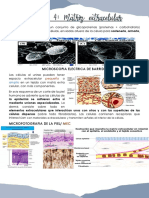
También podría gustarte
- Ados 2Documento3 páginasAdos 2Victoria Zorzopulos50% (2)
- Hormonas de Importancia para El Equilibrio HomeostaticoDocumento16 páginasHormonas de Importancia para El Equilibrio HomeostaticoTorresIvan20% (5)
- Guia Taller 4 Adhesion - CelularDocumento12 páginasGuia Taller 4 Adhesion - CelularKAREN MONSERRAT FRANCO ACHARAún no hay calificaciones
- S5-Matriz Celular ExtracelularDocumento18 páginasS5-Matriz Celular ExtracelularJhony Farith Farfán ReáteguiAún no hay calificaciones
- Tema 3Documento7 páginasTema 3letiAún no hay calificaciones
- Actividad 11 TallerSeñalizacionCelular-InteracciónCélulaCélulaDocumento9 páginasActividad 11 TallerSeñalizacionCelular-InteracciónCélulaCélulaAndrés FelipeAún no hay calificaciones
- INFORME S8 - Biología SeminariosDocumento8 páginasINFORME S8 - Biología SeminariosEsteban Gabriel Velásquez AdrianzénAún no hay calificaciones
- Uniones ComplejasDocumento6 páginasUniones ComplejasCarolina Alexandra ZEBALLOS LIPAAún no hay calificaciones
- Moleculas de Adhesion Celular 1223605666724403 9Documento54 páginasMoleculas de Adhesion Celular 1223605666724403 9Karen CabanaAún no hay calificaciones
- Adhesión y MECDocumento25 páginasAdhesión y MECJesus Gallardo CernaAún no hay calificaciones
- Adhesion Celular y Matriz ExtracelularDocumento14 páginasAdhesion Celular y Matriz ExtracelularIvan SolisAún no hay calificaciones
- X El MicroambienteDocumento6 páginasX El MicroambienteDiana Karen Butanda PuenteAún no hay calificaciones
- Catabolismo y AnabolismoDocumento4 páginasCatabolismo y AnabolismoKevin PachecoAún no hay calificaciones
- Membrana CelularDocumento5 páginasMembrana CelularKeniia AyalaAún no hay calificaciones
- Tema 7 Uniones CelularesDocumento6 páginasTema 7 Uniones CelularesDavid Molina LópezAún no hay calificaciones
- Clase 2 - Funcionamiento Celular (25 de Agosto)Documento10 páginasClase 2 - Funcionamiento Celular (25 de Agosto)Polette PalominosAún no hay calificaciones
- BC Clase 12Documento10 páginasBC Clase 12Diego AvendañoAún no hay calificaciones
- Clase 6 Biologia Comun - Membrana Plasmatica y TransportesDocumento22 páginasClase 6 Biologia Comun - Membrana Plasmatica y TransportesAlessyaAún no hay calificaciones
- Interacción Entre Las Células y Su EntornoDocumento8 páginasInteracción Entre Las Células y Su EntornoAnita EspinosaAún no hay calificaciones
- Diferenciaciones y MatrizDocumento17 páginasDiferenciaciones y MatrizmarinainesgonAún no hay calificaciones
- Taller Ensamblajes SupramolecularesDocumento9 páginasTaller Ensamblajes SupramolecularesLAURA MENA LEALAún no hay calificaciones
- FICHA 2 - Citología 1 IM 2023Documento43 páginasFICHA 2 - Citología 1 IM 2023Yamila Nicel LescanoAún no hay calificaciones
- BC 10 NecroticaenfmedDocumento4 páginasBC 10 NecroticaenfmedCasa LimpiezaAún no hay calificaciones
- Tema 03Documento7 páginasTema 03Rudolf CickoAún no hay calificaciones
- Histologia Humana - Sem 1Documento7 páginasHistologia Humana - Sem 1Clara Ramos CerranoAún no hay calificaciones
- M1 - 2 - Biologia (Comu - Trans)Documento43 páginasM1 - 2 - Biologia (Comu - Trans)Alejandra Torregrosa GutierrezAún no hay calificaciones
- Matriz IntracelularDocumento4 páginasMatriz Intracelularmirna victoria reynoso lopezAún no hay calificaciones
- Biología - Química UNED, Tema5Documento3 páginasBiología - Química UNED, Tema5Marta Arias FernandezAún no hay calificaciones
- Adhesion CelularDocumento7 páginasAdhesion CelularKate de LeonAún no hay calificaciones
- Clase 5. CitoesqueletoDocumento41 páginasClase 5. CitoesqueletofranciscoAún no hay calificaciones
- Los Filamentos de Actina en La Zona Apical Del Intestino DelgadoDocumento5 páginasLos Filamentos de Actina en La Zona Apical Del Intestino DelgadoYisenia VillegasAún no hay calificaciones
- Integración Célula Organismo - Moléculas de AdhesiónDocumento41 páginasIntegración Célula Organismo - Moléculas de Adhesiónrabl9250% (2)
- PDF Las Interacciones de Las Celulas Entre Si Grupo 5 DDDocumento7 páginasPDF Las Interacciones de Las Celulas Entre Si Grupo 5 DDpilar morochoAún no hay calificaciones
- TEMA 3 Cubiertas ExternasDocumento8 páginasTEMA 3 Cubiertas ExternasALEX BERMUDOAún no hay calificaciones
- Moleculas de Adhesion CelularDocumento3 páginasMoleculas de Adhesion Celularjuan dominguez100% (1)
- Membrana CelularDocumento16 páginasMembrana Celularni502556Aún no hay calificaciones
- Membrana Celular, Transporte y ParedDocumento15 páginasMembrana Celular, Transporte y ParedFernando LeonAún no hay calificaciones
- E6 Especializaciones de MembranaDocumento29 páginasE6 Especializaciones de MembranaIago AraújoAún no hay calificaciones
- Biologia Celular - Uab - Nanociencia y Nanotecnología Tema 15Documento9 páginasBiologia Celular - Uab - Nanociencia y Nanotecnología Tema 15SergioHerceHueteAún no hay calificaciones
- Moléculas de Relación CelularDocumento15 páginasMoléculas de Relación CelularAlejandro SuaresAún no hay calificaciones
- Fundamentos de La Célula y El CitoplasmaDocumento5 páginasFundamentos de La Célula y El CitoplasmaLuis InfanteAún no hay calificaciones
- Cuáles Son Las Proteínas Que Actúan Como Elementos Estructurales de La MembranaDocumento12 páginasCuáles Son Las Proteínas Que Actúan Como Elementos Estructurales de La MembranaGilmer Bautista TerronesAún no hay calificaciones
- Conferencia Citoplasma Completo OkDocumento60 páginasConferencia Citoplasma Completo OkPawan Alvarez PerezAún no hay calificaciones
- HOja de Trabajo Semana 13 - 1Documento7 páginasHOja de Trabajo Semana 13 - 1Estrella PalaciosAún no hay calificaciones
- Clase 2 - Funcionamiento Celular (25 de Agosto)Documento10 páginasClase 2 - Funcionamiento Celular (25 de Agosto)Polette PalominosAún no hay calificaciones
- T.5 Uniones y Adhesiones CelularesDocumento9 páginasT.5 Uniones y Adhesiones CelularesNatalia Villar CrespoAún no hay calificaciones
- Lucía Zárate Ramírez - Actividad 2 (Membrana Celular) - 0Documento7 páginasLucía Zárate Ramírez - Actividad 2 (Membrana Celular) - 0Lucy ZárateAún no hay calificaciones
- Biología GlosarioDocumento10 páginasBiología GlosarioMichelle FernandaAún no hay calificaciones
- Matriz Extracelular. Glicoproteínas. Atlas de Histología Vegetal y AnimalDocumento8 páginasMatriz Extracelular. Glicoproteínas. Atlas de Histología Vegetal y AnimalSolange MartínezAún no hay calificaciones
- Bioquimica de Las Membranas CellaresDocumento31 páginasBioquimica de Las Membranas CellaresJunior AlencarAún no hay calificaciones
- Membrana CelularDocumento39 páginasMembrana Celularkevin palloAún no hay calificaciones
- Semana 09 Citoesqueleto y Uniones CelularesDocumento27 páginasSemana 09 Citoesqueleto y Uniones CelularesDiana camila TiradoAún no hay calificaciones
- Especializaciones de Membrana CelularDocumento43 páginasEspecializaciones de Membrana Celularalejandra battiAún no hay calificaciones
- Matriz Extracelular de BiologíaDocumento3 páginasMatriz Extracelular de BiologíamariaoaAún no hay calificaciones
- Uniones Celulares y Matriz Extracelular: Biología Celular BCUM 1000 Prof. Alejandro ArriagadaDocumento32 páginasUniones Celulares y Matriz Extracelular: Biología Celular BCUM 1000 Prof. Alejandro Arriagadaandrea nettle bareaAún no hay calificaciones
- Interacciones Entre Células y Su AmbienteDocumento13 páginasInteracciones Entre Células y Su Ambientehemlocks100% (1)
- Clase 5 Matriz Extracelular y Adhesión Celular BetaDocumento41 páginasClase 5 Matriz Extracelular y Adhesión Celular BetaIsai QuirozAún no hay calificaciones
- Clase 4 Biologa Comn - Membrana Plasmtica y Transporte CompressedDocumento22 páginasClase 4 Biologa Comn - Membrana Plasmtica y Transporte CompressedMarx LopezAún no hay calificaciones
- Biología Semana9 - Sesión 17 - 2021-2Documento27 páginasBiología Semana9 - Sesión 17 - 2021-2Paula ManriqueAún no hay calificaciones
- Biología Semana9 Sesión 17 2021-2Documento27 páginasBiología Semana9 Sesión 17 2021-2Jerry VilcaAún no hay calificaciones
- Análisis funcional y microbiológico de derivados lácteos y cárnicosDe EverandAnálisis funcional y microbiológico de derivados lácteos y cárnicosAún no hay calificaciones
- Biología Molecular y Celular: Una guía introductoria para aprender Biología Celular y MolecularDe EverandBiología Molecular y Celular: Una guía introductoria para aprender Biología Celular y MolecularAún no hay calificaciones
- Tema 2 BMEPDocumento15 páginasTema 2 BMEPPaola Moyano GómezAún no hay calificaciones
- Tema 4 ImprimirDocumento13 páginasTema 4 ImprimirPaola Moyano GómezAún no hay calificaciones
- Tema 1 BMEPDocumento6 páginasTema 1 BMEPPaola Moyano GómezAún no hay calificaciones
- Tema 5 ImprimirDocumento14 páginasTema 5 ImprimirPaola Moyano GómezAún no hay calificaciones
- Tema 3 ImprimirDocumento11 páginasTema 3 ImprimirPaola Moyano GómezAún no hay calificaciones
- Tema 9 IMPRIMIRDocumento15 páginasTema 9 IMPRIMIRPaola Moyano GómezAún no hay calificaciones
- Tema 5 IMPRIMIRDocumento11 páginasTema 5 IMPRIMIRPaola Moyano GómezAún no hay calificaciones
- Tema 8 IMPRIMIRDocumento6 páginasTema 8 IMPRIMIRPaola Moyano GómezAún no hay calificaciones
- Tema 6 CIC IMPRIMIRDocumento7 páginasTema 6 CIC IMPRIMIRPaola Moyano GómezAún no hay calificaciones
- Hemoptisis Abordaje y TratamientoDocumento29 páginasHemoptisis Abordaje y TratamientoMariajosé BojorquezAún no hay calificaciones
- Paralisis Facial IdiopaticaDocumento13 páginasParalisis Facial IdiopaticaJose BernalAún no hay calificaciones
- Cap. 22 GeneDocumento5 páginasCap. 22 GeneAlexia AlvaradoAún no hay calificaciones
- Dx. Tratamiento Del Sindrome Sensorial en Adulto MayorDocumento9 páginasDx. Tratamiento Del Sindrome Sensorial en Adulto MayorJose Alejandro Palacios LópezAún no hay calificaciones
- Criterios de Clasificación AnimalDocumento3 páginasCriterios de Clasificación AnimalScott Mcgee80% (5)
- Alteraciones en El Ciclo Celular de Las Neuronas Podrían Relacionarse Con La Degeneración en AlzhéimerDocumento4 páginasAlteraciones en El Ciclo Celular de Las Neuronas Podrían Relacionarse Con La Degeneración en AlzhéimerPerla Li FigueroaAún no hay calificaciones
- Mastitis GangrenosaDocumento3 páginasMastitis Gangrenosasolange floresAún no hay calificaciones
- 1025 5583 Afm 82 01 00021Documento6 páginas1025 5583 Afm 82 01 00021dianabrito72273928Aún no hay calificaciones
- Taller Prevencion Anorexia y Bulimia Liliana ArceDocumento14 páginasTaller Prevencion Anorexia y Bulimia Liliana ArceSilvaniz Becerra Quiroz100% (1)
- Parametros Microbiologia de AlimentosDocumento12 páginasParametros Microbiologia de AlimentosJahir ArizaAún no hay calificaciones
- Organelos y CelulasDocumento30 páginasOrganelos y CelulasRomilioAntonioRetamalesGermanottaEncinaAún no hay calificaciones
- JUSTIFICACIONDocumento4 páginasJUSTIFICACIONAzalia Lopez RodriguezAún no hay calificaciones
- Ciclo BiologicoDocumento2 páginasCiclo BiologicomatiasAún no hay calificaciones
- Lupus Eritematoso Sitemico (1) CasoDocumento34 páginasLupus Eritematoso Sitemico (1) CasoRomulo Alegre LeónAún no hay calificaciones
- Silabo Adulto IIDocumento5 páginasSilabo Adulto IIanacervar6231Aún no hay calificaciones
- Sistema de Diagnostico PsicodinamicoDocumento33 páginasSistema de Diagnostico PsicodinamicoSilvia MoinoAún no hay calificaciones
- Familia ParvoviridaeDocumento23 páginasFamilia Parvoviridaealanytzeen100% (1)
- T4 Microbiota Obstetricia 23-I Prof. CuadraDocumento36 páginasT4 Microbiota Obstetricia 23-I Prof. CuadraMARJORIE BLANCA GARCIA TORPOCOAún no hay calificaciones
- Sistema Digestivo de AveDocumento27 páginasSistema Digestivo de AveDiego CastroAún no hay calificaciones
- AlprazolamDocumento7 páginasAlprazolamCristhian Vilchez RamosAún no hay calificaciones
- Caso 8Documento2 páginasCaso 8IXCHEL ESTRADA GONZALEZAún no hay calificaciones
- Fisioterapia Respiratoria - Cap 7. 2008Documento19 páginasFisioterapia Respiratoria - Cap 7. 2008Nacho OrtizAún no hay calificaciones
- Anatomia y Fisiologia Aparato CirculatorioDocumento17 páginasAnatomia y Fisiologia Aparato CirculatorioSantiago Gutiérrez.Aún no hay calificaciones
- Tuberculosis Juan Nicolás Calva CastilloDocumento5 páginasTuberculosis Juan Nicolás Calva CastilloNicolás CalvaAún no hay calificaciones
- Proyecto Ñuchco, Clases HoyDocumento24 páginasProyecto Ñuchco, Clases Hoybrigner cardenas azambujaAún no hay calificaciones
- Guía de La ExcreciónDocumento3 páginasGuía de La ExcreciónAngie Ruiz martinezAún no hay calificaciones
- Neumonia ConnatalDocumento8 páginasNeumonia Connatalfernando troya100% (1)
- Estudio de Caso Trastorno de Panico AdolecenteDocumento7 páginasEstudio de Caso Trastorno de Panico AdolecentepambriAún no hay calificaciones