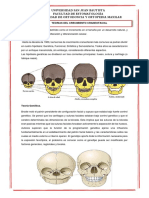
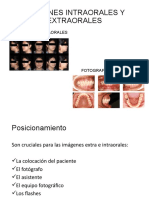
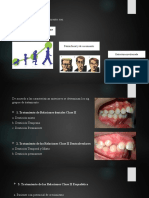
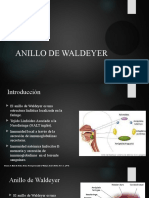
También podría gustarte
- Primer Taller de Crecimiento y DesarrolloDocumento7 páginasPrimer Taller de Crecimiento y DesarrolloVERONICA RIVAS LEIVAAún no hay calificaciones
- Teorías de Crecimiento CraneofacialDocumento10 páginasTeorías de Crecimiento Craneofacialalejandra gonzalezAún no hay calificaciones
- Teorias Del CrecimientoDocumento5 páginasTeorias Del CrecimientoITZEL EDITH CISNEROS CAMARENAAún no hay calificaciones
- Crecimiento y Desarrollo MaxilofacialDocumento35 páginasCrecimiento y Desarrollo MaxilofacialNelson Alvarado CorcueraAún no hay calificaciones
- Cap 2 Crec Mandib Pag 21-30Documento6 páginasCap 2 Crec Mandib Pag 21-30Pamela Peña MAún no hay calificaciones
- The Differential Roles of Periosteal and Capsular Functional - ESPAÑOLDocumento6 páginasThe Differential Roles of Periosteal and Capsular Functional - ESPAÑOLDayana MonroyAún no hay calificaciones
- Postura - Craneocervical - MaloclusionesDocumento12 páginasPostura - Craneocervical - Maloclusionesluz100% (1)
- Teorias de Crecimiento y Desarrollo CraneofacialDocumento7 páginasTeorias de Crecimiento y Desarrollo Craneofacialkitzia yarumiAún no hay calificaciones
- Desarrollo OdontopediatriaDocumento13 páginasDesarrollo OdontopediatriaCristhian VelasteguiAún no hay calificaciones
- Teorías Del Crecimiento CraneofacialDocumento4 páginasTeorías Del Crecimiento CraneofacialEstefany Capuñay100% (1)
- 2 Thilander Mecanismo de Crecimiento ESAPAÑOLDocumento8 páginas2 Thilander Mecanismo de Crecimiento ESAPAÑOLIsa PérezAún no hay calificaciones
- Teorías Del Crecimiento CraneofacialDocumento10 páginasTeorías Del Crecimiento CraneofacialJose Luis Vergara SarmientoAún no hay calificaciones
- Teorias de Desarrollo y Crecimiento - Jorge Junior Sanga GárateDocumento5 páginasTeorias de Desarrollo y Crecimiento - Jorge Junior Sanga GárateS.g. JorgeAún no hay calificaciones
- Thilander ESpDocumento10 páginasThilander ESpYady S. Londoño L.Aún no hay calificaciones
- Seminario Ortopedia IDocumento11 páginasSeminario Ortopedia IDr. JharAún no hay calificaciones
- The Primary Role of Functional Matrices in Facial Growth (2) Es (1) - 1Documento12 páginasThe Primary Role of Functional Matrices in Facial Growth (2) Es (1) - 1Nahomy MendozaAún no hay calificaciones
- Crecimiento y Desarrollo Craneofacial PostnatalDocumento47 páginasCrecimiento y Desarrollo Craneofacial PostnatalCristobal Escobar100% (1)
- Estudio de los huesos y su modificación cultural en poblaciones antiguasDocumento24 páginasEstudio de los huesos y su modificación cultural en poblaciones antiguasDeniss Garcia75% (4)
- Centros y Lugares de Crecimiento (Autoguardado)Documento20 páginasCentros y Lugares de Crecimiento (Autoguardado)Brian RodAún no hay calificaciones
- Kevin George Vasquez Bardales Infografia Teoria de MossDocumento1 páginaKevin George Vasquez Bardales Infografia Teoria de MossGeorge Vásquez BardalesAún no hay calificaciones
- Crecimiento y Desarrollo Del Sistema EstomatognáticoDocumento12 páginasCrecimiento y Desarrollo Del Sistema EstomatognáticoRoneida Chirinos50% (2)
- Moss Ana CDocumento24 páginasMoss Ana CJean Jose Orozco Trujillo100% (3)
- Crecimiento CraneofacialDocumento52 páginasCrecimiento CraneofacialManiiki100% (1)
- TeoriasDocumento5 páginasTeoriasortodoncia 2018Aún no hay calificaciones
- Crecimiento Vista EsquemaDocumento5 páginasCrecimiento Vista Esquemaviena100% (1)
- Teorías de Crecimiento Craneofacial PDFDocumento2 páginasTeorías de Crecimiento Craneofacial PDFValentina Licona NuñezAún no hay calificaciones
- Teorias de Crecimiento y Desarrollo CraneofacialDocumento11 páginasTeorias de Crecimiento y Desarrollo CraneofacialEver IspilcoAún no hay calificaciones
- Desarrollo Neural y MotorDocumento3 páginasDesarrollo Neural y MotorFernanda Beatriz Paredes Abarca 31Aún no hay calificaciones
- Crecimiento y desarrollo del esqueleto craneofacialDocumento25 páginasCrecimiento y desarrollo del esqueleto craneofacialFrancisca HenriquezAún no hay calificaciones
- Pontificia Universidad Catolica de Chile Facultad de Medicina Carrera de Odontología Clínica Integral Del Niño ODO 502 ADocumento24 páginasPontificia Universidad Catolica de Chile Facultad de Medicina Carrera de Odontología Clínica Integral Del Niño ODO 502 AMarcela Aroca ChandiaAún no hay calificaciones
- Guia de OrtodonciaDocumento19 páginasGuia de OrtodonciaPatricia ValentinaAún no hay calificaciones
- Características Biométricas Del Cuerpo CallosoDocumento4 páginasCaracterísticas Biométricas Del Cuerpo CallosoJorge MarquetAún no hay calificaciones
- Trabajo 1 Motricidad OralDocumento8 páginasTrabajo 1 Motricidad OralYely GuarateAún no hay calificaciones
- Crecimiento y Desarrollo Del Esqueleto CraneofacialDocumento7 páginasCrecimiento y Desarrollo Del Esqueleto CraneofacialAgustina AlbertengoAún no hay calificaciones
- Teorías del crecimiento craneofacialDocumento8 páginasTeorías del crecimiento craneofacialSuldeny TobónAún no hay calificaciones
- Control morfogénico del crecimiento craneofacialDocumento6 páginasControl morfogénico del crecimiento craneofacialPaola VasquezAún no hay calificaciones
- Crecimiento Preguntas DesarrolloDocumento3 páginasCrecimiento Preguntas DesarrolloDeyvid Gustavo Capelo RiosAún no hay calificaciones
- Seminario Crecimiento Postnatal y AdultoDocumento41 páginasSeminario Crecimiento Postnatal y Adultoelias100% (1)
- El Papel Principal de La Función Matrices en El Crecimiento FacialDocumento11 páginasEl Papel Principal de La Función Matrices en El Crecimiento FacialDayana MonroyAún no hay calificaciones
- 2022 4740 1 PB PDFDocumento11 páginas2022 4740 1 PB PDFMiguel Angel Mendoza CastroAún no hay calificaciones
- Teorias Del Crecimiento Craneofacial UnaDocumento11 páginasTeorias Del Crecimiento Craneofacial UnaEvelyn SegoviaAún no hay calificaciones
- Las Teorías Del Crecimiento CraneofacialDocumento2 páginasLas Teorías Del Crecimiento CraneofacialJhamilAún no hay calificaciones
- Teorías Del CrecimientoDocumento34 páginasTeorías Del CrecimientoJohann SolSán100% (1)
- Teorías Del CrecimientoDocumento8 páginasTeorías Del CrecimientoJaime Luis Guamán Quintuña100% (1)
- Moss 1969 TraducidoDocumento13 páginasMoss 1969 TraducidoEstefany SaldañaAún no hay calificaciones
- Indicaciones y Efectos Fisiologicos de Las Tecnicas Manipulativas CranealesDocumento11 páginasIndicaciones y Efectos Fisiologicos de Las Tecnicas Manipulativas CranealesCarlos Marques100% (1)
- Monografía 27 de NoviembreDocumento52 páginasMonografía 27 de NoviembreAnaAún no hay calificaciones
- Morfometria Geometrica Aplicada Al EstudDocumento20 páginasMorfometria Geometrica Aplicada Al EstudAdolfo QteletAún no hay calificaciones
- Generalidades de Anatomia para EstudiantesDocumento17 páginasGeneralidades de Anatomia para EstudiantesAlex FernandezAún no hay calificaciones
- Teorias Del CrecimientoDocumento5 páginasTeorias Del CrecimientoDaniela Robledo MontijoAún no hay calificaciones
- Fundamentos biomecánicos del complejo del hombroDocumento20 páginasFundamentos biomecánicos del complejo del hombroartgpsAún no hay calificaciones
- Teorias de Crecimiento CraneofacialDocumento10 páginasTeorias de Crecimiento CraneofacialVictoriaNavaLeyvaAún no hay calificaciones
- Crecimiento y Desarrollo - OdontologíaDocumento8 páginasCrecimiento y Desarrollo - OdontologíaMaria BisvalAún no hay calificaciones
- La importancia del análisis postural en pediatríaDocumento24 páginasLa importancia del análisis postural en pediatríaCentro Medico e Dentario FaroAún no hay calificaciones
- Estudio3.Pdfanalisis de RocabadoDocumento10 páginasEstudio3.Pdfanalisis de RocabadoJuan Simon100% (2)
- Teoria de MossDocumento10 páginasTeoria de MossAlan Roberto Flores GutierrezAún no hay calificaciones
- Fuerza muscular, funcionalidad y envejecimientoDe EverandFuerza muscular, funcionalidad y envejecimientoCalificación: 5 de 5 estrellas5/5 (2)
- Disfunción Cráneo Cérvico MandibularDe EverandDisfunción Cráneo Cérvico MandibularCalificación: 3 de 5 estrellas3/5 (3)
- Cinesiología: Alteraciones tónicas (Color)De EverandCinesiología: Alteraciones tónicas (Color)Calificación: 4.5 de 5 estrellas4.5/5 (4)
- Fuerza óptima para movimiento dental ortodóncicoDocumento6 páginasFuerza óptima para movimiento dental ortodóncicoAriadna MéndezAún no hay calificaciones
- Asociación Del Sistema de Fuerza de Ortodoncia y La Reabsorción RadicularDocumento8 páginasAsociación Del Sistema de Fuerza de Ortodoncia y La Reabsorción RadicularAriadna MéndezAún no hay calificaciones
- Hialinización y Reabsorción Radicular Durante El Movimiento Dentario Ortodóncico Temprano en AdolescentesDocumento3 páginasHialinización y Reabsorción Radicular Durante El Movimiento Dentario Ortodóncico Temprano en AdolescentesAriadna MéndezAún no hay calificaciones
- Cambios en El Tejido Incidentales Al Movimiento de Los Dientes OrtodóncicosDocumento7 páginasCambios en El Tejido Incidentales Al Movimiento de Los Dientes OrtodóncicosAriadna MéndezAún no hay calificaciones
- Hialinización Durante El Movimiento Dental OrtodóncicoDocumento5 páginasHialinización Durante El Movimiento Dental OrtodóncicoAriadna MéndezAún no hay calificaciones
- La Importancia de La Posición Del Incisivo en La Sonrisa EstéticaDocumento12 páginasLa Importancia de La Posición Del Incisivo en La Sonrisa EstéticaAriadna MéndezAún no hay calificaciones
- Imágenes Intraorales y ExtraoralesDocumento12 páginasImágenes Intraorales y ExtraoralesAriadna MéndezAún no hay calificaciones
- Microtomografía Basada en Radiación Sincrotrón de Tejidos de Soporte AlveolarDocumento4 páginasMicrotomografía Basada en Radiación Sincrotrón de Tejidos de Soporte AlveolarAriadna MéndezAún no hay calificaciones
- El Uso de Ángulos Incisales en La Cefalometría SteinerDocumento9 páginasEl Uso de Ángulos Incisales en La Cefalometría SteinerAriadna MéndezAún no hay calificaciones
- Linfáticos de La CabezaDocumento3 páginasLinfáticos de La CabezaAriadna MéndezAún no hay calificaciones
- Resumen Ángulos y FotosDocumento5 páginasResumen Ángulos y FotosAriadna MéndezAún no hay calificaciones
- Perspectivas en La Aplicación Clínica de La Cefalometría EnsayoDocumento18 páginasPerspectivas en La Aplicación Clínica de La Cefalometría EnsayoAriadna MéndezAún no hay calificaciones
- NUEVO Maloclusion Clase !!Documento17 páginasNUEVO Maloclusion Clase !!Ariadna MéndezAún no hay calificaciones
- Reacción Tisular Al Movimiento Dental OrtodóncicoDocumento7 páginasReacción Tisular Al Movimiento Dental OrtodóncicoAriadna Méndez100% (1)
- Raymond Begg adaptó el arco de cintaDocumento2 páginasRaymond Begg adaptó el arco de cintaAriadna MéndezAún no hay calificaciones
- Nervio facial: anatomía, trayecto y ramos del VII par cranealDocumento22 páginasNervio facial: anatomía, trayecto y ramos del VII par cranealAriadna MéndezAún no hay calificaciones
- Anillo WaldeyerDocumento13 páginasAnillo WaldeyerAriadna MéndezAún no hay calificaciones
- Ramos Colaterales IntrapetrososDocumento12 páginasRamos Colaterales IntrapetrososAriadna MéndezAún no hay calificaciones
- LinfangitisDocumento19 páginasLinfangitisAriadna MéndezAún no hay calificaciones
- Ramos Aferentes y Eferentes de Éstos GangliosDocumento7 páginasRamos Aferentes y Eferentes de Éstos GangliosAriadna MéndezAún no hay calificaciones
- Maloclusion Clase 2 Parte 4Documento11 páginasMaloclusion Clase 2 Parte 4Ariadna MéndezAún no hay calificaciones
- Análisis CefalométricoDocumento7 páginasAnálisis CefalométricoAriadna MéndezAún no hay calificaciones
- Ramos Aferentes y Eferentes de Éstos GangliosDocumento7 páginasRamos Aferentes y Eferentes de Éstos GangliosAriadna MéndezAún no hay calificaciones
- AdenopatíaDocumento7 páginasAdenopatíaAriadna MéndezAún no hay calificaciones
- Linfáticos de La CabezaDocumento3 páginasLinfáticos de La CabezaAriadna MéndezAún no hay calificaciones
- AdenopatíaDocumento7 páginasAdenopatíaAriadna MéndezAún no hay calificaciones
- Prevalencia de Variables AnatómicasDocumento10 páginasPrevalencia de Variables AnatómicasAriadna MéndezAún no hay calificaciones
- Análisis Cefalométrico 2Documento4 páginasAnálisis Cefalométrico 2Ariadna MéndezAún no hay calificaciones
- Ramos Colaterales IntrapetrososDocumento12 páginasRamos Colaterales IntrapetrososAriadna MéndezAún no hay calificaciones
- Variaciones Del Canal Mandibular Con Importancia Clínica Reporte de CasoDocumento4 páginasVariaciones Del Canal Mandibular Con Importancia Clínica Reporte de CasoMario Zúñiga GonzalesAún no hay calificaciones
- SST - FormatosDocumento17 páginasSST - FormatosMichel Bonilla BedoyaAún no hay calificaciones
- Guia2 Lab Ciclo 02-2021Documento14 páginasGuia2 Lab Ciclo 02-2021Héctor PerezAún no hay calificaciones
- Componentes Del SueloDocumento6 páginasComponentes Del SueloDiego Alessandro Marquez100% (1)
- Derecho Administrativo Sancionador en BoliviaDocumento75 páginasDerecho Administrativo Sancionador en BoliviaMario VillegasAún no hay calificaciones
- Indices Troficos DFDocumento34 páginasIndices Troficos DFFernandoAún no hay calificaciones
- Informe-Wais - Caso R.M.V. 19 AñosDocumento6 páginasInforme-Wais - Caso R.M.V. 19 AñosCamila PastorAún no hay calificaciones
- Proyecto Final Biologia 2Documento7 páginasProyecto Final Biologia 2Jordan AguilarAún no hay calificaciones
- Soluciones Ejercicios Derivadas (Final Tema) PDFDocumento11 páginasSoluciones Ejercicios Derivadas (Final Tema) PDFsantiago cantorAún no hay calificaciones
- 20201029-121009-La Biodiversidad en El PeruDocumento4 páginas20201029-121009-La Biodiversidad en El PeruJosh BecaAún no hay calificaciones
- Factores que afectan la gestión empresarialDocumento8 páginasFactores que afectan la gestión empresarialMauricio Andrés Ramirez BarcoAún no hay calificaciones
- 3.1 Marco Histórico y TeóricoDocumento10 páginas3.1 Marco Histórico y Teóricodianaverdinb5Aún no hay calificaciones
- PLANIFICACIÓN A CORTO PLAZO 17 DE AGOSTO Lenguaje ArtDocumento3 páginasPLANIFICACIÓN A CORTO PLAZO 17 DE AGOSTO Lenguaje ArtFranciscaBelénUrbinaMartínezAún no hay calificaciones
- Plan Clase TRDocumento7 páginasPlan Clase TRMarcelo VilemaAún no hay calificaciones
- Capítulo 1 - Importancia de La Biología Celular y Tisular y Técnicas de Estudio de Células y TejidosDocumento21 páginasCapítulo 1 - Importancia de La Biología Celular y Tisular y Técnicas de Estudio de Células y TejidosAlexandra AndradeAún no hay calificaciones
- EnacomDocumento11 páginasEnacomCrónicaAún no hay calificaciones
- p1 - Teoría de Exponentes y Ecuaciones Exponenciales - Cpu Fase Cero 2021 - IDocumento2 páginasp1 - Teoría de Exponentes y Ecuaciones Exponenciales - Cpu Fase Cero 2021 - IMagui MajuAún no hay calificaciones
- Palabras homófonasDocumento6 páginasPalabras homófonasAlex CortesAún no hay calificaciones
- Resolución de oraciones incompletas con criterios de precisión léxica, gramaticalidad y coherencia textualDocumento28 páginasResolución de oraciones incompletas con criterios de precisión léxica, gramaticalidad y coherencia textualYarome MendozaAún no hay calificaciones
- Perfil PuestosDocumento46 páginasPerfil Puestoschris bernabel100% (1)
- Ciclos de Refrigeración y RefrigerantesDocumento8 páginasCiclos de Refrigeración y RefrigerantesCarlos ArancibiaAún no hay calificaciones
- Lineamientos TA1 Comportamiento Al Consumidor - 2021Documento7 páginasLineamientos TA1 Comportamiento Al Consumidor - 2021José GarcíaAún no hay calificaciones
- Manual de Bolsillo EterboardDocumento44 páginasManual de Bolsillo EterboardFernando QuirozAún no hay calificaciones
- Actividad 2 - SonidoDocumento1 páginaActividad 2 - Sonidozulmaqv1Aún no hay calificaciones
- El Problema Social Del Indio (1923 y 1971)Documento4 páginasEl Problema Social Del Indio (1923 y 1971)Pierre CollazosAún no hay calificaciones
- Organo de La VisionDocumento33 páginasOrgano de La Visionjunior pinedaAún no hay calificaciones
- Te 1319Documento45 páginasTe 1319erlik aironAún no hay calificaciones
- Movimiento en Una Dimensión Con MRUADocumento16 páginasMovimiento en Una Dimensión Con MRUAElias NZGAún no hay calificaciones
- Sena - Trabajo 2Documento6 páginasSena - Trabajo 2santiago duque cundumiAún no hay calificaciones
- Gobernanza Forestal en Brasil Colombia Ecuador PeruDocumento9 páginasGobernanza Forestal en Brasil Colombia Ecuador PeruAitana SabatoAún no hay calificaciones
- Seguidor de Linea Dispo555Documento6 páginasSeguidor de Linea Dispo555Joseph Moreno GutierrezAún no hay calificaciones