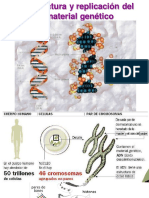
También podría gustarte
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- 2.toxicologia AmbientalDocumento33 páginas2.toxicologia AmbientalMax Jove Mayta100% (3)
- Replicación de ProcariotasDocumento36 páginasReplicación de ProcariotasCesar Rajapakse RiosAún no hay calificaciones
- Tema 2 ReplicacionDocumento26 páginasTema 2 ReplicacionLucero Gerarda Gonzalez Pizarro100% (1)
- Manual RpbiDocumento16 páginasManual RpbiisaacAún no hay calificaciones
- Replicacion, Transcripción Y Traducción.: Hugo Peña PiscoyaDocumento60 páginasReplicacion, Transcripción Y Traducción.: Hugo Peña PiscoyaCarlosAún no hay calificaciones
- Procedimiento para Identifiacion de Peligros Evaluacion y Control de RiesgosDocumento10 páginasProcedimiento para Identifiacion de Peligros Evaluacion y Control de RiesgosGermánAún no hay calificaciones
- ReplicaciónDocumento6 páginasReplicaciónLucia HernandezAún no hay calificaciones
- REPLICACIÓN DEL ADN SEMICONSERVADORADocumento25 páginasREPLICACIÓN DEL ADN SEMICONSERVADORAElsa MoranAún no hay calificaciones
- Tema 2 ReplicacionDocumento25 páginasTema 2 ReplicacionArsenio Flores VasquezAún no hay calificaciones
- Etapas de La ReplicaciónDocumento34 páginasEtapas de La ReplicaciónChris Guerrero100% (1)
- Biosintesis en Adn y ReparaciónDocumento11 páginasBiosintesis en Adn y ReparaciónPaul CapellanAún no hay calificaciones
- Foro Molecular Replicación MolecularDocumento7 páginasForo Molecular Replicación MolecularLeonardo Quintero MárquezAún no hay calificaciones
- 09 - Sintesis Del DNA. Reparacià NDocumento30 páginas09 - Sintesis Del DNA. Reparacià NAbraham HuamanAún no hay calificaciones
- REPLICACIONDocumento21 páginasREPLICACIONElias CortezAún no hay calificaciones
- Replicación: Síntesis de ADN: Biología Molecular y GenéticaDocumento34 páginasReplicación: Síntesis de ADN: Biología Molecular y GenéticaJosé CisternasAún no hay calificaciones
- Replicación en Bacterias 281119Documento11 páginasReplicación en Bacterias 281119Isabel GasparAún no hay calificaciones
- C3-Metabolismo Del ADNDocumento76 páginasC3-Metabolismo Del ADNJAIME ALEXANDER PEÑATE GONZALEZAún no hay calificaciones
- Replicación del ADN: proceso semiconservativo y bidireccionalDocumento12 páginasReplicación del ADN: proceso semiconservativo y bidireccionalUrielGranadosAún no hay calificaciones
- 3 Replicación de ADN. Grupo2Documento27 páginas3 Replicación de ADN. Grupo2July Aguila100% (1)
- ReplicaciónDocumento29 páginasReplicaciónPaula OrtegaAún no hay calificaciones
- Adn Inf GeneticaDocumento7 páginasAdn Inf Geneticaaitana pucheAún no hay calificaciones
- Replicacion y Reparacion Del DNADocumento25 páginasReplicacion y Reparacion Del DNAYúlian Loyola FloresAún no hay calificaciones
- REPLICACIÓNDocumento8 páginasREPLICACIÓNRuben Nuñez QuesadaAún no hay calificaciones
- Fragmento de OkasakiDocumento8 páginasFragmento de OkasakiYaritza Anzaldo PérezAún no hay calificaciones
- ReplicacionDocumento28 páginasReplicacionAurelia Gonzales CalleAún no hay calificaciones
- ReplicaciónDocumento6 páginasReplicaciónJesúsAún no hay calificaciones
- Ciclo Del CloroDocumento13 páginasCiclo Del Clorojoel freireAún no hay calificaciones
- ReplicaciónDocumento24 páginasReplicaciónCristhian OlivarAún no hay calificaciones
- Bmi TDocumento176 páginasBmi TJuan Arcos HuasasquicheAún no hay calificaciones
- 3 Replicación y Mecanismos de Reparación Del ADN (2021 PS) - Clase 3Documento173 páginas3 Replicación y Mecanismos de Reparación Del ADN (2021 PS) - Clase 3KATALINA IGNACIA AZÓCARAún no hay calificaciones
- 2 Replicacion-PCR-propiedades Bioquimicas ADN v1 PDFDocumento64 páginas2 Replicacion-PCR-propiedades Bioquimicas ADN v1 PDFIrene CarballoAún no hay calificaciones
- Dia Replicacion Del AdnDocumento13 páginasDia Replicacion Del Adnjoel freireAún no hay calificaciones
- Actividades genéticaDocumento6 páginasActividades genéticainfojordi.06Aún no hay calificaciones
- Unidad 4. Replicaci-N. BM 2024ADocumento38 páginasUnidad 4. Replicaci-N. BM 2024Adecker1813Aún no hay calificaciones
- Tema 15 Genética molecularDocumento80 páginasTema 15 Genética molecularCode - NoSoyjessAún no hay calificaciones
- Replicación Del ADNDocumento13 páginasReplicación Del ADNRomina OrejuelaAún no hay calificaciones
- Módulo IVDocumento28 páginasMódulo IVJuan Carlos Cazarez ZaragozaAún no hay calificaciones
- Clase 4 Unidad 3 - ReplicaciónDocumento41 páginasClase 4 Unidad 3 - ReplicaciónLeilaAún no hay calificaciones
- UntitledDocumento17 páginasUntitledNatalia LawwlietAún no hay calificaciones
- Las DNADocumento8 páginasLas DNAClara SerranoAún no hay calificaciones
- TEMA5Documento32 páginasTEMA5Claribel ValdezAún no hay calificaciones
- Replicación del ADN: Procesos, Diferencias y Enzimas InvolucradasDocumento6 páginasReplicación del ADN: Procesos, Diferencias y Enzimas InvolucradasPedro CandelarioAún no hay calificaciones
- 12 - Genética Molecular IDocumento17 páginas12 - Genética Molecular IMiguel SanchezAún no hay calificaciones
- Bio. (32) Dogma de La Biologia Molecular 57 - 68Documento12 páginasBio. (32) Dogma de La Biologia Molecular 57 - 68ALEXANDER DAVID LOPEZ ESCOBEDOAún no hay calificaciones
- Semana 5 - Sesión 2 - BiologíaDocumento15 páginasSemana 5 - Sesión 2 - Biologíasolanonataly362Aún no hay calificaciones
- 5 - Replicación Del DNA DMCT IIDocumento45 páginas5 - Replicación Del DNA DMCT IIsvrt0408Aún no hay calificaciones
- 2do PARCIALDocumento76 páginas2do PARCIALGiuliana EhlertAún no hay calificaciones
- Replicacion en Procariotas y EucariotasDocumento5 páginasReplicacion en Procariotas y EucariotasJose Santos Gonzales HernandesAún no hay calificaciones
- ReplicaciónDocumento34 páginasReplicaciónPAZ MARTINA ALLENDESAún no hay calificaciones
- Replicación del ADN: Mecanismos moleculares y diferencias entre bacterias y eucariotasDocumento51 páginasReplicación del ADN: Mecanismos moleculares y diferencias entre bacterias y eucariotasNathalie Rodriguez0% (1)
- REPLICACION PresentacionDocumento32 páginasREPLICACION PresentacionMaría Nathalie Hidálgo González BAún no hay calificaciones
- El Adn Mutaciones E Ingeniería GenéticaDocumento49 páginasEl Adn Mutaciones E Ingeniería GenéticaHjb HujjAún no hay calificaciones
- Replicaciondeladn 180416143831Documento22 páginasReplicaciondeladn 180416143831EVELIN NAOMI PEREZ MATEOAún no hay calificaciones
- Tema-5 3 - ReplicaciónDocumento59 páginasTema-5 3 - ReplicaciónElizabeth JiménezAún no hay calificaciones
- Tema 5 - Biología MolecularDocumento8 páginasTema 5 - Biología MolecularMari Carmen ReinéAún no hay calificaciones
- Metabolismo Del AdnDocumento59 páginasMetabolismo Del AdnM Orihuela100% (1)
- Impri MirDocumento16 páginasImpri MirDaniel HenryAún no hay calificaciones
- Bio Con ComentariosDocumento7 páginasBio Con Comentariosnamjoon940605Aún no hay calificaciones
- Semana13 Teoria replicacionADNDocumento18 páginasSemana13 Teoria replicacionADNwh0 4r3 y70 uwuAún no hay calificaciones
- Replicacion ADNDocumento39 páginasReplicacion ADNVidal RTAún no hay calificaciones
- Ensayo de ReplicaciónDocumento4 páginasEnsayo de ReplicaciónFernanda MorenoAún no hay calificaciones
- La Replicación Del ADNDocumento2 páginasLa Replicación Del ADNDavid Nieves CamañoAún no hay calificaciones
- Manual de Toma de MuestrasDocumento23 páginasManual de Toma de MuestrasisaacAún no hay calificaciones
- Expo BancoDocumento8 páginasExpo BancoisaacAún no hay calificaciones
- Reporte Práctica 7Documento7 páginasReporte Práctica 7isaacAún no hay calificaciones
- Estadística DescriptivaDocumento24 páginasEstadística Descriptivaisaac0% (1)
- Sustantivo FRDocumento5 páginasSustantivo FRAngelica ChoquecotaAún no hay calificaciones
- Microscopio: Una ventana al mundo microscópicoDocumento3 páginasMicroscopio: Una ventana al mundo microscópicoEdson Noel Flórez AlvaradoAún no hay calificaciones
- Prueba Diagnostica 1Documento4 páginasPrueba Diagnostica 1Constanza GuerreroAún no hay calificaciones
- Diapositivas de Psicologia EvolutivaDocumento15 páginasDiapositivas de Psicologia EvolutivaVanesa JuradoAún no hay calificaciones
- Redes tróficas y cadenas alimentariasDocumento8 páginasRedes tróficas y cadenas alimentariaselimarAún no hay calificaciones
- Aplicacion de Un Programa de EstimulaciónDocumento81 páginasAplicacion de Un Programa de Estimulacióngenesis estheiryAún no hay calificaciones
- QUIMIOTAXISDocumento8 páginasQUIMIOTAXISMaría D MoraLesAún no hay calificaciones
- Horario VETDocumento8 páginasHorario VETAlejandro Albarrán ÁlvarezAún no hay calificaciones
- Tema 10 Orgc3a1nulos Energc3a9ticos - PpsDocumento13 páginasTema 10 Orgc3a1nulos Energc3a9ticos - PpsLeidy CastroAún no hay calificaciones
- Solucion 1 Taller Central de EsterilizcionDocumento2 páginasSolucion 1 Taller Central de EsterilizcionLady Diana MarinAún no hay calificaciones
- Glosario 3Documento40 páginasGlosario 3Bryan Bustamante GuerraAún no hay calificaciones
- La Teoría de La Evolución, Explicada A PrincipiantesDocumento3 páginasLa Teoría de La Evolución, Explicada A PrincipiantesJhonn T. RojasAún no hay calificaciones
- Bioquimica HigadoDocumento6 páginasBioquimica HigadoJorge HipólitoAún no hay calificaciones
- Que Es AlelopatíaDocumento4 páginasQue Es AlelopatíaMultiservicios BatistaAún no hay calificaciones
- Recuento de leucocitos y cámara de NeubauerDocumento12 páginasRecuento de leucocitos y cámara de NeubauerDiego CortesAún no hay calificaciones
- TAREA1-FASE 2 (3) para SubrirDocumento12 páginasTAREA1-FASE 2 (3) para SubrirMAuricio LeytonAún no hay calificaciones
- Acción Inmunológica, Trasplantes y Sangre SolDocumento24 páginasAcción Inmunológica, Trasplantes y Sangre SolClaudia CáceresAún no hay calificaciones
- Triptico de FruticulturaDocumento2 páginasTriptico de FruticulturaKatheriine SteephanyAún no hay calificaciones
- Informe de Laboratorio 5-T Pinargote PDFDocumento7 páginasInforme de Laboratorio 5-T Pinargote PDFtania pinargoteAún no hay calificaciones
- Gonadas y Gametos #3Documento16 páginasGonadas y Gametos #3javier elias salcedo ramosAún no hay calificaciones
- Ciminalística ProtozoosDocumento74 páginasCiminalística ProtozoosLorena SanabriaAún no hay calificaciones
- Monografía Camellia Sinensis - Mateo Piñeros Rhon y Mara D. KujundzicDocumento11 páginasMonografía Camellia Sinensis - Mateo Piñeros Rhon y Mara D. KujundzicMateo Piñeros RhonAún no hay calificaciones
- FisiologíaDocumento9 páginasFisiologíaGerson LópezAún no hay calificaciones
- Tema 3 Clasificacion de TejidosDocumento59 páginasTema 3 Clasificacion de TejidosMauricio GuillenAún no hay calificaciones
- Metilación y cáncer: una nueva vía de tratamientoDocumento3 páginasMetilación y cáncer: una nueva vía de tratamientoBryan LámbarryAún no hay calificaciones
- Secrecion Interna Externa y MixtasDocumento2 páginasSecrecion Interna Externa y MixtasWilliam JimenezAún no hay calificaciones
- Penetracion Del Patogeno en La PlantaDocumento14 páginasPenetracion Del Patogeno en La PlantaEhidy Thania Barahona CardenasAún no hay calificaciones
- Tratamientos EnfermedadesDocumento1 páginaTratamientos EnfermedadesBrayanRojasAún no hay calificaciones