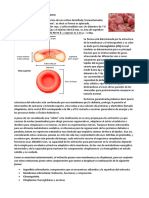
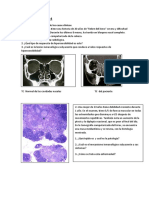
También podría gustarte
- Operación Atego - A9585841497-B (0315) PDFDocumento310 páginasOperación Atego - A9585841497-B (0315) PDFbeluAún no hay calificaciones
- Mayo Clinica Medicina InternaDocumento85 páginasMayo Clinica Medicina InternaDiana Zavala AlegreAún no hay calificaciones
- La Guía AnteriorDocumento3 páginasLa Guía AnteriorLaura Nieto100% (1)
- Sindrome de Consolidacion Pulmonar.Documento18 páginasSindrome de Consolidacion Pulmonar.Luis Ernesto Acuña MendezAún no hay calificaciones
- Retiro Nueva VidaDocumento3 páginasRetiro Nueva VidarubenAún no hay calificaciones
- DIMENSIONES Del Espectro AutistaDocumento12 páginasDIMENSIONES Del Espectro AutistaMarina Fleita100% (1)
- Examen Final md2n1 Desarrollo Morfofisiologico Del Ser Humano T 376772 Downloable 1827373Documento11 páginasExamen Final md2n1 Desarrollo Morfofisiologico Del Ser Humano T 376772 Downloable 1827373Abraham Josue Sulca ChucoAún no hay calificaciones
- Estudio de cohorte sobre diseño de investigación en epidemiologíaDocumento8 páginasEstudio de cohorte sobre diseño de investigación en epidemiologíaNicolás QuintanillaAún no hay calificaciones
- Cuestionario de HematologíaDocumento9 páginasCuestionario de HematologíaHenderson Vasquez ArmasAún no hay calificaciones
- Clinica Médica B 2019-I ActualizadoDocumento37 páginasClinica Médica B 2019-I ActualizadoBryan Yucra GarciaAún no hay calificaciones
- Emociones en el cuerpoDocumento7 páginasEmociones en el cuerpoMaritza RodriguezAún no hay calificaciones
- Examen Parcial - SEGUNDO INTENTO PRIMER BLOQUE-MEDICINA PREVENTIVA - (GRUPO3)Documento9 páginasExamen Parcial - SEGUNDO INTENTO PRIMER BLOQUE-MEDICINA PREVENTIVA - (GRUPO3)Mario0% (1)
- Fisiopatologia de La Isquemia CerebralDocumento204 páginasFisiopatologia de La Isquemia CerebralIsrael VillarrealAún no hay calificaciones
- Neurocisticercosis 151016095807 Lva1 App6891 PDFDocumento25 páginasNeurocisticercosis 151016095807 Lva1 App6891 PDFMartin AlarconAún no hay calificaciones
- CASO CLÍNCO N 5 Dr. FARÍAS FINALDocumento21 páginasCASO CLÍNCO N 5 Dr. FARÍAS FINALChristian Castro SanchezAún no hay calificaciones
- Casos Clinicos SCADocumento19 páginasCasos Clinicos SCAJohnAún no hay calificaciones
- Fraccionamiento Celular Con Higado de VacaDocumento10 páginasFraccionamiento Celular Con Higado de VacaBraulio GarciaAún no hay calificaciones
- Silabo SemiologiaDocumento31 páginasSilabo SemiologiaSofía CelisAún no hay calificaciones
- Recién Nacido Con Episodio de Atragantamiento y CianosisDocumento42 páginasRecién Nacido Con Episodio de Atragantamiento y CianosisVicky Leyne Alcantara BecerraAún no hay calificaciones
- Vena Cava Superior DobleDocumento3 páginasVena Cava Superior DobleJuliana MurziAún no hay calificaciones
- Semana 10 Cariotipo HumanoDocumento30 páginasSemana 10 Cariotipo HumanoDavid Cj AcAún no hay calificaciones
- Les 2023Documento50 páginasLes 2023Fio Marely Vargas PumacayoAún no hay calificaciones
- Banco de Preguntas Fármaco 2Documento4 páginasBanco de Preguntas Fármaco 2Danny83lAún no hay calificaciones
- CidDocumento28 páginasCidJhoannamalia Aurich RojasAún no hay calificaciones
- Insuficiencia Ovarica y TesticularDocumento89 páginasInsuficiencia Ovarica y TesticularPepe El ChidoAún no hay calificaciones
- Etimologias MedicasDocumento22 páginasEtimologias MedicasVICTOR LANDETA HERNANDEZAún no hay calificaciones
- UNMSM Sílabo Patología Especial 2016Documento27 páginasUNMSM Sílabo Patología Especial 2016MarcoAntonio Amaro CAún no hay calificaciones
- REPASO DE PATOLOGIA 3 Segmento 1Documento41 páginasREPASO DE PATOLOGIA 3 Segmento 1Dianha RuizAún no hay calificaciones
- Exp. HematoDocumento8 páginasExp. HematoSanta MendezAún no hay calificaciones
- Carlos Magdiel Abreu MendozaDocumento5 páginasCarlos Magdiel Abreu MendozaCarlos Magdiel Abreu MendozaAún no hay calificaciones
- AcumetriaDocumento6 páginasAcumetriaInés Muhr RieraAún no hay calificaciones
- Síndrome de disfunción múltiple de órganosDocumento24 páginasSíndrome de disfunción múltiple de órganosGiordan Sanchez100% (1)
- 5.3 Examen Físico Cardiovascular - Percusión y AuscultaciónDocumento7 páginas5.3 Examen Físico Cardiovascular - Percusión y AuscultaciónFaze DonatelloAún no hay calificaciones
- Taller sobre hipersensibilidad e inmunidadDocumento10 páginasTaller sobre hipersensibilidad e inmunidadLuis Enrique Silva ValenciaAún no hay calificaciones
- Sistema Nervioso Central - Grupo 1Documento16 páginasSistema Nervioso Central - Grupo 1ARIANA GREEIS LUJAN REBAZAAún no hay calificaciones
- Biofisica 2019 Clase 1 Silabo 2019 IDocumento11 páginasBiofisica 2019 Clase 1 Silabo 2019 IPercy Jando Muñoz Ramos100% (1)
- Causalidad de La LepraDocumento6 páginasCausalidad de La LepraPatricia ArguelloAún no hay calificaciones
- Caso Clinico Sesión 8 Semana 14 Caso 13Documento2 páginasCaso Clinico Sesión 8 Semana 14 Caso 13Ana Luisa Guevara Prieto0% (1)
- Láminas Corazon HISTODocumento7 páginasLáminas Corazon HISTOAriana VargasAún no hay calificaciones
- Explorac Corazón y Sist Vascular 22 Mayo 20-22 2020Documento73 páginasExplorac Corazón y Sist Vascular 22 Mayo 20-22 2020Ismael Coronado Flores100% (1)
- Nº6 Teoría Genética Resuelto Semana 6 UpaoDocumento7 páginasNº6 Teoría Genética Resuelto Semana 6 UpaoMaría Gracia MachadoAún no hay calificaciones
- Uso de Opioides en El Tratamiento Del Dolor OncologicoDocumento8 páginasUso de Opioides en El Tratamiento Del Dolor OncologicoDiego MartínezAún no hay calificaciones
- MIR 2013 Comentado p06Documento1 páginaMIR 2013 Comentado p06CristopherJairEstupiñanVirúAún no hay calificaciones
- MúsculoDocumento23 páginasMúsculoClara46Aún no hay calificaciones
- Silabo Histologia UpsjbDocumento16 páginasSilabo Histologia UpsjbKatherine RosseAún no hay calificaciones
- RELACIÓN ENTRE TÉCNICA QUIRÚRGICA Y COMPLICACIONES EN APENDICITIS AGUDADocumento57 páginasRELACIÓN ENTRE TÉCNICA QUIRÚRGICA Y COMPLICACIONES EN APENDICITIS AGUDAConsuelo Delgado RodriguezAún no hay calificaciones
- Morfo Ii Grupo 01 - Embrio (S04)Documento19 páginasMorfo Ii Grupo 01 - Embrio (S04)KEYSI GIOVELA DELGADO CHUMACEROAún no hay calificaciones
- Laboratorio de FarmacologíaDocumento25 páginasLaboratorio de FarmacologíaNicky100% (1)
- Preguntas de Metabolismo de HierroDocumento3 páginasPreguntas de Metabolismo de HierroAle Roque100% (1)
- Porfirinas y HHDocumento20 páginasPorfirinas y HHPau Mesa100% (1)
- Caso Clinico 3Documento3 páginasCaso Clinico 3Julissa Soto VelasquezAún no hay calificaciones
- Lógica JurídicaDocumento16 páginasLógica JurídicaeduardoAún no hay calificaciones
- Clasificación AntibióticosDocumento4 páginasClasificación AntibióticosLAURA VALENTINA RODRIGUEZ ROJASAún no hay calificaciones
- Caso Clínico Corregido U2M2T7Documento10 páginasCaso Clínico Corregido U2M2T7Adriana Ximena Mendoza MelendezAún no hay calificaciones
- Farmacología Informe N9Documento26 páginasFarmacología Informe N9Natali Thalia LongaAún no hay calificaciones
- Espacios PerifaringeosDocumento2 páginasEspacios PerifaringeosMaria PiñaAún no hay calificaciones
- Guia 2016Documento75 páginasGuia 2016VILLANUEVAMARCELAAún no hay calificaciones
- G.P. Diagnóstico Por Imágenes 2023-IDocumento36 páginasG.P. Diagnóstico Por Imágenes 2023-ICamille KirschbaumAún no hay calificaciones
- Resumen de FarmacologiaDocumento418 páginasResumen de FarmacologiaRafael Jiménez100% (5)
- Metabolismo AguaDocumento17 páginasMetabolismo AguaMaria Sallari GutiérrezAún no hay calificaciones
- Autoinstructivo 4Documento3 páginasAutoinstructivo 4Naomi CarreraAún no hay calificaciones
- Conceptos Básicos y Exploración: PsicopatologíaDocumento122 páginasConceptos Básicos y Exploración: PsicopatologíaKimberly Echaiz ibarraAún no hay calificaciones
- Taller N°2 - Elementos Base Del APOCDocumento6 páginasTaller N°2 - Elementos Base Del APOCmit CasAún no hay calificaciones
- 1 Silabo Primeros Auxilios 2021-2Documento5 páginas1 Silabo Primeros Auxilios 2021-2Mena NomuraAún no hay calificaciones
- Músculo: Repasando Un PocoDocumento16 páginasMúsculo: Repasando Un PocoLaru MarguliesAún no hay calificaciones
- Contracción Muscular v1Documento7 páginasContracción Muscular v1Xarls StratAún no hay calificaciones
- Fases y Tipos de La Contracción MuscularDocumento6 páginasFases y Tipos de La Contracción MuscularYuval RojasAún no hay calificaciones
- 878-Texto Del Artículo-1916-1-10-20161111Documento12 páginas878-Texto Del Artículo-1916-1-10-20161111beluAún no hay calificaciones
- 05 - GlucidosDocumento61 páginas05 - GlucidosbeluAún no hay calificaciones
- 878-Texto Del Artículo-1916-1-10-20161111Documento12 páginas878-Texto Del Artículo-1916-1-10-20161111beluAún no hay calificaciones
- Linares CasasDocumento6 páginasLinares CasasCarlos CastroAún no hay calificaciones
- 878-Texto Del Artículo-1916-1-10-20161111Documento12 páginas878-Texto Del Artículo-1916-1-10-20161111beluAún no hay calificaciones
- 27-29 - SalvaneschiDocumento3 páginas27-29 - SalvaneschibeluAún no hay calificaciones
- Cirugía Robótica Transanal y Nuevas Plataformas - SACP - RevistaDocumento12 páginasCirugía Robótica Transanal y Nuevas Plataformas - SACP - RevistabeluAún no hay calificaciones
- Histo Pract 17Documento12 páginasHisto Pract 17DirectangelAún no hay calificaciones
- Programa 2019 AnatomiaDocumento23 páginasPrograma 2019 AnatomiabeluAún no hay calificaciones
- Técnicas para La Comunicación Clase 3 SumilladoDocumento22 páginasTécnicas para La Comunicación Clase 3 SumilladoJosé Antonio Rojas CalderónAún no hay calificaciones
- Programa Anual SSOMADocumento3 páginasPrograma Anual SSOMAZharytha Salinas AraucoAún no hay calificaciones
- Técnicas de Control EmocionalDocumento7 páginasTécnicas de Control EmocionalCarlos CondeAún no hay calificaciones
- C3. Rueda Cañas Jehiner AndresDocumento7 páginasC3. Rueda Cañas Jehiner AndresJavier BuenoAún no hay calificaciones
- Dilemas tecnoéticos y bioéticosDocumento4 páginasDilemas tecnoéticos y bioéticosHECTOR ARMANDO RAMIREZ ESTRADA100% (1)
- Copia de Uni1 - Act1 - Tal - Sob - Aná - Uni - 1 222Documento9 páginasCopia de Uni1 - Act1 - Tal - Sob - Aná - Uni - 1 222Nancy Lucia REYES CALDERONAún no hay calificaciones
- Cuerpos ExtrañosDocumento9 páginasCuerpos ExtrañosJonathan Martinez MaldonadoAún no hay calificaciones
- Documentos Darwin DíazDocumento13 páginasDocumentos Darwin Díazcamilo muñetonAún no hay calificaciones
- P26-EPP Proc. para Uso de EPPDocumento10 páginasP26-EPP Proc. para Uso de EPPSysse PortugalAún no hay calificaciones
- Modulo 3Documento33 páginasModulo 3Edith GonzalezAún no hay calificaciones
- El Agua y EmprendimientoDocumento3 páginasEl Agua y EmprendimientoYael PinedaAún no hay calificaciones
- Solicitud de BiopsiaDocumento2 páginasSolicitud de BiopsiaKimberly Moquete MercadoAún no hay calificaciones
- Copia de Andrés Lozano - Capitulo 11Documento12 páginasCopia de Andrés Lozano - Capitulo 11Fco Miguel CAún no hay calificaciones
- Tesis FRECUENCIA DE ÚLCERAS CORNEALES EN CABALLOS DE PASODocumento22 páginasTesis FRECUENCIA DE ÚLCERAS CORNEALES EN CABALLOS DE PASOFrancisco JulianAún no hay calificaciones
- 2do Grado Conocimiento Del Medio (2019-2020)Documento30 páginas2do Grado Conocimiento Del Medio (2019-2020)Ana AntonioAún no hay calificaciones
- El Sistema de Control GerencialDocumento43 páginasEl Sistema de Control GerencialHOSPITAL BELEN LAMBAYEQUE100% (1)
- Evaluación 1ADocumento3 páginasEvaluación 1AAna Bárbara Alemán JiménezAún no hay calificaciones
- Bioseguridad Cosmopolitan Peluqueria2020Documento38 páginasBioseguridad Cosmopolitan Peluqueria2020Dario ChachaAún no hay calificaciones
- Marco Legal Latinoamérica Sobre El Sistema Inocuidad deDocumento59 páginasMarco Legal Latinoamérica Sobre El Sistema Inocuidad deMaryanod DuqueAún no hay calificaciones
- Debilidad Adquirida en UciDocumento8 páginasDebilidad Adquirida en UciMaría Camila Arias CastroAún no hay calificaciones
- Manual SITAM 1.2. Cancer de Cuello UterinoDocumento41 páginasManual SITAM 1.2. Cancer de Cuello UterinoFlorencia IbarraAún no hay calificaciones
- Estudio de prefactibilidad para planta productora de salsa de palta envasadaDocumento168 páginasEstudio de prefactibilidad para planta productora de salsa de palta envasadaMarc Munaylla AltamiranoAún no hay calificaciones
- Matriz - Unidad 2 - Fase 3 - Analisis ColaborativoDocumento22 páginasMatriz - Unidad 2 - Fase 3 - Analisis ColaborativoDavid GoyesAún no hay calificaciones
- IncapacidadDocumento3 páginasIncapacidadKarlos MartínezAún no hay calificaciones
- 113 - Desnutrición Aguda, Moderada y Severa en Menores de 5 Años - 2020Documento3 páginas113 - Desnutrición Aguda, Moderada y Severa en Menores de 5 Años - 2020Isaias Orozco OspinoAún no hay calificaciones