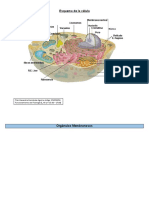
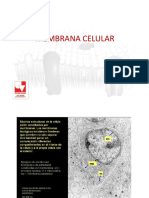
También podría gustarte
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
- Materia Solemne II Biología Pte 1Documento7 páginasMateria Solemne II Biología Pte 1karlarenatavalenzuelasAún no hay calificaciones
- Trabajo N1 La Neurona UCESDocumento7 páginasTrabajo N1 La Neurona UCESFlorcita Dominguez BankowskiAún no hay calificaciones
- Membranas Celulares InternasDocumento9 páginasMembranas Celulares InternasANA SIBAJAAún no hay calificaciones
- Organelos IIDocumento8 páginasOrganelos IIantonia muñozAún no hay calificaciones
- Cuadro Resumen Organelos CitoplásmaticosDocumento2 páginasCuadro Resumen Organelos Citoplásmaticosmilagros bruzual50% (2)
- Trafico de ProteinasDocumento22 páginasTrafico de ProteinasMilkobich 05Aún no hay calificaciones
- (ppt6) Compartimientos y Transporte Intracelular 1 y 2 PDFDocumento8 páginas(ppt6) Compartimientos y Transporte Intracelular 1 y 2 PDFcamila palomino100% (2)
- Celula Eucariotica y Sus FuncionesDocumento3 páginasCelula Eucariotica y Sus FuncionesJulieth Lorena Ruiz100% (4)
- Celula VegetalDocumento6 páginasCelula VegetalPerla Midary Dominguez CruzAún no hay calificaciones
- Unidad 2 Sesión 3 - Ribosomas - RER y REL FSDocumento27 páginasUnidad 2 Sesión 3 - Ribosomas - RER y REL FSTamara Riquelme PerezAún no hay calificaciones
- Histologia N3-1Documento5 páginasHistologia N3-1Samantha Cardenas VilcaAún no hay calificaciones
- CLASE 3 Sistema de EndomembranasDocumento30 páginasCLASE 3 Sistema de EndomembranasKeren RuizAún no hay calificaciones
- Documento Sin TítuloDocumento7 páginasDocumento Sin TítuloabayodeAún no hay calificaciones
- Clase 9 Destinacion de Proteinas Via Exocitica. Fono 2023 Editado 1Documento30 páginasClase 9 Destinacion de Proteinas Via Exocitica. Fono 2023 Editado 1Darlen AlvarezAún no hay calificaciones
- CLASE 6 Reticulo Endoplasmatico MEDICINA 2021 FINALDocumento23 páginasCLASE 6 Reticulo Endoplasmatico MEDICINA 2021 FINALya tu sabeAún no hay calificaciones
- Sistema de EndomembranasDocumento13 páginasSistema de EndomembranasDiegoArenasCataldoAún no hay calificaciones
- Histologia N3Documento5 páginasHistologia N3Samantha Cardenas VilcaAún no hay calificaciones
- Organelos MembranososDocumento17 páginasOrganelos MembranososAline Mac Pherson VásquezAún no hay calificaciones
- La CélulaDocumento7 páginasLa CélulaAlexandra VizcayaAún no hay calificaciones
- RamirezSilva Base57 Tarea02Documento2 páginasRamirezSilva Base57 Tarea02jesus.ramirez5534Aún no hay calificaciones
- Orgánulos CelularesDocumento21 páginasOrgánulos Celulares30211202046Aún no hay calificaciones
- Histologia N3 2Documento8 páginasHistologia N3 2Samantha Cardenas VilcaAún no hay calificaciones
- Transporte Intracelular de Proteinas FUSMDocumento52 páginasTransporte Intracelular de Proteinas FUSMBrayan David Martinez MartinezAún no hay calificaciones
- Organelos Célula EucariotaDocumento4 páginasOrganelos Célula Eucariotavalentina valarezoAún no hay calificaciones
- OrganelosDocumento2 páginasOrganelosRoberto AyalaAún no hay calificaciones
- Catedra de Biología 3 de MayoDocumento9 páginasCatedra de Biología 3 de MayoCamila Olguin GutierrezAún no hay calificaciones
- Modulo III Reticulo Endoplásmicosistema de Endomembranas Con Audio Final 2023Documento29 páginasModulo III Reticulo Endoplásmicosistema de Endomembranas Con Audio Final 2023Madelin BolañosAún no hay calificaciones
- Rejilla Capitulo 8Documento6 páginasRejilla Capitulo 8Wendy Paola Barragan JimenezAún no hay calificaciones
- TO Organelos IDocumento22 páginasTO Organelos IConstanza ValentinAún no hay calificaciones
- Cuadro de Los Organelos en Las Células EucariotasDocumento3 páginasCuadro de Los Organelos en Las Células EucariotasJackelim ReyesAún no hay calificaciones
- Estructura CelularDocumento4 páginasEstructura CelularJorge Aranda MedinaAún no hay calificaciones
- Histologia N3Documento5 páginasHistologia N3Samantha Cardenas VilcaAún no hay calificaciones
- Cuadro de OrganelosDocumento2 páginasCuadro de OrganelosDafAún no hay calificaciones
- Los Organulos CelularesDocumento30 páginasLos Organulos CelularesRainbow JocbkAún no hay calificaciones
- 9° Apunte de Cátedra Biologia Humana Retículo Endoplasmático Y Complejo de Golgi Tráfico VesicularDocumento11 páginas9° Apunte de Cátedra Biologia Humana Retículo Endoplasmático Y Complejo de Golgi Tráfico VesiculardanitzaAún no hay calificaciones
- Tema 5 Endomembranas Acortado 2018-2019Documento46 páginasTema 5 Endomembranas Acortado 2018-2019Daniel Fernandez-ValenciaAún no hay calificaciones
- Organelos Citoplasmaticos 2022Documento4 páginasOrganelos Citoplasmaticos 2022OlivSanMaria100% (1)
- La CelulaDocumento23 páginasLa CelulaNicolásAún no hay calificaciones
- Sem05 BiologíaDocumento34 páginasSem05 Biologíajharen morillo palominoAún no hay calificaciones
- Biología CelularDocumento8 páginasBiología CelularTriana CanalesAún no hay calificaciones
- Biología Bloque 2Documento60 páginasBiología Bloque 2Kelly Valencia LoorAún no hay calificaciones
- Esquema de La CélulaDocumento10 páginasEsquema de La CélulaPILAR ALEXANDRA HERNANDEZ AGUIRREAún no hay calificaciones
- Bic U2 t3 PDF 18Documento5 páginasBic U2 t3 PDF 18bombitopazAún no hay calificaciones
- Resumen Histo Todos Los TemasDocumento65 páginasResumen Histo Todos Los TemasM PAún no hay calificaciones
- Distribución de Macro MolesculasDocumento40 páginasDistribución de Macro MolesculasPaloma VeronAún no hay calificaciones
- 6 BiomembraDocumento8 páginas6 BiomembraNataly FerreiraAún no hay calificaciones
- Clase 12 Sistemas de EndomembranasDocumento22 páginasClase 12 Sistemas de EndomembranasJuan Pablo LempAún no hay calificaciones
- Flashcard O.Membranosos.Documento1 páginaFlashcard O.Membranosos.Isabella Muñoz CamposAún no hay calificaciones
- Distribución y Transporte de ProteínasDocumento41 páginasDistribución y Transporte de ProteínasJhonny Ponce Agui100% (2)
- U2 - Sesión 8 - EndomembranasDocumento42 páginasU2 - Sesión 8 - EndomembranasJosé Pablo QuevedoAún no hay calificaciones
- T6 EndomembranasDocumento18 páginasT6 Endomembranasalberto rodriguez lopezAún no hay calificaciones
- Seminario BC3-2020Documento70 páginasSeminario BC3-2020Oscar MartinezAún no hay calificaciones
- Organelos CelularesDocumento20 páginasOrganelos CelularesdevyAún no hay calificaciones
- Prueba n2 de BiologiaDocumento22 páginasPrueba n2 de BiologiaNohelis MelgarAún no hay calificaciones
- Sistema de Endomembranas IDocumento9 páginasSistema de Endomembranas IRocio RollanAún no hay calificaciones
- Biologia 2024-IiDocumento12 páginasBiologia 2024-Iiisabel chaconAún no hay calificaciones
- Estructura de La CelulaDocumento6 páginasEstructura de La Celulacarlos ticsAún no hay calificaciones
- Célula Eucariota AnimalDocumento4 páginasCélula Eucariota AnimalPerla PagesAún no hay calificaciones
- Sistema de Membranas Citoplasmaticas o Sistema de EndomembranasDocumento9 páginasSistema de Membranas Citoplasmaticas o Sistema de EndomembranasValentina CelaniAún no hay calificaciones
- Canales IónicosDocumento10 páginasCanales IónicosMariajosé Spataro MatheusAún no hay calificaciones
- CLASE 3 Control PostraduccionalDocumento47 páginasCLASE 3 Control PostraduccionalmelissaAún no hay calificaciones
- Expo SISTEMA MEMBRANASDocumento28 páginasExpo SISTEMA MEMBRANASVANIA ALEXA COTRINA ESTEVESAún no hay calificaciones
- Proteinas Biologia LauDocumento14 páginasProteinas Biologia LauRita TolabaAún no hay calificaciones
- Atlas Final 2020 Grupo No.12Documento48 páginasAtlas Final 2020 Grupo No.12graciela del rosario GalindoAún no hay calificaciones
- Translocon WikiDocumento2 páginasTranslocon WikiDiego J-fAún no hay calificaciones
- Reticulo EndoplasmaticoDocumento10 páginasReticulo EndoplasmaticoAnnaAún no hay calificaciones
- Sistemas de Secreción BDocumento49 páginasSistemas de Secreción Batsushi sakuraiAún no hay calificaciones
- 11-Sistema de Endomembranas-Apunte GeneralDocumento19 páginas11-Sistema de Endomembranas-Apunte GeneralTomas Scialfa BenderskyAún no hay calificaciones
- Guia Translocacion ProteicaDocumento4 páginasGuia Translocacion Proteicapatricia rodriguezAún no hay calificaciones
- Resumen Biocel Solemne 2Documento25 páginasResumen Biocel Solemne 2Josefa CastilloAún no hay calificaciones
- Sistemas de SecrecionDocumento29 páginasSistemas de SecrecionPaulina Santiago de RosasAún no hay calificaciones
- Sistema de EndomembranasDocumento50 páginasSistema de EndomembranasAngela AliagaAún no hay calificaciones
- Clase 4 Microbiología SanitariaDocumento31 páginasClase 4 Microbiología SanitariaCopy TextAún no hay calificaciones
- Reticulo Endoplasmatico Aparato de Golgi LisosomasDocumento28 páginasReticulo Endoplasmatico Aparato de Golgi LisosomasDiegoAún no hay calificaciones
- 5.wuolah Premium Apuntes T5 Sistemas de Membranas CitoplasmaticasDocumento27 páginas5.wuolah Premium Apuntes T5 Sistemas de Membranas CitoplasmaticasMARTA 036Aún no hay calificaciones
- Preguntas Cap 11Documento12 páginasPreguntas Cap 11Ariadni ChiquilloAún no hay calificaciones
- Repaso Módulo 2Documento30 páginasRepaso Módulo 2Laura Camila Alvarez HernandezAún no hay calificaciones
- Sistemas de SecrecionDocumento51 páginasSistemas de SecrecionMari Marisol RodriguezAún no hay calificaciones
- U2c SistemasSecrecion 19651Documento48 páginasU2c SistemasSecrecion 19651Unbreakable CreativityAún no hay calificaciones
- Reticulo EndolasmaticoDocumento16 páginasReticulo EndolasmaticoDavid PeralesAún no hay calificaciones
- Unidad 3 - G1 Sistema de Endomembranas 2020Documento89 páginasUnidad 3 - G1 Sistema de Endomembranas 2020Daiana DinamarcaAún no hay calificaciones
- Slide 22Documento42 páginasSlide 22hfhviufjiirkowowkwiqowoidfAún no hay calificaciones
- Trafico ExociticoDocumento13 páginasTrafico ExociticoIsi AnimatedAún no hay calificaciones
- Resumen Capitulo 16.1 de LodishDocumento1 páginaResumen Capitulo 16.1 de LodishDanii Alejandra100% (1)
- TEMA 10 Apuntes Bio-CelDocumento12 páginasTEMA 10 Apuntes Bio-CelBelén FernándezAún no hay calificaciones
- Insulin SyntesisDocumento23 páginasInsulin SyntesisJefren DaliosAún no hay calificaciones
- Trafico de ProteinasDocumento22 páginasTrafico de ProteinasMilkobich 05Aún no hay calificaciones
- Membrana Celular y Sistema Endomembranoso PDFDocumento65 páginasMembrana Celular y Sistema Endomembranoso PDFDanielAún no hay calificaciones