También podría gustarte
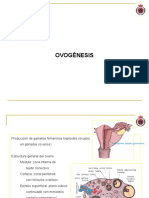
- 03 Gónadas Femeninas - Origen Embriológico - Ovogénesis - Ciclos EstralesDocumento23 páginas03 Gónadas Femeninas - Origen Embriológico - Ovogénesis - Ciclos EstralesLucía María BradleyAún no hay calificaciones
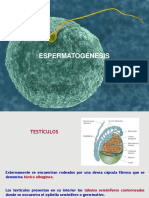
- Procesos reproductivos masculinos: Espermatogénesis y formación de espermatozoidesDocumento53 páginasProcesos reproductivos masculinos: Espermatogénesis y formación de espermatozoidesAmanda Hernández CuadraAún no hay calificaciones
- Anemia Para Todos: Medicina Para TodosDe EverandAnemia Para Todos: Medicina Para TodosCalificación: 3 de 5 estrellas3/5 (1)
- Primeras SemanasDocumento7 páginasPrimeras SemanasJoel Zabala zuritaAún no hay calificaciones
- MF0712_1 - Operaciones auxiliares en reproducción ganaderaDe EverandMF0712_1 - Operaciones auxiliares en reproducción ganaderaAún no hay calificaciones
- Aparato Reproductor Femeninoo pptDocumento41 páginasAparato Reproductor Femeninoo pptCecilia De la Cruz de Santa CruzAún no hay calificaciones
- GametogénesisDocumento7 páginasGametogénesis2doKAún no hay calificaciones
- Histología Sistemas ReproductoresDocumento20 páginasHistología Sistemas ReproductoresAle tcAún no hay calificaciones
- Embrio SegundasDocumento35 páginasEmbrio SegundasKaryme GisAún no hay calificaciones
- Establecimiento de La GestacionDocumento7 páginasEstablecimiento de La GestacionmarizhgAún no hay calificaciones
- Embriología: Desarrollo Ovulo Espermatozoide FecundaciónDocumento35 páginasEmbriología: Desarrollo Ovulo Espermatozoide FecundaciónLissetteAlexandraPerezAún no hay calificaciones
- Lección No 1. Fisiología de La Reproduccion1Documento29 páginasLección No 1. Fisiología de La Reproduccion1vimarjuAún no hay calificaciones
- Capitulo 23 Sistema Genital FemeninoDocumento16 páginasCapitulo 23 Sistema Genital FemeninoJohan ChoqueAún no hay calificaciones
- Proceso GametogénesisDocumento4 páginasProceso GametogénesisAlexander EztradaAún no hay calificaciones
- Embrioapuntes (Parte I) - Diego MeléndezDocumento16 páginasEmbrioapuntes (Parte I) - Diego Meléndezdiego melendezAún no hay calificaciones
- EmbriologíaDocumento17 páginasEmbriologíaCarmen Porras Jarufe50% (2)
- Gametogénesis y embriología del aparato genital femeninoDocumento12 páginasGametogénesis y embriología del aparato genital femeninoAlma Linda Peréz Santeliz100% (1)
- El Pibe Resumen de FecundaciónDocumento29 páginasEl Pibe Resumen de FecundaciónAntonella Violo100% (1)
- Loe11 FisiologiaDocumento30 páginasLoe11 FisiologiaJuan Pablo Giraldo MontoyaAún no hay calificaciones
- Aparato Reproductor HembraDocumento46 páginasAparato Reproductor HembraGonzalo MunAún no hay calificaciones
- Resumen Embriologia LangmanDocumento13 páginasResumen Embriologia Langmanmanuel_aguirre_13Aún no hay calificaciones
- Gametogénesis y ciclo sexual.docxDocumento4 páginasGametogénesis y ciclo sexual.docxValentina BOLDRINIAún no hay calificaciones
- Embriologia BasicaDocumento22 páginasEmbriologia BasicaJose AvilaAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento17 páginasAparato Reproductor FemeninoJuan Esteban GarciaAún no hay calificaciones
- Fisiologia Del Eje H-H-ODocumento18 páginasFisiologia Del Eje H-H-OLuz Milagritos100% (1)
- Resumen Langman Apuntes CompletoDocumento21 páginasResumen Langman Apuntes CompletoL J0% (1)
- Sesión 03 - Citología ExfoliativaDocumento10 páginasSesión 03 - Citología Exfoliativafreddy.quisperuedaAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento20 páginasAparato Reproductor FemeninoKoa NunchiAún no hay calificaciones
- UABP 2, 6 Y 9 - EmbriologíaDocumento15 páginasUABP 2, 6 Y 9 - EmbriologíaGiovanni MedranoAún no hay calificaciones
- Semanas 1-8 EmbriologíaDocumento15 páginasSemanas 1-8 EmbriologíaAndreita GarciaAún no hay calificaciones
- Ovogénesis (WIkipediaDocumento4 páginasOvogénesis (WIkipediaFernandoAún no hay calificaciones
- Clase 2 ObstetriciaDocumento47 páginasClase 2 ObstetriciaRaúl Alexis Menudo ProblemaAún no hay calificaciones
- 1.-Reproduccion-humanaenero21Documento23 páginas1.-Reproduccion-humanaenero21vsifontes91Aún no hay calificaciones
- Primera Semana de Desarrollo EmbrionarioDocumento5 páginasPrimera Semana de Desarrollo EmbrionarioEnoc GonzálezAún no hay calificaciones
- Cuadro Comparativo Entre Ovogenesis y EspermatogenesisDocumento5 páginasCuadro Comparativo Entre Ovogenesis y Espermatogenesisgavio_ta26100% (1)
- Fisiología de La Reproducción HumanaDocumento9 páginasFisiología de La Reproducción HumanasuperchiitoAún no hay calificaciones
- Gametogenesis M1.3Documento10 páginasGametogenesis M1.3Hector Efrain Garcia ChavezAún no hay calificaciones
- Células Germinales PrimordialesDocumento11 páginasCélulas Germinales Primordialesmariferc319Aún no hay calificaciones
- Ciclo ReproductorDocumento9 páginasCiclo ReproductorMily BenincaAún no hay calificaciones
- Gametogénesis: cambios morfológicos durante la maduración de los gametosDocumento8 páginasGametogénesis: cambios morfológicos durante la maduración de los gametosFlor Denisse Villa LópezAún no hay calificaciones
- Preparación para el embarazo: Gametogénesis y maduración de óvulos y espermatozoidesDocumento3 páginasPreparación para el embarazo: Gametogénesis y maduración de óvulos y espermatozoidesJean SalgadoAún no hay calificaciones
- GAMETOGENÉSISDocumento6 páginasGAMETOGENÉSISanon_757572911Aún no hay calificaciones
- Aparato Reproductor FemeninoDocumento10 páginasAparato Reproductor FemeninoMANRIQUE GUTIÉRREZ STEPHANY GIOMAYRAAún no hay calificaciones
- Ciclo Menstrual y Sus TrastornosDocumento19 páginasCiclo Menstrual y Sus TrastornosGustavo Enrique Carrasco Perez100% (1)
- GametogenesisDocumento28 páginasGametogenesisReinaldo Chirinos ArredondoAún no hay calificaciones
- Proceso de la gametogénesis, fecundación e implantaciónDocumento80 páginasProceso de la gametogénesis, fecundación e implantaciónvalepiastre100% (1)
- Ovogénesis y FoliculogénesisDocumento36 páginasOvogénesis y FoliculogénesisSusana SotoAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento20 páginasAparato Reproductor FemeninojavierAún no hay calificaciones
- Espermatogenesis y OvogenesisDocumento13 páginasEspermatogenesis y OvogenesisElvis Humantupa Estrada100% (3)
- OvogénesisDocumento11 páginasOvogénesisMaria Claudia Castro HumpireAún no hay calificaciones
- Cuadro Comparativo Entre Ovogenesis y EspermatogenesisDocumento5 páginasCuadro Comparativo Entre Ovogenesis y Espermatogenesisgavio_ta2669% (13)
- ChloeDocumento10 páginasChloeSovietWantanAún no hay calificaciones
- Ciclo Ovárico y Ciclo MenstrualDocumento4 páginasCiclo Ovárico y Ciclo MenstrualDenisse SarabiaAún no hay calificaciones
- 37 Espermatogénesis, Ovogénesis, FecundaciónDocumento29 páginas37 Espermatogénesis, Ovogénesis, FecundaciónCarlaAún no hay calificaciones
- Ciclo Sexual y Ovarios. OvariosDocumento9 páginasCiclo Sexual y Ovarios. Ovariosmaria nochettiAún no hay calificaciones
- Reproduccion 3Documento57 páginasReproduccion 3Angela YanezAún no hay calificaciones
- Eje hipotálamo-hipófisis-ovario (HHODocumento58 páginasEje hipotálamo-hipófisis-ovario (HHOSaul Saavedra PeraltaAún no hay calificaciones
- OvonogenesisDocumento15 páginasOvonogenesisNicole SalamancaAún no hay calificaciones
- Diabetesgestacional 17040301Documento34 páginasDiabetesgestacional 17040301Danilo Jesús Mendoza MaldonadoAún no hay calificaciones
- 0EV Adulto Mayor 2013Documento13 páginas0EV Adulto Mayor 2013YOshie ParrasAún no hay calificaciones
- Asma Bronq DanDocumento21 páginasAsma Bronq DanDanilo Jesús Mendoza MaldonadoAún no hay calificaciones
- Historia Clinica PediatricaDocumento4 páginasHistoria Clinica PediatricaDanilo Jesús Mendoza MaldonadoAún no hay calificaciones
- 1 Presentacion Protocolo Esavi Campana SR 2006Documento34 páginas1 Presentacion Protocolo Esavi Campana SR 2006tibinurseAún no hay calificaciones
- 10 Clase - PartogramaDocumento78 páginas10 Clase - PartogramaDanilo Jesús Mendoza MaldonadoAún no hay calificaciones
- ArtritispsoriasicaDocumento15 páginasArtritispsoriasicaDanilo Jesús Mendoza MaldonadoAún no hay calificaciones
- Banco de Preguntas #01Documento241 páginasBanco de Preguntas #01Eduardo R Cano Luján100% (1)
- 10 Clase - PartogramaDocumento78 páginas10 Clase - PartogramaDanilo Jesús Mendoza MaldonadoAún no hay calificaciones
- PRUEBA A (1) Examen Extraordinario 2015 PDFDocumento11 páginasPRUEBA A (1) Examen Extraordinario 2015 PDFPool Cárdenas Delgado67% (3)
- Etica y DeontologiaDocumento32 páginasEtica y DeontologiaDanilo Jesús Mendoza MaldonadoAún no hay calificaciones
- Responsabilidad Medica 01.PtxDocumento7 páginasResponsabilidad Medica 01.PtxDanilo Jesús Mendoza MaldonadoAún no hay calificaciones
- Liderazgo SaludDocumento25 páginasLiderazgo SaludDanilo Jesús Mendoza MaldonadoAún no hay calificaciones
- Bases GenotipicasDocumento5 páginasBases Genotipicasjose luis de la cruz100% (1)
- Ciclo Azufre y NitrógenoDocumento4 páginasCiclo Azufre y NitrógenoBerenice S. OleaAún no hay calificaciones
- Potencial de AcciónDocumento5 páginasPotencial de AcciónJose Cisneros0% (1)
- S9 - Ficha de Actividades - 4toDocumento3 páginasS9 - Ficha de Actividades - 4toJoselyn Valeria Gòngora MarcosAún no hay calificaciones
- Impacto Social Del Uso de Losplaguicidas Químicos en El Mundo - Plaguicidas Químicos en El MundoDocumento24 páginasImpacto Social Del Uso de Losplaguicidas Químicos en El Mundo - Plaguicidas Químicos en El MundocastcarlAún no hay calificaciones
- Concepto de Ser Vivo y Metodo CientificoDocumento7 páginasConcepto de Ser Vivo y Metodo CientificopepeAún no hay calificaciones
- Sistema Crisp RDocumento4 páginasSistema Crisp RNormaAún no hay calificaciones
- Hibridación de Ácidos NucleicosDocumento34 páginasHibridación de Ácidos NucleicosDiana Camacho PlascenciaAún no hay calificaciones
- Semiologia de Sistema Neurologico 2021.Documento62 páginasSemiologia de Sistema Neurologico 2021.TERESA ESPINOZA SABAAún no hay calificaciones
- Enfermedad ParasitariaDocumento3 páginasEnfermedad ParasitariaTony Milian UnlockerAún no hay calificaciones
- Plantas Trabajo de InvestigaciónDocumento3 páginasPlantas Trabajo de InvestigaciónPriscila Ruth Avezón SaavedraAún no hay calificaciones
- Clasificación Del MetabolismoDocumento7 páginasClasificación Del MetabolismoAna Gabriela Hans SalazarAún no hay calificaciones
- Quien Quiere Ser FonoaudiologoDocumento3 páginasQuien Quiere Ser FonoaudiologoCristian MorenoAún no hay calificaciones
- Hibridación de PecesDocumento7 páginasHibridación de Pecesbladimir zamora100% (1)
- 5 Reinos PDFDocumento7 páginas5 Reinos PDFJeffrey WangAún no hay calificaciones
- Naturales 9 Guia 3Documento13 páginasNaturales 9 Guia 3danna cabrera rodriguezAún no hay calificaciones
- Diapositivas BacteriasDocumento26 páginasDiapositivas BacteriasGloria ReyesAún no hay calificaciones
- Tejido GlandularDocumento47 páginasTejido Glandularmywnrj8r78Aún no hay calificaciones
- Los hongos: características, clasificación y reproducciónDocumento29 páginasLos hongos: características, clasificación y reproducciónJOSUE FRANCISCO MOJICA LEON100% (1)
- Métodos Rápidos y Automatización en Microbiología AlimentariaDocumento13 páginasMétodos Rápidos y Automatización en Microbiología AlimentariaCarlos jesus hernandez mezaAún no hay calificaciones
- Botulismo FusionadoDocumento135 páginasBotulismo FusionadoKelly Puerta DavisAún no hay calificaciones
- Fitopatología Primera Evaluación Práctica 2020-I (Reparado)Documento6 páginasFitopatología Primera Evaluación Práctica 2020-I (Reparado)Valentina Pulido HuertasAún no hay calificaciones
- Resumen NutricionPlantasDocumento4 páginasResumen NutricionPlantasMarino Mendoza Ruiz100% (1)
- Syllabus Sist Neuroendocrino Cardiopulmonar y Renal 2019 - 2 (3941)Documento40 páginasSyllabus Sist Neuroendocrino Cardiopulmonar y Renal 2019 - 2 (3941)Yoiner BustosAún no hay calificaciones
- Estructura de La Célula ProcariotaDocumento4 páginasEstructura de La Célula ProcariotaCompu RhemaAún no hay calificaciones
- Organelos bacterianos: Cápsula, biofilms y esporasDocumento20 páginasOrganelos bacterianos: Cápsula, biofilms y esporasTeffaLoAún no hay calificaciones
- CervicovaginitisDocumento39 páginasCervicovaginitisXóchitl Velázquez Lara100% (1)
- Celula Euca y Proca 4°a JBDocumento27 páginasCelula Euca y Proca 4°a JBKenny GuerreroAún no hay calificaciones
- Ciclo de La UreaDocumento4 páginasCiclo de La UreaLuu MoralezAún no hay calificaciones
- Tema 2 Estructura Del AmbienteDocumento28 páginasTema 2 Estructura Del AmbienteBlacmont Josset AndreaAún no hay calificaciones