También podría gustarte
- Informe - Procesual 3 - Tercera Semana de DesarrolloDocumento9 páginasInforme - Procesual 3 - Tercera Semana de DesarrolloDaniela TrigoAún no hay calificaciones
- Resumen Libro Osificación ImágenesDocumento22 páginasResumen Libro Osificación ImágenesJulian OchoaAún no hay calificaciones
- Periodo Somitico 1 Vladimir FloresDocumento16 páginasPeriodo Somitico 1 Vladimir FloresChistian DavidAún no hay calificaciones
- Sistema Musculo-EsqueleticoDocumento10 páginasSistema Musculo-EsqueleticoII-D Melissa Hernandez MedinaAún no hay calificaciones
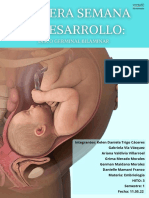
- Tercera SemanaDocumento8 páginasTercera Semanalarapaloma698Aún no hay calificaciones
- Crestas Neurales Embriologia PDFDocumento17 páginasCrestas Neurales Embriologia PDFLuis Gerardo OlivaAún no hay calificaciones
- Nervio FacialDocumento18 páginasNervio FacialAguus CabreraAún no hay calificaciones
- Informe de Biología Del DesarrolloDocumento14 páginasInforme de Biología Del DesarrolloOlaznog Serolf SelaromAún no hay calificaciones
- Tema 5. Aparato Locomotor (Embriología)Documento3 páginasTema 5. Aparato Locomotor (Embriología)Mercedes MartínAún no hay calificaciones
- Malformaciones Del Sistema Músculo Esquelético y Del Sistema TegumentarioDocumento11 páginasMalformaciones Del Sistema Músculo Esquelético y Del Sistema TegumentarioDaniel SeclenAún no hay calificaciones
- Neural Crest 1 EspañolDocumento10 páginasNeural Crest 1 EspañolSamuel AlvarezAún no hay calificaciones
- Trilaminar ResumenDocumento8 páginasTrilaminar ResumenJonathan ClementeAún no hay calificaciones
- UT3-Seminario Gastrulación SEM03Documento39 páginasUT3-Seminario Gastrulación SEM03Lauti GomezAún no hay calificaciones
- Cara Cuello Ojo y OidoDocumento27 páginasCara Cuello Ojo y OidoJimena GuerrerosAún no hay calificaciones
- Conocimiento Del Desarrollo Evolutivo Del CráneoDocumento17 páginasConocimiento Del Desarrollo Evolutivo Del CráneoEstefany SaldañaAún no hay calificaciones
- DMFSH CT 7Documento41 páginasDMFSH CT 7Ronald Italo Marcos DagaAún no hay calificaciones
- Resume NDocumento24 páginasResume NDINA REBECA FUENTES PÉREZAún no hay calificaciones
- BiologiaDocumento26 páginasBiologiaKarin FuentesAún no hay calificaciones
- Comparto Resumen Embriología Bloque 2pdf Versión 1 ContigoDocumento64 páginasComparto Resumen Embriología Bloque 2pdf Versión 1 ContigoFer FrAún no hay calificaciones
- Brodman AreasDocumento36 páginasBrodman Areasabi100% (1)
- Resumen Capitulo 5 LangmanDocumento7 páginasResumen Capitulo 5 LangmanDarkinfire Western100% (1)
- Anato TareaDocumento20 páginasAnato TareaSofiAún no hay calificaciones
- Disco TrilaminarDocumento28 páginasDisco TrilaminarDayanna Armendariz SaltosAún no hay calificaciones
- Cardiomiocito - Wikipedia, La Enciclopedia LibreDocumento21 páginasCardiomiocito - Wikipedia, La Enciclopedia LibreMilo MartinezAún no hay calificaciones
- Sistema Nervioso 2. Nervios Espinales - Meninges EspinalesDocumento10 páginasSistema Nervioso 2. Nervios Espinales - Meninges Espinaleseukenetavera23Aún no hay calificaciones
- Crestas Neurales, Placodas y Arcos Barnquiales, Una RevisioDocumento17 páginasCrestas Neurales, Placodas y Arcos Barnquiales, Una RevisioPilar CastiblancoAún no hay calificaciones
- Cuestionario M2.2Documento3 páginasCuestionario M2.2Hector Efrain Garcia ChavezAún no hay calificaciones
- 120 PreguntasDocumento64 páginas120 PreguntasJudith CJAún no hay calificaciones
- Embriologia LLDocumento17 páginasEmbriologia LLLESLIE LAISHA DE LA TORRE PEREZAún no hay calificaciones
- Embriología - 3ra Semana Del DesarrolloDocumento18 páginasEmbriología - 3ra Semana Del DesarrolloFernanda Valeria Gutiérrez LeónAún no hay calificaciones
- Embriología Arcos FaríngeosDocumento9 páginasEmbriología Arcos FaríngeosAbigailAún no hay calificaciones
- Taller de Embriologia IiDocumento2 páginasTaller de Embriologia Iijake lolAún no hay calificaciones
- Capitulo 5 Tercera Semana de Desarrollo Disco Germinal Trilaminar 359286 Downloable 1777082Documento5 páginasCapitulo 5 Tercera Semana de Desarrollo Disco Germinal Trilaminar 359286 Downloable 1777082Yasmin TatianaAún no hay calificaciones
- RESUMEN3erBLOQUEDocumento26 páginasRESUMEN3erBLOQUEIsadora DuarteAún no hay calificaciones
- Guías LL Parcial Equipo #8Documento40 páginasGuías LL Parcial Equipo #8Luisa SevillaAún no hay calificaciones
- Sistema MuscularDocumento7 páginasSistema MuscularGCanisAún no hay calificaciones
- Presentacion Salud Del Cuerpo Humano Ilustracion RosaDocumento40 páginasPresentacion Salud Del Cuerpo Humano Ilustracion RosaKarina BarbosaAún no hay calificaciones
- Art 33Documento2 páginasArt 33danielaeenestapc jimenezAún no hay calificaciones
- 4 Periodo SomiticoDocumento7 páginas4 Periodo SomiticoIvan Ariel MambrinAún no hay calificaciones
- Guia de AutoconceptoDocumento6 páginasGuia de AutoconceptoYeison PedrazaAún no hay calificaciones
- EmbriologiaDocumento9 páginasEmbriologiaYessenia Tasilla HuaripataAún no hay calificaciones
- CCBB CardioDocumento102 páginasCCBB CardioAntony AsllaAún no hay calificaciones
- MANUAL DE LABORATORIO MICROANATOMIA I LAB. 5 (Reparado)Documento13 páginasMANUAL DE LABORATORIO MICROANATOMIA I LAB. 5 (Reparado)Xd LolsñAún no hay calificaciones
- 7ma CLASE PERIODO ORGANOGENICO Y FETAL SABDocumento56 páginas7ma CLASE PERIODO ORGANOGENICO Y FETAL SABJuan Diego Lazarte FernandezAún no hay calificaciones
- GLIOBLASTOSDocumento27 páginasGLIOBLASTOSMarintalero100% (1)
- Formato Pract Cerebelo Marzo 2012Documento6 páginasFormato Pract Cerebelo Marzo 2012Julio PerezAún no hay calificaciones
- MonofrafíaDocumento12 páginasMonofrafíaLucero Palomino RomeroAún no hay calificaciones
- NerviosoDocumento51 páginasNerviosoMARLYN MATAAún no hay calificaciones
- Cuestionario Del Sistema Musculo Esquelético EmbriologiaDocumento5 páginasCuestionario Del Sistema Musculo Esquelético EmbriologiaDavid FloresAún no hay calificaciones
- Apuntes BIOELECTROMAGNETISMO - 1 - 2010Documento76 páginasApuntes BIOELECTROMAGNETISMO - 1 - 2010ayaloAún no hay calificaciones
- Lab 6 Manual MicroanatomiaDocumento12 páginasLab 6 Manual MicroanatomiaElmer BenavidesAún no hay calificaciones
- Apunte Embriologia UncDocumento4 páginasApunte Embriologia UnccolsaniflorenciaAún no hay calificaciones
- Guía de Estudio No 3 KDocumento6 páginasGuía de Estudio No 3 KkatherinefonsecasilvaAún no hay calificaciones
- Cuestionario Embriologia 3pDocumento8 páginasCuestionario Embriologia 3pALDAIR CANAVIRI SILESAún no hay calificaciones
- Desarrollo Embrionario Del Sistema NerviosoDocumento51 páginasDesarrollo Embrionario Del Sistema Nerviosogerardo vasquezAún no hay calificaciones
- Embriología Del Sistema Nervioso Vesículas Primarias y SecundariasDocumento7 páginasEmbriología Del Sistema Nervioso Vesículas Primarias y SecundariasXavier GuevaraAún no hay calificaciones
- pdf-12 en EsDocumento8 páginaspdf-12 en EsSamuel AlvarezAún no hay calificaciones
- The Influence of The Respiration - En.esDocumento5 páginasThe Influence of The Respiration - En.esSamuel AlvarezAún no hay calificaciones
- Neural Crest 1 EspañolDocumento10 páginasNeural Crest 1 EspañolSamuel AlvarezAún no hay calificaciones
- Evolutionoforthodonticappliances en EsDocumento12 páginasEvolutionoforthodonticappliances en EsSamuel AlvarezAún no hay calificaciones
- Presupuesto Oficial EP 29279 - AjustadoDocumento10 páginasPresupuesto Oficial EP 29279 - AjustadoSamuel AlvarezAún no hay calificaciones
- Sistema nervios-WPS OfficeDocumento7 páginasSistema nervios-WPS OfficeDavid peñaAún no hay calificaciones
- Caso Clinico Litiasis Vesicular PDFDocumento34 páginasCaso Clinico Litiasis Vesicular PDFJuan Lezama100% (1)
- QUÍAS DE VOZ CatalinaDocumento15 páginasQUÍAS DE VOZ CatalinaAngie Catalina Victoria BolañosAún no hay calificaciones
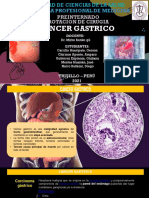
- Ca Estomago-JmhDocumento34 páginasCa Estomago-JmhAna Maria Ipanaque AlfaroAún no hay calificaciones
- P. Malignas Del Cuerpo Uterino y EndometrioDocumento36 páginasP. Malignas Del Cuerpo Uterino y EndometrioIgnacio GonzálezAún no hay calificaciones
- Medula Oblongada - Cuestionario 3Documento8 páginasMedula Oblongada - Cuestionario 3Silvia Torrico CéspedesAún no hay calificaciones
- Autónomo Biología MolecularDocumento14 páginasAutónomo Biología MolecularAnais MachadoAún no hay calificaciones
- Practica 1 BotanicaDocumento5 páginasPractica 1 BotanicaLuz Angela Tamára GuerraAún no hay calificaciones
- Tps EveaDocumento12 páginasTps Eveajsw03Aún no hay calificaciones
- Historia Clinica 3Documento8 páginasHistoria Clinica 3Carlos Toscano0% (1)
- GUIA DE ESTUDIO: Cerebro, Corteza, Hemisferios y Lóbulos Anatomía y Fisiología Del Sistema Nervioso Dr. Adrian Chávez García VALOR: 4 PuntosDocumento2 páginasGUIA DE ESTUDIO: Cerebro, Corteza, Hemisferios y Lóbulos Anatomía y Fisiología Del Sistema Nervioso Dr. Adrian Chávez García VALOR: 4 PuntosJENNIFER VERENICE GARCIA COLINDRESAún no hay calificaciones
- Dinamica UterinaDocumento24 páginasDinamica UterinaAngelAún no hay calificaciones
- RPT Consulta PDocumento2 páginasRPT Consulta PmichaelAún no hay calificaciones
- OclusionDocumento54 páginasOclusionLaura Garcia75% (4)
- Re Pa SoDocumento21 páginasRe Pa SoFainier Beita MoraAún no hay calificaciones
- PuerperioDocumento21 páginasPuerperioAbnel BalamAún no hay calificaciones
- Sistema Digestivo Invertebrado Power PointDocumento13 páginasSistema Digestivo Invertebrado Power Pointanggie100% (1)
- Esqueleto Del PatoDocumento13 páginasEsqueleto Del PatoGary Andres Oleas CevallosAún no hay calificaciones
- Wuolah Free AHG Tema 7Documento5 páginasWuolah Free AHG Tema 7mayra EscobedoAún no hay calificaciones
- Taller Animales VertebradosDocumento4 páginasTaller Animales VertebradosJessi GallegoAún no hay calificaciones
- Desarrollo Y Crecimiento: SeminariosDocumento48 páginasDesarrollo Y Crecimiento: SeminariosAndrea gAún no hay calificaciones
- Mecánica de La Ventilación PulmonarDocumento11 páginasMecánica de La Ventilación PulmonarAna janeth Avalos mataAún no hay calificaciones
- Infeccion Por ToxoplasmaDocumento19 páginasInfeccion Por ToxoplasmaMaleh BuitrónAún no hay calificaciones
- Banqueo de Hematologia Enaminternado Anual SabDocumento24 páginasBanqueo de Hematologia Enaminternado Anual SabJosue Dlc OrtizAún no hay calificaciones
- Inmunología 3Documento22 páginasInmunología 3Ignacio RotellaAún no hay calificaciones
- 1PARCIALDocumento16 páginas1PARCIALEmily CantorAún no hay calificaciones
- Hipersensibilidad Tipo II XDDocumento26 páginasHipersensibilidad Tipo II XDMendoza Rodríguez Danna MichelleAún no hay calificaciones
- 20-Actividad 3 - Octavo A DecimoDocumento5 páginas20-Actividad 3 - Octavo A DecimoLïz GómezAún no hay calificaciones
- Anatomia Toraco AbdominalDocumento17 páginasAnatomia Toraco AbdominalAbrahanAún no hay calificaciones
- Ciencias Naturales Guia #9 Sexto AlejandraDocumento5 páginasCiencias Naturales Guia #9 Sexto AlejandraNataliaAún no hay calificaciones