También podría gustarte
- Informe Exposición Grupo N2 BioinformáticaDocumento4 páginasInforme Exposición Grupo N2 BioinformáticaNADIA NICOLE CELI BOHORQUEZAún no hay calificaciones
- RAPDDocumento10 páginasRAPDAlejandro MuñozAún no hay calificaciones
- Clasificación y Estimasión de La Diversidad Genética Del NopalDocumento11 páginasClasificación y Estimasión de La Diversidad Genética Del NopalMiguel Ángel Cervantes GonzálezAún no hay calificaciones
- Práctica Xiii XVDocumento16 páginasPráctica Xiii XVJUBER CALEB TOVAR VARGASAún no hay calificaciones
- Cuestionario P12Documento2 páginasCuestionario P12Diana RamosAún no hay calificaciones
- Hongos Nativos Con Potencial Degradador de Tintes IndustrialesDocumento18 páginasHongos Nativos Con Potencial Degradador de Tintes IndustrialesMaria Alejandra ChAún no hay calificaciones
- Barrero MicroarraysDocumento10 páginasBarrero MicroarraysMiriam AcostaAún no hay calificaciones
- Práctica 2 - Reacción en Cadena de La Polimerasa.Documento6 páginasPráctica 2 - Reacción en Cadena de La Polimerasa.José CarlosAún no hay calificaciones
- Trabajo Final PCR y Secuenciación - TiradoZazuetaGuadalupeDocumento10 páginasTrabajo Final PCR y Secuenciación - TiradoZazuetaGuadalupeGuadalupe TZAún no hay calificaciones
- 47 Secuenciación DnaDocumento35 páginas47 Secuenciación DnaXio EstradaAún no hay calificaciones
- Informe 2 Genetica - Prueba PCRDocumento6 páginasInforme 2 Genetica - Prueba PCRCamila Cifuentes CorreaAún no hay calificaciones
- Sistemática MicrobianaDocumento3 páginasSistemática MicrobianaeugeniadonosoAún no hay calificaciones
- Informe de Actividades de Practicas Profecionales Del Grupo de Investigacion en Evolucion y Sistematica MolecularDocumento9 páginasInforme de Actividades de Practicas Profecionales Del Grupo de Investigacion en Evolucion y Sistematica MolecularEdgardo Galofre Diaz-GranadosAún no hay calificaciones
- Biotecnologia Agricola. Unidad 1 - Lectura 1 (Iquira Zabanick, Enrique)Documento7 páginasBiotecnologia Agricola. Unidad 1 - Lectura 1 (Iquira Zabanick, Enrique)Enrique Iquira ZabanickAún no hay calificaciones
- Práctica No.3 "Simulación de Extracción de ADN, PCR, Electroforesis y Microarreglos Nivel Laboratorio in Silico"Documento7 páginasPráctica No.3 "Simulación de Extracción de ADN, PCR, Electroforesis y Microarreglos Nivel Laboratorio in Silico"Stephanie ChaparroAún no hay calificaciones
- 2do Previo B.M b1Documento5 páginas2do Previo B.M b1dina luz montejp riconAún no hay calificaciones
- Práctica 5. Métodos de Identificación Genética e Inmunológica.Documento7 páginasPráctica 5. Métodos de Identificación Genética e Inmunológica.Tabata SantanaAún no hay calificaciones
- Informe 13 - BioinfomaticaDocumento9 páginasInforme 13 - Bioinfomaticajavier ramosAún no hay calificaciones
- Extraccion ModificadoDocumento11 páginasExtraccion ModificadoSULBEY KARINA FERNANDEZ FIGUEROAAún no hay calificaciones
- RFLP (Restriction Fragment Length Polymorphisms)Documento16 páginasRFLP (Restriction Fragment Length Polymorphisms)Jhon Kevin Bonilla LeónAún no hay calificaciones
- T10. Analisis de Datos de ExpresionDocumento36 páginasT10. Analisis de Datos de ExpresionSamuel Peralta OjedaAún no hay calificaciones
- Identificación Bacteriana Mediante SecuenciaciónDocumento7 páginasIdentificación Bacteriana Mediante SecuenciaciónLuis Andres Henriquez PastenAún no hay calificaciones
- Introducción A La Genetica MicrobianaDocumento4 páginasIntroducción A La Genetica MicrobianaMarcos MontielAún no hay calificaciones
- Microarrays Como Herramientas de Investigación y Dispositivos de DiagnósticoDocumento15 páginasMicroarrays Como Herramientas de Investigación y Dispositivos de DiagnósticoSyndel ConchaAún no hay calificaciones
- INFORME PCR TerminadoDocumento14 páginasINFORME PCR TerminadoDavid AnlaAún no hay calificaciones
- Informe N°6 PCR-Electroforesis 2018Documento12 páginasInforme N°6 PCR-Electroforesis 2018JoséGuillénAún no hay calificaciones
- Lectura 5Documento16 páginasLectura 5Oscar O.Aún no hay calificaciones
- Práctica de Laboratorio Sobre PCRDocumento5 páginasPráctica de Laboratorio Sobre PCRDayana Guadalupe Medina MendozaAún no hay calificaciones
- Transcriptómica PDFDocumento46 páginasTranscriptómica PDFLeonardo Paulin100% (1)
- Lab - PCRDocumento8 páginasLab - PCRNatividad OrtegaAún no hay calificaciones
- RFLP y PCRDocumento2 páginasRFLP y PCRAlexander Casian PlazaAún no hay calificaciones
- 2045-3701-2-26 EsDocumento13 páginas2045-3701-2-26 EsFloral_Ana_Gut_6430Aún no hay calificaciones
- Taller Numero 4Documento9 páginasTaller Numero 4HANS STEVEN VELASCO ARIASAún no hay calificaciones
- Informe G&e-S10Documento8 páginasInforme G&e-S10MARIA FERNANDA PINEDO AZA�EROAún no hay calificaciones
- Universidad Nacional de Tumbes Facultad de Ciencias Agrarias Escuela de Medicina Veterinaria Y ZootecniaDocumento7 páginasUniversidad Nacional de Tumbes Facultad de Ciencias Agrarias Escuela de Medicina Veterinaria Y ZootecniaMaritza ZapataAún no hay calificaciones
- PCR InformeDocumento6 páginasPCR InformeSergio PinedoAún no hay calificaciones
- CibertorioDocumento17 páginasCibertorioMayra LoorAún no hay calificaciones
- Dna Polimórfico Amplificado Al AzarDocumento4 páginasDna Polimórfico Amplificado Al AzarJosé PaterninaAún no hay calificaciones
- Análisis de La Expresión DiferencialDocumento4 páginasAnálisis de La Expresión DiferencialJose Luis Can EkAún no hay calificaciones
- Informe 7Documento7 páginasInforme 7Julieth Salcedo100% (1)
- Lab 3 MoleDocumento3 páginasLab 3 MolePaulaAún no hay calificaciones
- PCR y Deteccion de VirusDocumento41 páginasPCR y Deteccion de VirusJhon JimenezAún no hay calificaciones
- Micro ArraysDocumento6 páginasMicro ArraysKevin AnrangoAún no hay calificaciones
- A05v31n2 PDFDocumento6 páginasA05v31n2 PDFAlejandro JoséAún no hay calificaciones
- ARNr 16 S GRUPO3Documento6 páginasARNr 16 S GRUPO3Francisco OlivaresAún no hay calificaciones
- Artículo Completo PCRDocumento9 páginasArtículo Completo PCRfranciabasquetAún no hay calificaciones
- Reporte 2 PCR Eq3Documento6 páginasReporte 2 PCR Eq3Lenia Gabriela Landeros HernandezAún no hay calificaciones
- Tecnicas de Laboratorio Usadas en GeneticaDocumento12 páginasTecnicas de Laboratorio Usadas en Geneticajgtv12Aún no hay calificaciones
- PCR - Reaccion en Cadena de La Polimerasa y AluDocumento9 páginasPCR - Reaccion en Cadena de La Polimerasa y AluArantxa González IturraAún no hay calificaciones
- Nuevos Métodos de Extracción de Ácidos Ribonucleicos (ARN) : Herramientas Básicas en La Biología MolecularDocumento4 páginasNuevos Métodos de Extracción de Ácidos Ribonucleicos (ARN) : Herramientas Básicas en La Biología Molecularpaola centenoAún no hay calificaciones
- PCRDocumento43 páginasPCRSam CuocoAún no hay calificaciones
- Reaccion en Cadena de PolimerasaDocumento5 páginasReaccion en Cadena de PolimerasaMedicina Gen2018Aún no hay calificaciones
- Dogma de La Biologia MolecularDocumento66 páginasDogma de La Biologia MolecularAddy AcevedoAún no hay calificaciones
- Informe de Protocolos de Extracción de ADN e Identificación de Bacterias Por El 16S y MetogenomicaDocumento11 páginasInforme de Protocolos de Extracción de ADN e Identificación de Bacterias Por El 16S y MetogenomicaViviana Jenifer VILCA ARPAAún no hay calificaciones
- Caso 2 BCMDocumento5 páginasCaso 2 BCMCecilia CaiafaAún no hay calificaciones
- Fundamentos y técnicas básicas en biología molecularDe EverandFundamentos y técnicas básicas en biología molecularAún no hay calificaciones
- Introducción a la antropología biológica: Origen, variabilidad y adaptación de las poblaciones humanasDe EverandIntroducción a la antropología biológica: Origen, variabilidad y adaptación de las poblaciones humanasAún no hay calificaciones
- Estimulacion Electrica FuncionalDocumento3 páginasEstimulacion Electrica FuncionalCristian Fernando Montenegro SalinasAún no hay calificaciones
- Bases BioAlei - Sugerencias de Uso y Recetas PrácticasDocumento35 páginasBases BioAlei - Sugerencias de Uso y Recetas Prácticasto son círculos100% (2)
- Las Proteasa Son Enzimas Que Rompen Los Enlaces Peptídicos de Las ProteínasDocumento5 páginasLas Proteasa Son Enzimas Que Rompen Los Enlaces Peptídicos de Las ProteínasrayAún no hay calificaciones
- GENERO BACILLUSyhyyDocumento28 páginasGENERO BACILLUSyhyyJen Uculmanha100% (1)
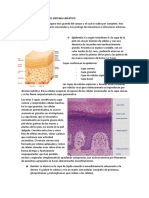
- Anatomia de La Piel y El Sistema LinfaticoDocumento2 páginasAnatomia de La Piel y El Sistema LinfaticoJosefo VlogsAún no hay calificaciones
- Fisiopatologia 2Documento17 páginasFisiopatologia 2RossAún no hay calificaciones
- Wuolah Free EXAMENESSDocumento11 páginasWuolah Free EXAMENESSAlberto Gonzalez BarrosoAún no hay calificaciones
- Estrés en Los Animales de AbastoDocumento15 páginasEstrés en Los Animales de AbastoCarmenn RvAún no hay calificaciones
- Capitulo Movimientos Elementales ComplejosDocumento13 páginasCapitulo Movimientos Elementales ComplejosOmar NavaAún no hay calificaciones
- Metabolismo Del HemoDocumento12 páginasMetabolismo Del HemoBoris 07Med-101Aún no hay calificaciones
- Un Sencillo Truco Mental - Mitch CullinDocumento889 páginasUn Sencillo Truco Mental - Mitch Cullinarenaslau100% (1)
- Manual ParasitologíaDocumento40 páginasManual ParasitologíaBastian2290Aún no hay calificaciones
- Ficha de ResumenDocumento5 páginasFicha de ResumenSolange Katherine Limaylla CoronelAún no hay calificaciones
- Información Detallada Sobre Diversos Aspectos Relacionados Con Las PalomasDocumento3 páginasInformación Detallada Sobre Diversos Aspectos Relacionados Con Las Palomas0402021032Aún no hay calificaciones
- Leyenda, Rimas, Chistes, Etc.Documento6 páginasLeyenda, Rimas, Chistes, Etc.Valentino UlinAún no hay calificaciones
- Guia Chilena de HTA Minsal 2010 PDFDocumento63 páginasGuia Chilena de HTA Minsal 2010 PDFJosefa100% (1)
- Ficha de Refuerzo: Unidad 2Documento3 páginasFicha de Refuerzo: Unidad 2Ingrid ParedesAún no hay calificaciones
- PETS # 002 (Fajas Transportadoras) OkDocumento4 páginasPETS # 002 (Fajas Transportadoras) OkgilmarAún no hay calificaciones
- 8° Guía 3 Organelos CelularesDocumento3 páginas8° Guía 3 Organelos Celularescesar villaAún no hay calificaciones
- Triptico Experimento Feria de CienciaDocumento4 páginasTriptico Experimento Feria de CienciaEner Vilchez CuevasAún no hay calificaciones
- Listado de Productos Químicos 2022Documento1 páginaListado de Productos Químicos 2022Jackelim MasiasAún no hay calificaciones
- Fisiologia CelularDocumento49 páginasFisiologia CelularPaul Cardenas BernalAún no hay calificaciones
- La Celula Cuarentena 2020 2 DoDocumento2 páginasLa Celula Cuarentena 2020 2 DoAnonymous JYHGS3JMEiAún no hay calificaciones
- La Dispersión de Las Semillas - para Chicas y Chicos de 3°Documento8 páginasLa Dispersión de Las Semillas - para Chicas y Chicos de 3°Vale GonzalezAún no hay calificaciones
- Cuadro Resumen de Los MusculosDocumento57 páginasCuadro Resumen de Los MusculosJair OrtegaAún no hay calificaciones
- Analisis Secuencia SegmentarioDocumento25 páginasAnalisis Secuencia SegmentarioMaverick 18Aún no hay calificaciones
- República Bolivariana de Venezuela Ministerio Del Poder Popular para La Educación U.E "Colegio Divino Maestro" El Tigre, Edo AnzoáteguiDocumento4 páginasRepública Bolivariana de Venezuela Ministerio Del Poder Popular para La Educación U.E "Colegio Divino Maestro" El Tigre, Edo AnzoáteguiAngelina ochoaAún no hay calificaciones
- Anamnesis Fonoaudiológica InfantilDocumento6 páginasAnamnesis Fonoaudiológica InfantilPaloma López100% (3)
- Discectomia CervicalDocumento17 páginasDiscectomia CervicalBrayan Popokitha Martinez0% (1)
- Presentacion Empresa Agricola de Papas.Documento22 páginasPresentacion Empresa Agricola de Papas.Mariana RodriguezAún no hay calificaciones