También podría gustarte
- Fundamentos y técnicas básicas en biología molecularDe EverandFundamentos y técnicas básicas en biología molecularAún no hay calificaciones
- Verde Tema de La Clase Electricidad Simple Minimalista PresentaciónDocumento63 páginasVerde Tema de La Clase Electricidad Simple Minimalista PresentaciónNashira NhaAún no hay calificaciones
- Replicacion Del Adn Guía de AprendizajeDocumento5 páginasReplicacion Del Adn Guía de AprendizajeAngieSantiAún no hay calificaciones
- Replicacion Del ADNDocumento16 páginasReplicacion Del ADNbarbara belen escobar jaraAún no hay calificaciones
- Replicacion de ADNDocumento9 páginasReplicacion de ADNmamenmarinAún no hay calificaciones
- Modelos de Replicación Del ADNDocumento8 páginasModelos de Replicación Del ADNFernando RamirezAún no hay calificaciones
- Tema 10 Replic Transc TraducDocumento14 páginasTema 10 Replic Transc TraducAlexander PinzonAún no hay calificaciones
- Clases Tema 10Documento12 páginasClases Tema 10UrielGranadosAún no hay calificaciones
- Replicacio Del AdnDocumento8 páginasReplicacio Del Adnpaola rojasAún no hay calificaciones
- Replicacion Del AdnDocumento7 páginasReplicacion Del AdnAngelicaMoranAún no hay calificaciones
- Replicación y Reparación Del DnaDocumento21 páginasReplicación y Reparación Del DnaAlbert AznaranAún no hay calificaciones
- Replicación de ADNDocumento11 páginasReplicación de ADNRene Gtz SAún no hay calificaciones
- Tipos de Replicación Del ADNDocumento5 páginasTipos de Replicación Del ADNDavidAún no hay calificaciones
- Experiment OsDocumento46 páginasExperiment OsLuz M. MolinaAún no hay calificaciones
- Investigacion Documental ReplicacionDocumento5 páginasInvestigacion Documental ReplicacionEmmanuel GutiérrezAún no hay calificaciones
- Columbus University Facultad de Medicina y Ciencia de La Salud Curso de BiologíaDocumento29 páginasColumbus University Facultad de Medicina y Ciencia de La Salud Curso de BiologíaManuel PeraltaAún no hay calificaciones
- Cuestionario Replicación Conceptos Básicos 1Documento7 páginasCuestionario Replicación Conceptos Básicos 1Vanessa SanabriaAún no hay calificaciones
- I5 ReplicacionDocumento36 páginasI5 ReplicacionHeidy TanguilaAún no hay calificaciones
- 4to Medio Guía 1 Biología - Replicación Del ADNDocumento6 páginas4to Medio Guía 1 Biología - Replicación Del ADNFran CárdenasAún no hay calificaciones
- Clase 12Documento40 páginasClase 12alfredoAún no hay calificaciones
- Cap 25Documento5 páginasCap 25Jazmín Valdiviezo HuamánAún no hay calificaciones
- Replicación UTPDocumento14 páginasReplicación UTPJudith San Martin0% (1)
- Universidad Nacional Del Altiplano-Puno Facultad de Ciencias Agrarias Escuela Profesional de Ingeniería AgronómicaDocumento10 páginasUniversidad Nacional Del Altiplano-Puno Facultad de Ciencias Agrarias Escuela Profesional de Ingeniería Agronómicasergio felix pacosonccoAún no hay calificaciones
- Replicación de ADNDocumento68 páginasReplicación de ADNlucia janet100% (1)
- Replicación de ADNDocumento4 páginasReplicación de ADNNayeli HernándezAún no hay calificaciones
- Replicación de ADN BAmbDocumento13 páginasReplicación de ADN BAmbMaldonado López GabrielaAún no hay calificaciones
- Replicación Del ADNDocumento5 páginasReplicación Del ADNMarisol TorresAún no hay calificaciones
- Dogma Central de La BiologíaDocumento16 páginasDogma Central de La Biologíajunior alarconAún no hay calificaciones
- Biologia 101Documento13 páginasBiologia 101EDUARDO ESQUIVELAún no hay calificaciones
- 01 02 BIO-002 U03 Replicacià NDocumento14 páginas01 02 BIO-002 U03 Replicacià NIgnacio CortesAún no hay calificaciones
- 2012 t13 Genetica MolecularDocumento40 páginas2012 t13 Genetica MolecularAnna MallorquínAún no hay calificaciones
- Guía Replicación Del ADNDocumento2 páginasGuía Replicación Del ADNAndres Oyanadel CortesAún no hay calificaciones
- Lectura Sobre La Replicación El ADNDocumento24 páginasLectura Sobre La Replicación El ADNNohelia DavilaAún no hay calificaciones
- Fragmento de OkasakiDocumento8 páginasFragmento de OkasakiYaritza Anzaldo PérezAún no hay calificaciones
- VI DNA Estructura y FunciónDocumento72 páginasVI DNA Estructura y FunciónXsiSkullAún no hay calificaciones
- La Replicación: Universidad Nacional Jorge Basadre GrohmannDocumento25 páginasLa Replicación: Universidad Nacional Jorge Basadre GrohmannEmmanuel Escalera RosalesAún no hay calificaciones
- 12 - Genética Molecular IDocumento17 páginas12 - Genética Molecular IMiguel SanchezAún no hay calificaciones
- Ciclo Del CloroDocumento13 páginasCiclo Del Clorojoel freireAún no hay calificaciones
- Traduccion Del ADNDocumento6 páginasTraduccion Del ADNDavid ChandiAún no hay calificaciones
- Replicacion ResumenDocumento5 páginasReplicacion ResumenCielo MataAún no hay calificaciones
- Etica y Deontologia MedicaDocumento8 páginasEtica y Deontologia Medicajnorberto876Aún no hay calificaciones
- REPLICACIONDocumento56 páginasREPLICACIONXimena CajamarcaAún no hay calificaciones
- TEMA 7 Biología MolecularDocumento27 páginasTEMA 7 Biología MolecularRocio MAún no hay calificaciones
- Guión GB Tema 3Documento2 páginasGuión GB Tema 3antonio garcia martinezAún no hay calificaciones
- Diapositivas de Biologia ReplicacionDocumento43 páginasDiapositivas de Biologia Replicacionmajo gsAún no hay calificaciones
- GenomicaDocumento33 páginasGenomicaCarlos Segovia DominguezAún no hay calificaciones
- Guia de Complemento de ContenidosDocumento8 páginasGuia de Complemento de ContenidosAntonia OñateAún no hay calificaciones
- Replicación Del DNA 2022Documento101 páginasReplicación Del DNA 2022jeniferAún no hay calificaciones
- Bases de La Replicacion Del Adn1Documento25 páginasBases de La Replicacion Del Adn1DanitzaDelRosarioMenesesBardalesAún no hay calificaciones
- REPLICA AD ONCE 2021docxDocumento11 páginasREPLICA AD ONCE 2021docxpaula mendozaAún no hay calificaciones
- Replicación Del DnaDocumento11 páginasReplicación Del DnaMirna Aranda BarbozaAún no hay calificaciones
- Seminario G1 - Replicación Del DNADocumento11 páginasSeminario G1 - Replicación Del DNAABRIL SOFIA CALDERON TERANAún no hay calificaciones
- Características Generales Del ADNDocumento3 páginasCaracterísticas Generales Del ADNyeagerAún no hay calificaciones
- Dia Replicacion Del AdnDocumento13 páginasDia Replicacion Del Adnjoel freireAún no hay calificaciones
- 3 Replicación y Mecanismos de Reparación Del ADN (2021 PS) - Clase 3Documento173 páginas3 Replicación y Mecanismos de Reparación Del ADN (2021 PS) - Clase 3KATALINA IGNACIA AZÓCARAún no hay calificaciones
- Replicación de ADN en Células EucariotasDocumento16 páginasReplicación de ADN en Células EucariotasAbigail MerazAún no hay calificaciones
- 116200603037Documento8 páginas116200603037luzmarinaAún no hay calificaciones
- Guicc81a nc2b0 4 4c2ba Medio Comucc81n Replicaciocc81n 2Documento6 páginasGuicc81a nc2b0 4 4c2ba Medio Comucc81n Replicaciocc81n 2Daniela PerezAún no hay calificaciones
- REPLICACION DEL ADN Equipo 3Documento7 páginasREPLICACION DEL ADN Equipo 3Kengar De La SelvaAún no hay calificaciones
- La Comunidad Científica Había Propuesto Tres Modelos Básicos Sobre La Replicación Del ADN Después Del Descubrimiento de La Estructura Del ADNDocumento2 páginasLa Comunidad Científica Había Propuesto Tres Modelos Básicos Sobre La Replicación Del ADN Después Del Descubrimiento de La Estructura Del ADNAntenita 01Aún no hay calificaciones
- CAPÍTULO 13 - Replicación y Reparación Del DNADocumento45 páginasCAPÍTULO 13 - Replicación y Reparación Del DNALaura Diaz ChaverraAún no hay calificaciones
- AcentuaciónDocumento3 páginasAcentuaciónLaura Diaz ChaverraAún no hay calificaciones
- CAPÍTULO 13 - Replicación y Reparación Del DNADocumento45 páginasCAPÍTULO 13 - Replicación y Reparación Del DNALaura Diaz ChaverraAún no hay calificaciones
- 4 Esp Sintomatologia Derivada de Los Pares Craneales Clinica y Topografia Radiologia 2019Documento25 páginas4 Esp Sintomatologia Derivada de Los Pares Craneales Clinica y Topografia Radiologia 2019Laura Diaz ChaverraAún no hay calificaciones
- 7671 26022 1 PBDocumento9 páginas7671 26022 1 PBAlejandro GomezAún no hay calificaciones
- Articulopublicado 2Documento16 páginasArticulopublicado 2Laura Diaz ChaverraAún no hay calificaciones
- Patologia Pares 1Documento13 páginasPatologia Pares 1Alan SaucedoAún no hay calificaciones
- Exploracion Neurologica y Atencion PrimariaDocumento10 páginasExploracion Neurologica y Atencion Primariasally000000Aún no hay calificaciones
- Complejo Mayor de Histocompatibilidad (MHC)Documento17 páginasComplejo Mayor de Histocompatibilidad (MHC)Luis DiazzAún no hay calificaciones
- Programa Curso BioinformaticaDocumento1 páginaPrograma Curso Bioinformaticajuan1401Aún no hay calificaciones
- Plastos y CloroplastosDocumento10 páginasPlastos y CloroplastosPaola SerratoAún no hay calificaciones
- Biología Molecular y BioquímicaDocumento8 páginasBiología Molecular y BioquímicaAri FernandezAún no hay calificaciones
- Examen FisiologiaDocumento5 páginasExamen FisiologiaMonica ReyesAún no hay calificaciones
- 15 - Regulación II - AlosterismoDocumento4 páginas15 - Regulación II - AlosterismoferminAún no hay calificaciones
- Fga - 23-2 Biología MolecularDocumento16 páginasFga - 23-2 Biología MolecularJhon Sundose FilesAún no hay calificaciones
- Via de Las Pentosas y GlutationDocumento16 páginasVia de Las Pentosas y GlutationAlexander Salinas CalderónAún no hay calificaciones
- 6 Transporte CelularDocumento9 páginas6 Transporte Celularmateo alvaradoAún no hay calificaciones
- Composición Química de Los Seres VivosDocumento22 páginasComposición Química de Los Seres VivosRAMIRO GAEL SAHAGUN IBARRAAún no hay calificaciones
- Replicación de ADN BAmbDocumento13 páginasReplicación de ADN BAmbMaldonado López GabrielaAún no hay calificaciones
- Bloque1 2Documento23 páginasBloque1 2Telmo Olivas SanchesAún no hay calificaciones
- Semana 3 y 4 - DécimoDocumento2 páginasSemana 3 y 4 - DécimoVilma Eduviges Roca Bosquez100% (1)
- Vectores de ClonaciónDocumento3 páginasVectores de Clonaciónselenelemus259Aún no hay calificaciones
- SECUENCIA DE REACCIONES DE FinalDocumento40 páginasSECUENCIA DE REACCIONES DE FinalCristhian BeizagaAún no hay calificaciones
- Núcleo y NucleoloDocumento1 páginaNúcleo y NucleoloValeria Karolina Ochoa CazaresAún no hay calificaciones
- Informe PrionesDocumento44 páginasInforme PrionesJose Arias Ayala100% (1)
- Tema 12..... Boletín..... 04-09-2019Documento7 páginasTema 12..... Boletín..... 04-09-2019wilmanAún no hay calificaciones
- Ramas de La BiologiaDocumento6 páginasRamas de La BiologiaTEPACH COBAXIN LENCY MARIAAún no hay calificaciones
- Diferencia de La Replicación Entre Procariotas y EucariotasDocumento5 páginasDiferencia de La Replicación Entre Procariotas y EucariotasVanessa100% (1)
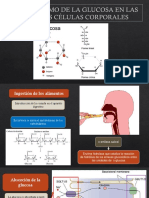
- Metabolismo de La Glucosa en Las Diversas Células Corporales e Integración Del Mantenimiento de Los Niveles de GlucosaDocumento24 páginasMetabolismo de La Glucosa en Las Diversas Células Corporales e Integración Del Mantenimiento de Los Niveles de GlucosaKevin Minchola CastañedaAún no hay calificaciones
- Recepción Celular. Mecanismos MolécularesDocumento24 páginasRecepción Celular. Mecanismos MolécularesTEFAAún no hay calificaciones
- Inmunoglobulinas y AnticuerposDocumento20 páginasInmunoglobulinas y Anticuerposmichel100% (1)
- Informe de Las ProteínasDocumento7 páginasInforme de Las ProteínasEmanuel SáezAún no hay calificaciones
- Biomoleculas III (Carbohidratos, Metabolismo, Ciclo de Krebs)Documento55 páginasBiomoleculas III (Carbohidratos, Metabolismo, Ciclo de Krebs)CAMILA SUAZAAún no hay calificaciones
- 2medio Repasando1medioguiadeaprendizajeDocumento6 páginas2medio Repasando1medioguiadeaprendizajeNatalia Arias FigueroaAún no hay calificaciones
- Marco Teórico Transporte de Membranas - ÓsmosisDocumento4 páginasMarco Teórico Transporte de Membranas - ÓsmosisNuñez BrayanAún no hay calificaciones
- Organelos CelulalesDocumento7 páginasOrganelos CelulalesKIMBERLY BETZABETH VELARDE CORONADOAún no hay calificaciones
- Conductos y Bombas de MembranaDocumento20 páginasConductos y Bombas de MembranaGaby Lisbeth Llh100% (1)
- Infografía Sobre Insulina y Receptor Tirosina SinasaDocumento2 páginasInfografía Sobre Insulina y Receptor Tirosina Sinasajesus batlliaAún no hay calificaciones
- Homo antecessor: El nacimiento de una especieDe EverandHomo antecessor: El nacimiento de una especieCalificación: 5 de 5 estrellas5/5 (1)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Neuropsicología: Los fundamentos de la materiaDe EverandNeuropsicología: Los fundamentos de la materiaCalificación: 5 de 5 estrellas5/5 (1)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- Nuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosDe EverandNuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosCalificación: 3.5 de 5 estrellas3.5/5 (6)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Cerebros rotos: Pacientes asombrosos que me enseñaron a vivirDe EverandCerebros rotos: Pacientes asombrosos que me enseñaron a vivirCalificación: 5 de 5 estrellas5/5 (3)
- Sacale partido a tu cerebro: Todo lo que necesitas saber para mejorar tu memoria, tomar decisiones y aprovechar todo tu potencialDe EverandSacale partido a tu cerebro: Todo lo que necesitas saber para mejorar tu memoria, tomar decisiones y aprovechar todo tu potencialCalificación: 3 de 5 estrellas3/5 (1)
- La belleza tiene su ciencia: Mitos y verdades sobre los cosméticos, el cuidado de la piel y el cabelloDe EverandLa belleza tiene su ciencia: Mitos y verdades sobre los cosméticos, el cuidado de la piel y el cabelloCalificación: 5 de 5 estrellas5/5 (2)
- Terapia de vidas pasadas: Un camino hacia la luz del alma. Técnica y prácticaDe EverandTerapia de vidas pasadas: Un camino hacia la luz del alma. Técnica y prácticaCalificación: 4.5 de 5 estrellas4.5/5 (11)
- El cerebro del triunfador: 8 estrategias de las grandes mentes para alcanzar el éxitoDe EverandEl cerebro del triunfador: 8 estrategias de las grandes mentes para alcanzar el éxitoCalificación: 5 de 5 estrellas5/5 (1)
- La biología no miente: Revolución en salud. Basada en las cinco leyes biológicas descubiertas por el Dr. HamerDe EverandLa biología no miente: Revolución en salud. Basada en las cinco leyes biológicas descubiertas por el Dr. HamerCalificación: 4 de 5 estrellas4/5 (4)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Minerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.De EverandMinerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.Calificación: 3 de 5 estrellas3/5 (2)
- Inteligencia artificial: Una exploración filosófica sobre el futuro de la mente y la concienciaDe EverandInteligencia artificial: Una exploración filosófica sobre el futuro de la mente y la concienciaAna Isabel Sánchez DíezCalificación: 4 de 5 estrellas4/5 (3)
- Sesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesDe EverandSesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Los cinco ritos tibetanos de la eterna juventud: El ojo de la revelaciónDe EverandLos cinco ritos tibetanos de la eterna juventud: El ojo de la revelaciónCalificación: 4.5 de 5 estrellas4.5/5 (12)
- Curso Práctico de Química GeneralDe EverandCurso Práctico de Química GeneralCalificación: 4 de 5 estrellas4/5 (4)