero Palomino RomeroAún no hay calificaciones
- Primera Parte EmbriologiaDocumento30 páginasPrimera Parte EmbriologiamiguelAún no hay calificaciones
- NEURULACIÓNDocumento4 páginasNEURULACIÓNAbraham SánchezAún no hay calificaciones
- Desarrollo de Sistema NerviosoDocumento8 páginasDesarrollo de Sistema NerviosoSebastián Andrés Urrea GonzalezAún no hay calificaciones
- 10 Segundo SISTEMA NERVIOSO 1Documento61 páginas10 Segundo SISTEMA NERVIOSO 1elpinchosblogAún no hay calificaciones
- 13 Na 1Documento22 páginas13 Na 1Sanatú PsicoterapiaAún no hay calificaciones
- Guia 5Documento4 páginasGuia 5Ana Belén MedinaAún no hay calificaciones
- Investigación de Nuerologia Con APADocumento81 páginasInvestigación de Nuerologia Con APAEvelyn Edith Zetino Garc�aAún no hay calificaciones
- Sistema Nervioso Central EmbrioDocumento16 páginasSistema Nervioso Central EmbrioRicardo Molina OrtizAún no hay calificaciones
- Embriologia Del Sistema Nervioso CentralDocumento24 páginasEmbriologia Del Sistema Nervioso CentralGELEXSAún no hay calificaciones
- Informe NeurocienciasDocumento8 páginasInforme NeurocienciasCésar AntonioAún no hay calificaciones
- Embriología del SN enDocumento26 páginasEmbriología del SN enAngeles Cifuentes Caceres100% (1)
- Desarrollo SNC y órganos sentidosDocumento5 páginasDesarrollo SNC y órganos sentidosmv glAún no hay calificaciones
- Sistema nervioso: desarrollo del cerebro, médula espinal y ganglios raquídeosDocumento13 páginasSistema nervioso: desarrollo del cerebro, médula espinal y ganglios raquídeosTodoMePuedeAlvAún no hay calificaciones
- Gould, D. (2015) - Neuroanatomía (5th Edition Ed.) y Purves de NeurocienciaDocumento5 páginasGould, D. (2015) - Neuroanatomía (5th Edition Ed.) y Purves de NeurocienciaFelipe GuzmanAún no hay calificaciones
- Trastornos AsociadosDocumento46 páginasTrastornos AsociadosJuan Diego Umaña AguilarAún no hay calificaciones
- Sistema nervioso centralDocumento7 páginasSistema nervioso centralGreysmar Vanessa Chirinos GuapeAún no hay calificaciones
- Transcripcion Desarrollo Del Sistema NerviosoDocumento9 páginasTranscripcion Desarrollo Del Sistema NerviosoValentina EcAún no hay calificaciones
- Formación tubo neuralDocumento16 páginasFormación tubo neuralMARLYN MATAAún no hay calificaciones
- Neurología Columna VertebralDocumento58 páginasNeurología Columna VertebraldianaAún no hay calificaciones
- Embriologia Del SNC - Filogenia y OntogeniaDocumento19 páginasEmbriologia Del SNC - Filogenia y OntogeniaSlim InfinitoAún no hay calificaciones
- Desarrollo SNC desde gastrulación a formación cerebroDocumento28 páginasDesarrollo SNC desde gastrulación a formación cerebroKaterine BurbanoAún no hay calificaciones
- Resumen Del Sistema NerviosoDocumento5 páginasResumen Del Sistema NerviosoAtenea 03Aún no hay calificaciones
- Desarrollo Del Sistema Nervioso Central EmbriologíaDocumento3 páginasDesarrollo Del Sistema Nervioso Central EmbriologíaKaterine BurbanoAún no hay calificaciones
- Sistema Nervioso EmbrioDocumento7 páginasSistema Nervioso EmbrioRico tovar Fernanda YezabelAún no hay calificaciones
- Desarrollo Embriologico Del Sist NerviosoDocumento31 páginasDesarrollo Embriologico Del Sist NerviosoMagali Ramirez MonterAún no hay calificaciones
- Embriología Del Sistema NerviosoDocumento25 páginasEmbriología Del Sistema NerviosoJaime Miranda Mendez100% (2)
- Desarrollo Embrionario SISTEMA NERVIOSO-1Documento70 páginasDesarrollo Embrionario SISTEMA NERVIOSO-1Brenda Gress ViverosAún no hay calificaciones
- Fases Del DesarrolloDocumento32 páginasFases Del DesarrollomariaAún no hay calificaciones
- EmbriologiaDocumento22 páginasEmbriologiaEdisson Morales100% (1)
- Sistema nervioso central 2.0Documento5 páginasSistema nervioso central 2.0Jorge Chz subsAún no hay calificaciones
- EmbriologiaDocumento5 páginasEmbriologiaFrank ReyesAún no hay calificaciones
- HDocumento10 páginasHGeorge BetancourthAún no hay calificaciones
- Sistema Nervioso PDFDocumento18 páginasSistema Nervioso PDFJOCELYN FIERRO GUERREROAún no hay calificaciones
- NEURULACIONDocumento12 páginasNEURULACIONDulce Rayo LealAún no hay calificaciones
- Desarrollo del Sistema Nervioso CentralDocumento8 páginasDesarrollo del Sistema Nervioso CentralDiego Bedón AscurraAún no hay calificaciones
- Manual de Neuro Dr. GaribayDocumento120 páginasManual de Neuro Dr. GaribayCarla A. CorriloAún no hay calificaciones
- Embriología Del Sistema NerviosoDocumento104 páginasEmbriología Del Sistema NerviosoDaniel Sebastian100% (2)
- Clase 3 EmbriologiaDocumento32 páginasClase 3 EmbriologiaMarcial Hernández MercadoAún no hay calificaciones
- Tarea 1Documento23 páginasTarea 1Loanyely MorilloAún no hay calificaciones
- Embriología Del SNCDocumento5 páginasEmbriología Del SNCManuel Alejandro Parada100% (1)
- Desarrollo Embrionario PDFDocumento11 páginasDesarrollo Embrionario PDFMoises Skazas Neuronas100% (1)
- Catedra de Laboratorio de Neuroanatomía Embriologia e HistogenesisDocumento8 páginasCatedra de Laboratorio de Neuroanatomía Embriologia e HistogenesisGregory DislaAún no hay calificaciones
- Nervioso CKQDocumento33 páginasNervioso CKQMatty MarchandAún no hay calificaciones
- Clases de NeuroDocumento221 páginasClases de NeuroPaul StewartAún no hay calificaciones
- Sistema Nervioso - Embriologìa - UNPRG - 2017 - Dr. Jorge OrtizDocumento84 páginasSistema Nervioso - Embriologìa - UNPRG - 2017 - Dr. Jorge Ortizruben castroAún no hay calificaciones
- NEUROLOGÍA CompletoDocumento124 páginasNEUROLOGÍA CompletoAmanita HernandezAún no hay calificaciones
- Anatomia y Fisiología del Sistema Nervioso IIDe EverandAnatomia y Fisiología del Sistema Nervioso IICalificación: 4 de 5 estrellas4/5 (28)
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2De EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Aún no hay calificaciones
- El cannabis en patologías del sistema nervioso centralDe EverandEl cannabis en patologías del sistema nervioso centralAún no hay calificaciones
- Cómo se comunican las neuronas: El milagro de la transmisión sinápticaDe EverandCómo se comunican las neuronas: El milagro de la transmisión sinápticaAún no hay calificaciones
- Diagramas ProcesoDocumento64 páginasDiagramas ProcesoIker RV100% (1)
- 01-Teorias Del AprendizajeDocumento21 páginas01-Teorias Del AprendizajeCristián EcheverríaAún no hay calificaciones
- Es - yDocumento647 páginasEs - yChihiro NekotaAún no hay calificaciones
- Poemas para El Día de La Madre - WWW - Mundodeportivo.com - Uncomo PDFDocumento5 páginasPoemas para El Día de La Madre - WWW - Mundodeportivo.com - Uncomo PDFDamris Naghel Jimenez AlfaroAún no hay calificaciones
- InyectologíaDocumento5 páginasInyectologíaVivi ViviAún no hay calificaciones
- Caida LibreDocumento5 páginasCaida LibreKAREN DAYANA MESA NUÑEZAún no hay calificaciones
- Plantilla Excel Punto de Equilibrio MORATIVADocumento6 páginasPlantilla Excel Punto de Equilibrio MORATIVAJulio James ZeaAún no hay calificaciones
- Inversión extranjera Honduras 2017 crecerá 6Documento4 páginasInversión extranjera Honduras 2017 crecerá 6lopezmoisesAún no hay calificaciones
- Diagrama de Nyquist para evaluar estabilidadDocumento4 páginasDiagrama de Nyquist para evaluar estabilidadKevin Ascue ContrerasAún no hay calificaciones
- Gastronomía Peruana: Ingredientes ProtagonistasDocumento48 páginasGastronomía Peruana: Ingredientes ProtagonistasAlex Huallpa IncaAún no hay calificaciones
- Leccion 1. DESARROLLO SOSTENIBLE PDFDocumento5 páginasLeccion 1. DESARROLLO SOSTENIBLE PDFFabian PerillaAún no hay calificaciones
- NTC4403Documento9 páginasNTC4403LUIS FERNANDO MUÑOZ DE ARCOSAún no hay calificaciones
- Acuerdo MDT-2015-0262 Ecuador - TransportistasDocumento7 páginasAcuerdo MDT-2015-0262 Ecuador - TransportistasStefany RodriguezAún no hay calificaciones
- Reporte 1 Modelado MolecularDocumento3 páginasReporte 1 Modelado MolecularAleida Sugey Ovalle GarcíaAún no hay calificaciones
- Análisis del estudio de pronósticos de ventas de Mentzer y su aplicación en LatinoaméricaDocumento5 páginasAnálisis del estudio de pronósticos de ventas de Mentzer y su aplicación en LatinoaméricaJhon Cubas CalderonAún no hay calificaciones
- Tema 6 - Instalacion de Redes Inalambricas y VSAT - Introduccion para AlumnadoDocumento3 páginasTema 6 - Instalacion de Redes Inalambricas y VSAT - Introduccion para Alumnadoperiquito pinpinAún no hay calificaciones
- Roncadora CajamarcaDocumento13 páginasRoncadora CajamarcaAlessandra herrera gongoraAún no hay calificaciones
- Rodrigo - Antonio - Mota - Martínez - U4 - A2 - Pronostico de La Demanda - Pag - 506-511Documento11 páginasRodrigo - Antonio - Mota - Martínez - U4 - A2 - Pronostico de La Demanda - Pag - 506-511Rodrigo Antonio Mota MartinezAún no hay calificaciones
- Ept - Agropecuaria - Pucayacu IIDocumento32 páginasEpt - Agropecuaria - Pucayacu IIliguis4pinedo4esquivAún no hay calificaciones
- AISI-304 y AISI-316 resistencia corrosión agentesDocumento3 páginasAISI-304 y AISI-316 resistencia corrosión agentesJuan Carlos Marquez MoralesAún no hay calificaciones
- Reglamento Sobre Prevencion de IncendiosDocumento0 páginasReglamento Sobre Prevencion de IncendiosYimmy VelasquezAún no hay calificaciones
- Desinfectante concentrado industrial CitraKlinDocumento2 páginasDesinfectante concentrado industrial CitraKlinYOLANDA BUELNAAún no hay calificaciones
- Estabilidad Criterios Routh HurwitzDocumento20 páginasEstabilidad Criterios Routh HurwitzFernando Vasquez CornejoAún no hay calificaciones
- Evidencia de Tarea 4 OSMORREGULACIONDocumento16 páginasEvidencia de Tarea 4 OSMORREGULACIONCardenas Stella100% (1)
- Instrumento de Valoración 6to GradoDocumento4 páginasInstrumento de Valoración 6to GradoElizabeth Francisco HernándezAún no hay calificaciones
- 7.2. 3 Botín Dieléctrico Nazca Borazón Nu-615 - Ficha TécnicaDocumento1 página7.2. 3 Botín Dieléctrico Nazca Borazón Nu-615 - Ficha TécnicaDennis GoyaAún no hay calificaciones
- 001-PTS - Instalación de Inclinómetros - Exploración Profunda ENGIE-SIERRA (Rev RGG)Documento15 páginas001-PTS - Instalación de Inclinómetros - Exploración Profunda ENGIE-SIERRA (Rev RGG)Anonymous QZdKvyAún no hay calificaciones
- Solidos amorfos: estructura y aplicacionesDocumento10 páginasSolidos amorfos: estructura y aplicacionesJuan ReyesAún no hay calificaciones
- TEMPERATURADocumento22 páginasTEMPERATURARonald Jhonn Condori FloresAún no hay calificaciones
- PEUGEOT EXPERT 2001-2002 Usuario EsDocumento148 páginasPEUGEOT EXPERT 2001-2002 Usuario Esgau3koAún no hay calificaciones