También podría gustarte
- Trilaminar ResumenDocumento8 páginasTrilaminar ResumenJonathan ClementeAún no hay calificaciones
- Disco Germinativo TrilaminarDocumento29 páginasDisco Germinativo TrilaminarJoseph Chilan Arteaga50% (2)
- 3ra Semana Del DesarrolloDocumento6 páginas3ra Semana Del DesarrolloAdri Mirelis Gaelyk ZoskolokAún no hay calificaciones
- TERCERA SEMANA DEL DESARROLLO. Embriología.Documento6 páginasTERCERA SEMANA DEL DESARROLLO. Embriología.Nakura IzaAún no hay calificaciones
- 3semanadedesarrollo 3 151127062225 Lva1 App6891Documento35 páginas3semanadedesarrollo 3 151127062225 Lva1 App6891Nestor Zury HVAún no hay calificaciones
- Meta 2.2.:VILLA LOPEZDocumento15 páginasMeta 2.2.:VILLA LOPEZFlor Denisse Villa LópezAún no hay calificaciones
- Meta2.2.:VILLA LÓPEZDocumento15 páginasMeta2.2.:VILLA LÓPEZFlor Denisse Villa LópezAún no hay calificaciones
- Capitulo 5 Embriologia 2023 .Documento25 páginasCapitulo 5 Embriologia 2023 .Mario RivasAún no hay calificaciones
- Capitulo 5 Tercera Semana de Desarrollo Disco Germinal Trilaminar 359286 Downloable 1777082Documento5 páginasCapitulo 5 Tercera Semana de Desarrollo Disco Germinal Trilaminar 359286 Downloable 1777082Yasmin TatianaAún no hay calificaciones
- Disco Germinativo TrilaminarDocumento7 páginasDisco Germinativo TrilaminarJoel Lopez MagneAún no hay calificaciones
- 3era Semana Del EmbarazoDocumento2 páginas3era Semana Del EmbarazoJeandely Ventura RodrguezAún no hay calificaciones
- Formación CorporalDocumento30 páginasFormación CorporalMiguel Angel Tristan RamirezAún no hay calificaciones
- M2.1 - Periodo EmbrionarioDocumento11 páginasM2.1 - Periodo EmbrionarioYamileth RamirezAún no hay calificaciones
- Tercera SemanaDocumento8 páginasTercera Semanalarapaloma698Aún no hay calificaciones
- Establecimiento de Los Ejes CorporalesDocumento2 páginasEstablecimiento de Los Ejes CorporalesPrinces Katherine Aguilar SantiagoAún no hay calificaciones
- Tercera Semana de Desarrollo: Disco Germinal TrilaminarDocumento6 páginasTercera Semana de Desarrollo: Disco Germinal TrilaminarBrenda Itzel Martinez AgustinAún no hay calificaciones
- Resumen Capitulo 5 LangmanDocumento7 páginasResumen Capitulo 5 LangmanDarkinfire Western100% (1)
- EMBRIOLOGIADocumento33 páginasEMBRIOLOGIAGABRIEL ALEJANDRO LOPEZ AVALOSAún no hay calificaciones
- 05 - Tercera Semana Del DesarrolloDocumento31 páginas05 - Tercera Semana Del DesarrolloHelena BarrientosAún no hay calificaciones
- Capítulo 7, de La Tercera A La Octava SemanaDocumento42 páginasCapítulo 7, de La Tercera A La Octava Semanajenny cruzAún no hay calificaciones
- Formación del Mesodermo y sus DerivadosDocumento4 páginasFormación del Mesodermo y sus DerivadosLidia RamirezAún no hay calificaciones
- Guà - A 3Documento11 páginasGuà - A 3Javier EspinozaAún no hay calificaciones
- Disco Germinativo TrilaminarDocumento7 páginasDisco Germinativo TrilaminarJosé Ramón Fernández ChacínAún no hay calificaciones
- Embriologia 13Documento7 páginasEmbriologia 13Duvan Zavaleta BazanAún no hay calificaciones
- Núcleo N6Documento22 páginasNúcleo N6Franco TaylorAún no hay calificaciones
- 2da y 3ra Semana de GestaciónDocumento4 páginas2da y 3ra Semana de GestaciónEsthefany AgostiniAún no hay calificaciones
- DE 3ERA A 8VA SEMANA DEL DESARROLLO - PERÍODO EMBRIONARIO (Embriologia Langmann 8ed)Documento6 páginasDE 3ERA A 8VA SEMANA DEL DESARROLLO - PERÍODO EMBRIONARIO (Embriologia Langmann 8ed)Maria Silva DiazAún no hay calificaciones
- GastrulaciónDocumento12 páginasGastrulaciónAna Lilia Santana LópezAún no hay calificaciones
- Resumnes de EmbriologiaDocumento18 páginasResumnes de EmbriologiaYari Hernandez100% (1)
- 5º La Gastrulación - Disco GerminativoDocumento7 páginas5º La Gastrulación - Disco GerminativoaleAún no hay calificaciones
- A2. 3ra A 8va Semana 2P - GBD 1010Documento6 páginasA2. 3ra A 8va Semana 2P - GBD 1010Guillermo Burgos DiazAún no hay calificaciones
- Informe Embriologia 1 Exposion Grupo 3Documento16 páginasInforme Embriologia 1 Exposion Grupo 3linsey infante foxAún no hay calificaciones
- Genes MorfoDocumento34 páginasGenes MorfoJose Luis Maizani LlanosAún no hay calificaciones
- Disco TrilaminarDocumento28 páginasDisco TrilaminarDayanna Armendariz SaltosAún no hay calificaciones
- Doctora: Shirley Sissi Choqueticlla Puma EstudiantesDocumento45 páginasDoctora: Shirley Sissi Choqueticlla Puma EstudiantesIvan Freddy Cruz PacoAún no hay calificaciones
- Ensayo de EmbriologiaDocumento9 páginasEnsayo de EmbriologiaJana Misseth Gaxiola CastroAún no hay calificaciones
- Cuestionario M2.3Documento6 páginasCuestionario M2.3Hector Efrain Garcia ChavezAún no hay calificaciones
- 4dela3a8semanadeldesarrollo-Documento77 páginas4dela3a8semanadeldesarrollo-Andrea SotoAún no hay calificaciones
- Embriologia Craneofacial DRA RODRIGUEZDocumento8 páginasEmbriologia Craneofacial DRA RODRIGUEZErika Rodriguez MarteAún no hay calificaciones
- Embriología: Desarrollo de la 3ra semanaDocumento18 páginasEmbriología: Desarrollo de la 3ra semanaFernanda Valeria Gutiérrez LeónAún no hay calificaciones
- Desarrollo EmbrionarioDocumento14 páginasDesarrollo EmbrionarioHesther CoronaAún no hay calificaciones
- EMBRIOLOGIA - Docx ExamenDocumento3 páginasEMBRIOLOGIA - Docx Examenjose luis burgaAún no hay calificaciones
- REGULACIÓN GENÉTICA EJES CUERPODocumento4 páginasREGULACIÓN GENÉTICA EJES CUERPOAndre ChevarriaAún no hay calificaciones
- Periodo Somitico 1 Vladimir FloresDocumento16 páginasPeriodo Somitico 1 Vladimir FloresChistian DavidAún no hay calificaciones
- Resumen Organismos ModelosDocumento6 páginasResumen Organismos ModelosPaula GiraldoAún no hay calificaciones
- Tercera Semana Del Desarrollo EmbriologicoDocumento6 páginasTercera Semana Del Desarrollo EmbriologicoAriana AntonioAún no hay calificaciones
- Resumen de Los 9 CapitulosDocumento36 páginasResumen de Los 9 CapitulosSebastian Pinto MirandaAún no hay calificaciones
- Desarrollo embrionario de la tercera a la octava semanaDocumento12 páginasDesarrollo embrionario de la tercera a la octava semanaEnoc GonzálezAún no hay calificaciones
- Definición de OrganizadorDocumento2 páginasDefinición de OrganizadortrenquelauquenAún no hay calificaciones
- Tema 2 Psicobiología II KoraDocumento4 páginasTema 2 Psicobiología II KorapalomaAún no hay calificaciones
- Resumen Embriologia Semanas 1 A 8Documento4 páginasResumen Embriologia Semanas 1 A 8Liza Cifuentes100% (1)
- Gastrulacion y NeurulacionDocumento6 páginasGastrulacion y NeurulacionVale MAún no hay calificaciones
- Embrio 2 Semana-1Documento50 páginasEmbrio 2 Semana-1xd cualquier vosa xd xd khe vrg :vAún no hay calificaciones
- Embrio 3Documento9 páginasEmbrio 3Meme Nella CastroAún no hay calificaciones
- 3 Carpeta de EmbriologíaDocumento129 páginas3 Carpeta de Embriologíaemelin garciaAún no hay calificaciones
- SistemaNerviosoDocumento64 páginasSistemaNerviosoFer FrAún no hay calificaciones
- Cuestionario M2.2Documento3 páginasCuestionario M2.2Hector Efrain Garcia ChavezAún no hay calificaciones
- Cap 5. GastrulaciónDocumento2 páginasCap 5. Gastrulación4-5 CHARRIS SAMUELAún no hay calificaciones
- Hito 4 - Informe Del Procesual - Caso Clinico 1Documento7 páginasHito 4 - Informe Del Procesual - Caso Clinico 1Daniela TrigoAún no hay calificaciones
- PROCESUAL 4 - KDTCDocumento15 páginasPROCESUAL 4 - KDTCDaniela TrigoAún no hay calificaciones
- Articulo Métodos Anticonceptivos - Daniela Trigo CaceresDocumento2 páginasArticulo Métodos Anticonceptivos - Daniela Trigo CaceresDaniela TrigoAún no hay calificaciones
- GametogénesisDocumento1 páginaGametogénesisDaniela TrigoAún no hay calificaciones
- Dilemas Éticos en El Contexto de La Pandemia Del Covid 19 - KDTCDocumento10 páginasDilemas Éticos en El Contexto de La Pandemia Del Covid 19 - KDTCDaniela TrigoAún no hay calificaciones
- Covid-19 - KDTCDocumento6 páginasCovid-19 - KDTCDaniela TrigoAún no hay calificaciones
- Kelen Daniela Trigo Caceres - Actividad 1 - Hito 4Documento12 páginasKelen Daniela Trigo Caceres - Actividad 1 - Hito 4Daniela TrigoAún no hay calificaciones
- Informe Nervio Vestibulococlear - KDTCDocumento6 páginasInforme Nervio Vestibulococlear - KDTCDaniela TrigoAún no hay calificaciones
- Kelne Daniela Trigo Caceres - ACTIVIDAD 2 - HITO 4Documento6 páginasKelne Daniela Trigo Caceres - ACTIVIDAD 2 - HITO 4Daniela TrigoAún no hay calificaciones
- Arquietctura EclécticaDocumento6 páginasArquietctura EclécticaDaniela TrigoAún no hay calificaciones
- Aplicacion de MaterialesDocumento3 páginasAplicacion de MaterialesDaniela TrigoAún no hay calificaciones
- Copia de Obtencion y Preparacion de Materias PrimasDocumento1 páginaCopia de Obtencion y Preparacion de Materias PrimasDaniela TrigoAún no hay calificaciones
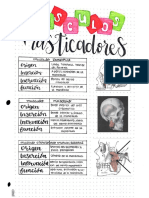
- Músculos Faciales y MasticadoresDocumento5 páginasMúsculos Faciales y MasticadoresDaniela TrigoAún no hay calificaciones
- Principios EticosDocumento10 páginasPrincipios EticosDaniela TrigoAún no hay calificaciones
- Historia del movimiento orgánico en la arquitecturaDocumento6 páginasHistoria del movimiento orgánico en la arquitecturaDaniela TrigoAún no hay calificaciones
- BRUTALISMODocumento4 páginasBRUTALISMODaniela TrigoAún no hay calificaciones
- Arquitectura Contemporanea LatinoamericanaDocumento8 páginasArquitectura Contemporanea LatinoamericanaDaniela TrigoAún no hay calificaciones
- Arquitectura EclécticaDocumento6 páginasArquitectura EclécticaDaniela TrigoAún no hay calificaciones
- MINIMALISMODocumento3 páginasMINIMALISMODaniela TrigoAún no hay calificaciones
- Arts & CraftsDocumento7 páginasArts & CraftsDaniela TrigoAún no hay calificaciones
- La Bauhause - El ConstructivismoDocumento5 páginasLa Bauhause - El ConstructivismoDaniela TrigoAún no hay calificaciones
- Curtido de Pieles IntroduccionDocumento94 páginasCurtido de Pieles IntroduccionMarco RomeroAún no hay calificaciones
- La logística a través de la historia: evolución del concepto y aplicacionesDocumento49 páginasLa logística a través de la historia: evolución del concepto y aplicacionesAlbertoAún no hay calificaciones
- Miastenia Gravis WalterDocumento8 páginasMiastenia Gravis WalterAnthony Sanchez MartinezAún no hay calificaciones
- Cuentos para AdultosDocumento1 páginaCuentos para Adultosgabriel peraltaAún no hay calificaciones
- NUTRICIÓN CANINA BÁSICA UNAM 2015 R Aguila PDFDocumento10 páginasNUTRICIÓN CANINA BÁSICA UNAM 2015 R Aguila PDFjhonAún no hay calificaciones
- Noelle I5501966Documento4 páginasNoelle I5501966Jhoompieer VFAún no hay calificaciones
- RecetasDocumento31 páginasRecetasOscar Gavilan RequenaAún no hay calificaciones
- Informe laboratorio análisis sangreDocumento12 páginasInforme laboratorio análisis sangreJuliana HortuaAún no hay calificaciones
- Rojo y negro: palabras clave del análisis literarioDocumento7 páginasRojo y negro: palabras clave del análisis literarioMonica Beatriz Urcia VegaAún no hay calificaciones
- INFORME DE INSPECCION JUDICIAL FINCA LA GUARONA Con FotosDocumento17 páginasINFORME DE INSPECCION JUDICIAL FINCA LA GUARONA Con Fotosangelica molinaAún no hay calificaciones
- Donde Vive El TigreDocumento7 páginasDonde Vive El TigreViviana Concha AnguitaAún no hay calificaciones
- Alma de Gato - Ruth BergerDocumento170 páginasAlma de Gato - Ruth BergerliclauramartinoAún no hay calificaciones
- OsificacionDocumento5 páginasOsificacionEli FernandoAún no hay calificaciones
- NEFROLOGIADocumento104 páginasNEFROLOGIAEdwinOmarRosarioLeon100% (1)
- JUEGOS POPULARESDocumento9 páginasJUEGOS POPULARESMaria AngélicaAún no hay calificaciones
- Lista de Logros Minecraft Java (Versión 1.20.4)Documento18 páginasLista de Logros Minecraft Java (Versión 1.20.4)Sebastián VidalAún no hay calificaciones
- Colás El Chico y Colás El Grande Hans Christian AndersenDocumento3 páginasColás El Chico y Colás El Grande Hans Christian AndersenIngridGonzalezAún no hay calificaciones
- La Biblia en Volúmenes T1Documento74 páginasLa Biblia en Volúmenes T1EuniceFustyAún no hay calificaciones
- Musculos Del CuelloDocumento3 páginasMusculos Del CuellosavkapericAún no hay calificaciones
- Triphala PDFDocumento5 páginasTriphala PDFparadojxAún no hay calificaciones
- Producción de gallinas procesadas en bandeja de 20 librasDocumento12 páginasProducción de gallinas procesadas en bandeja de 20 libraskarenAún no hay calificaciones
- Watson OkDocumento29 páginasWatson OkGrylam Odalys Aguedo ApolinarioAún no hay calificaciones
- Laboratorio Contabilidad GanaderaDocumento20 páginasLaboratorio Contabilidad GanaderaSANDRA PAULINA PABLO TEMAJAún no hay calificaciones
- El Corazon TrigonometricoDocumento1 páginaEl Corazon TrigonometricoJose RipAún no hay calificaciones
- Parkinson Rutina de Ejercicios de FrenkelDocumento6 páginasParkinson Rutina de Ejercicios de FrenkelAmabelia GuerrerosAún no hay calificaciones
- Síndrome NefróticoDocumento13 páginasSíndrome NefróticoxshighsxAún no hay calificaciones
- Definición de EcologíaDocumento2 páginasDefinición de Ecologíasamatarua100% (2)
- Proyecto Granja AutosostenibleDocumento12 páginasProyecto Granja AutosostenibleDani Leo Rada0% (1)
- Ogbe Fun.Documento19 páginasOgbe Fun.AwoIfabilaye Idin Oloyin100% (2)