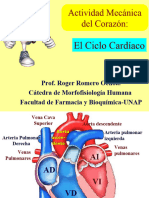
También podría gustarte
- Mecanica CardiacaDocumento8 páginasMecanica CardiacadaviidojedaAún no hay calificaciones
- Ciclo CardiacoDocumento6 páginasCiclo CardiacoexcadeteAún no hay calificaciones
- CardiovascularDocumento52 páginasCardiovascularFrancisco Altamirano CamposAún no hay calificaciones
- CorazonDocumento10 páginasCorazonJuan Camilo SagñayAún no hay calificaciones
- Ciclo CardiacoDocumento23 páginasCiclo Cardiacoramosmargarita996Aún no hay calificaciones
- RELATORIADocumento3 páginasRELATORIAJuan GomezAún no hay calificaciones
- Ciclo CardiacoDocumento6 páginasCiclo CardiacoSofía DalmolinAún no hay calificaciones
- Ciclo CardiacoDocumento11 páginasCiclo Cardiacoapi-3699557100% (2)
- Ciclo Cardiaco DR PeraltaDocumento40 páginasCiclo Cardiaco DR PeraltaEdgar Joaquin Lara VallejosAún no hay calificaciones
- Resumen Capitulo 9 FisioDocumento7 páginasResumen Capitulo 9 FisioJenny ValdezAún no hay calificaciones
- Ciclo CardiacoDocumento2 páginasCiclo CardiacoYessy nekoAún no hay calificaciones
- Capítulo 9 GuytonDocumento3 páginasCapítulo 9 GuytonFrancisco CubeAún no hay calificaciones
- El Corazón Como Bomba y Función de Las Válvulas Cardíacas: FisiologíaDocumento40 páginasEl Corazón Como Bomba y Función de Las Válvulas Cardíacas: FisiologíaGABRIEL TRIVEO ORTUOAún no hay calificaciones
- El CorazónDocumento11 páginasEl CorazónCaroline Menares CastilloAún no hay calificaciones
- Fisio Gasto CardiacoDocumento11 páginasFisio Gasto CardiacoRebeca Sandra Meza MayorgaAún no hay calificaciones
- Semana 1Documento5 páginasSemana 1Josseph SucapucaAún no hay calificaciones
- Ciclo CardiacoDocumento37 páginasCiclo CardiacoJoscelia SilvaAún no hay calificaciones
- Seminario de CorazonDocumento4 páginasSeminario de CorazonJomaes0411Aún no hay calificaciones
- Fisio Ciclo CardiacoDocumento6 páginasFisio Ciclo CardiacoAnabel RolónAún no hay calificaciones
- Ciclo CardiacoDocumento12 páginasCiclo CardiacoDiana Lizeth Cartagena JuarezAún no hay calificaciones
- Ciclo CardiacoDocumento23 páginasCiclo CardiacoenarmanykaAún no hay calificaciones
- Fisiologia Cardiovascular 2.0Documento18 páginasFisiologia Cardiovascular 2.0Denisse EsparzaAún no hay calificaciones
- Diagrama de WiggersDocumento8 páginasDiagrama de WiggersCatita Xd Tapia Sobarzo0% (2)
- El Corazón Como Bomba (Autoguardado)Documento27 páginasEl Corazón Como Bomba (Autoguardado)yoseannysgomez1617Aún no hay calificaciones
- Ciclo cardíaco-WPS OfficeDocumento4 páginasCiclo cardíaco-WPS OfficeMaríaAún no hay calificaciones
- Ciclo Cardiaco - 1Documento2 páginasCiclo Cardiaco - 1Daniel Londoño RoldanAún no hay calificaciones
- Fisiologia Del Ciclo Cardiaco y BombeoDocumento23 páginasFisiologia Del Ciclo Cardiaco y BombeoTamara Ruiz MaureiraAún no hay calificaciones
- Sistema ConductorDocumento4 páginasSistema ConductorBlanca Denisse Alonso rodriguezAún no hay calificaciones
- Parcial FisiologíaDocumento23 páginasParcial FisiologíaMaria Fernanda Carvajal CastillaAún no hay calificaciones
- Curva de Presion AorticaDocumento8 páginasCurva de Presion AorticajeandickAún no hay calificaciones
- Toma de Presión ArterialDocumento5 páginasToma de Presión ArterialDANNA LIZETTE CRUZ JUAREZAún no hay calificaciones
- Musculo Cardiaco El Corazon Como Bomba y La Funcion de Las Valvulas CardiacasDocumento3 páginasMusculo Cardiaco El Corazon Como Bomba y La Funcion de Las Valvulas CardiacasAlba Raquel Maldonado JiménezAún no hay calificaciones
- El CorazónDocumento6 páginasEl CorazónDAYRIN MILEYDI LINARES AGUIRREAún no hay calificaciones
- Fisiología CardiacaDocumento38 páginasFisiología CardiacaJose Oscategui C100% (1)
- El Ciclo CardiacoDocumento25 páginasEl Ciclo CardiacoRocio Lucas100% (1)
- Electrofisiologia Cardiaca 2014Documento22 páginasElectrofisiologia Cardiaca 2014Angella CamposAún no hay calificaciones
- Monografía Del Corazón (Fisiología Guyton)Documento8 páginasMonografía Del Corazón (Fisiología Guyton)ximenaraniAún no hay calificaciones
- Mecanica Cardiaca PDFDocumento59 páginasMecanica Cardiaca PDFmeli murAún no hay calificaciones
- Mecanica Cardiaca PDFDocumento59 páginasMecanica Cardiaca PDFmeli murAún no hay calificaciones
- 7.ciclo Cardiaco 1Documento40 páginas7.ciclo Cardiaco 1Oscar Paima CárdenasAún no hay calificaciones
- Apuntes Corazon FisioDocumento8 páginasApuntes Corazon FisioDylan VulgarinAún no hay calificaciones
- Auscultación Cardíaca I - Documentos de GoogleDocumento8 páginasAuscultación Cardíaca I - Documentos de GoogleCamilaAún no hay calificaciones
- Cuestionario Gasto CardiacoDocumento8 páginasCuestionario Gasto CardiacoRosangela Macavilca CruzAún no hay calificaciones
- Ciclo CardiacoDocumento5 páginasCiclo Cardiacoarturo reimiAún no hay calificaciones
- Fisio-25 2Documento4 páginasFisio-25 2Dani Fernandez GilarteAún no hay calificaciones
- Ciclo CardiacoDocumento14 páginasCiclo CardiacoAbigail AlvarezAún no hay calificaciones
- Tema 7. Ciclo Cardíaco. Gasto Cardíaco y Factores Que LoDocumento17 páginasTema 7. Ciclo Cardíaco. Gasto Cardíaco y Factores Que LoRosa SotoAún no hay calificaciones
- Ciclo Cardiaco WordDocumento3 páginasCiclo Cardiaco WordMedicina Sexto SemestreAún no hay calificaciones
- 13.4 Ciclo CardiacoDocumento4 páginas13.4 Ciclo CardiacoJoselyn Meriño SuarezAún no hay calificaciones
- Ciclo CardiacoDocumento4 páginasCiclo CardiacoPamela AguilarAún no hay calificaciones
- Ciclo CardiácoDocumento5 páginasCiclo CardiácoElizabeth Pérez LimaAún no hay calificaciones
- Función de Las Aurículas Como Bombas de CebadoDocumento31 páginasFunción de Las Aurículas Como Bombas de CebadoSebastián Tello Armijos50% (2)
- Ciclo Cardíaco-Equipo 3 - Gpo261Documento19 páginasCiclo Cardíaco-Equipo 3 - Gpo261Melanye MartinezAún no hay calificaciones
- Ciclo CardíacoDocumento10 páginasCiclo CardíacoyubissaAún no hay calificaciones
- Cuestionario Sobre El Capítulo 9 y 23 de Fisiología de GuytonDocumento9 páginasCuestionario Sobre El Capítulo 9 y 23 de Fisiología de GuytonKatherine YisselAún no hay calificaciones
- Resumen de El Corazón Como Bomba - FISIOLOGÍA DE GUYTONDocumento4 páginasResumen de El Corazón Como Bomba - FISIOLOGÍA DE GUYTONJaime Jara AlvaradoAún no hay calificaciones
- Mejorar la Circulación Sanguínea de Forma Natural: Sanar el Sistema Circulatorio y inmunológico para Prevenir Enfermedades, Aumentar la Energía, Aliviar el Estrés y Sentirte Mejor que NuncaDe EverandMejorar la Circulación Sanguínea de Forma Natural: Sanar el Sistema Circulatorio y inmunológico para Prevenir Enfermedades, Aumentar la Energía, Aliviar el Estrés y Sentirte Mejor que NuncaCalificación: 4.5 de 5 estrellas4.5/5 (7)
- Presión Arterial Baja: Cómo reconocer, manejar y solucionar esta alteración que también puede afectar tu bienestarDe EverandPresión Arterial Baja: Cómo reconocer, manejar y solucionar esta alteración que también puede afectar tu bienestarCalificación: 5 de 5 estrellas5/5 (1)
- Introducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludDe EverandIntroducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludAún no hay calificaciones
- 2-Membrana CelularDocumento10 páginas2-Membrana CelularJoyas Cosmeticos SolangeAún no hay calificaciones
- Sistema RenalDocumento8 páginasSistema RenalJoyas Cosmeticos SolangeAún no hay calificaciones
- 1-Introducción FisiologíaDocumento24 páginas1-Introducción FisiologíaJoyas Cosmeticos SolangeAún no hay calificaciones
- Sistema RespiratorioDocumento16 páginasSistema RespiratorioJoyas Cosmeticos SolangeAún no hay calificaciones
- Original Trabajo Mitos y Leyendas Zona CentralDocumento14 páginasOriginal Trabajo Mitos y Leyendas Zona CentralJoyas Cosmeticos SolangeAún no hay calificaciones
- Accede Al Ilimitado Poder de Tu Mente y Transforma Tu Vida 12 SetiembreDocumento109 páginasAccede Al Ilimitado Poder de Tu Mente y Transforma Tu Vida 12 Setiembreomkara7777777Aún no hay calificaciones
- Anatomía Nervioso VeterinariaDocumento5 páginasAnatomía Nervioso VeterinariaTANIA DENISE CORTES ISLASAún no hay calificaciones
- Lacrimacion CPODocumento6 páginasLacrimacion CPOJossie TeneriaAún no hay calificaciones
- Material de Lectura Clase 6Documento53 páginasMaterial de Lectura Clase 6Ale BentAún no hay calificaciones
- Repaso de La Unidad de Sistema Nervioso. Autoevaluación EDAIN MONSERRAT BARBA SALINASDocumento9 páginasRepaso de La Unidad de Sistema Nervioso. Autoevaluación EDAIN MONSERRAT BARBA SALINASmonse barbaAún no hay calificaciones
- Consideraciones NeuroanatomicasDocumento4 páginasConsideraciones NeuroanatomicasYenifer Huamani AncasiAún no hay calificaciones
- Bynf Vi-MovimientoDocumento23 páginasBynf Vi-MovimientoPaula PajónAún no hay calificaciones
- El Sistema Nervioso Autónomo para Tercero de SecundariaDocumento6 páginasEl Sistema Nervioso Autónomo para Tercero de SecundariaMara LoplanAún no hay calificaciones
- Pulmones DerechoDocumento36 páginasPulmones Derechoalex adrianoAún no hay calificaciones
- Sistema Nervioso PeriféricoDocumento2 páginasSistema Nervioso PeriféricoKEVIN TEBES BENAVIDES GARCIAAún no hay calificaciones
- Farmacología Del SNCDocumento7 páginasFarmacología Del SNCAdriana MarinAún no hay calificaciones
- 9 Guía de Estudio 1-9 CorazónDocumento12 páginas9 Guía de Estudio 1-9 CorazónagustinaAún no hay calificaciones
- Guia de AutoestudioDocumento7 páginasGuia de AutoestudioMinastauriel AlassëaAún no hay calificaciones
- Afi2 U1 A1 NotcDocumento19 páginasAfi2 U1 A1 NotcNorma Patricia Torres CabreraAún no hay calificaciones
- Sistema Nervioso AutonomoDocumento5 páginasSistema Nervioso AutonomoHernández Arroyo RodrigoAún no hay calificaciones
- GUIA DE ANATOMÍA SESION 2 Sistema Nervioso II, Ojos, OídosDocumento8 páginasGUIA DE ANATOMÍA SESION 2 Sistema Nervioso II, Ojos, OídosMayka YAHUANAAún no hay calificaciones
- Fisiologia Del Sistema Reproductor GanonDocumento42 páginasFisiologia Del Sistema Reproductor GanonAngeline GuerreroAún no hay calificaciones
- CLASE 1 y 2Documento19 páginasCLASE 1 y 2lilianaAún no hay calificaciones
- SimpaticomiméticosDocumento15 páginasSimpaticomiméticosDebra Rosas100% (1)
- Detwcción de Mentira (Poligrafía)Documento35 páginasDetwcción de Mentira (Poligrafía)ScribdTranslationsAún no hay calificaciones
- Mencione Las Características Básicas de La Función Simpática y ParasimpáticaDocumento2 páginasMencione Las Características Básicas de La Función Simpática y ParasimpáticaJostin Moises Santos AlmonteAún no hay calificaciones
- Concluciones 22Documento3 páginasConcluciones 22Leidi Flor Uriarte SaldañaAún no hay calificaciones
- NTP 513 Plaguicidas Organofosforados 2 Toxicodinamia y Control Biológico 2Documento6 páginasNTP 513 Plaguicidas Organofosforados 2 Toxicodinamia y Control Biológico 2Jesica Andrea Nina EugenioAún no hay calificaciones
- Notas de Anatomía para EstudiantesDocumento268 páginasNotas de Anatomía para EstudiantesVelmarAún no hay calificaciones
- Fisiología Del Sistema Nervioso AutónomoDocumento11 páginasFisiología Del Sistema Nervioso AutónomoNathaly MontésAún no hay calificaciones
- Fisiologia Del Sistema NerviosoDocumento22 páginasFisiologia Del Sistema NerviosoHaiber TintinagoAún no hay calificaciones
- Comienza Tu Viaje Por La Nueva Medicina Germanica PDFDocumento94 páginasComienza Tu Viaje Por La Nueva Medicina Germanica PDFSERGIO alarcon88% (8)
- Enfermedades Del Sistema Nervioso.Documento30 páginasEnfermedades Del Sistema Nervioso.Andrea PaosulineAún no hay calificaciones
- Notas Rápidas PDFDocumento2 páginasNotas Rápidas PDFMónica Andrea Quispe RojasAún no hay calificaciones
- PDF Los Cuatro Pilares de La SaludpdfDocumento209 páginasPDF Los Cuatro Pilares de La SaludpdfJoseph Antonio Apaza GómezAún no hay calificaciones