También podría gustarte
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- TranscripciónDocumento28 páginasTranscripciónAurelia Gonzales Calle100% (2)
- Teoría Replicación y Transcripción Del ADN, Traducción Del ARN.Documento37 páginasTeoría Replicación y Transcripción Del ADN, Traducción Del ARN.Juan Torres GalindoAún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Patología Ano-RectalDocumento73 páginasPatología Ano-Rectal4317322Aún no hay calificaciones
- Dogma de La GeneticaDocumento10 páginasDogma de La GeneticaUllquiorra100% (1)
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
- Transcripcion y TraduccionDocumento22 páginasTranscripcion y TraduccionChristian Correa VerdeAún no hay calificaciones
- 4 Mod. PostranscripcionalesDocumento25 páginas4 Mod. PostranscripcionalesMelissa AlonsoAún no hay calificaciones
- TranscripciónDocumento17 páginasTranscripciónroquehernandezmonseAún no hay calificaciones
- 8 TrabajoDocumento12 páginas8 TrabajoBarrera RonnyAún no hay calificaciones
- Traducción GenéticaDocumento27 páginasTraducción GenéticaIsabel NogueiraAún no hay calificaciones
- Sintesis de ProteínasDocumento4 páginasSintesis de ProteínasKevin Andres Z.Aún no hay calificaciones
- CAPITULO 19 Splicing - En.esDocumento108 páginasCAPITULO 19 Splicing - En.esGenesis MaldonadoAún no hay calificaciones
- GENETICADocumento9 páginasGENETICAaimenAún no hay calificaciones
- Procesamiento de Arn y Su TraduccionDocumento6 páginasProcesamiento de Arn y Su TraduccionmiliferpaolaAún no hay calificaciones
- Síntesis de Proteínas - Transcripción y TraducciónDocumento3 páginasSíntesis de Proteínas - Transcripción y TraducciónEmma OchovaAún no hay calificaciones
- ArnDocumento3 páginasArnDaniel Garcia100% (1)
- Traducción en ProcariotasDocumento41 páginasTraducción en ProcariotasFreyyyyBmAún no hay calificaciones
- Taller #1 BiologiaDocumento7 páginasTaller #1 BiologiaDavid Santiago SarchiAún no hay calificaciones
- Cushicondor Alexis PAE8 AI 19 07 2022Documento3 páginasCushicondor Alexis PAE8 AI 19 07 2022Alexis CushicondorAún no hay calificaciones
- Transcripcion Del Adn - ExposicionDocumento31 páginasTranscripcion Del Adn - ExposicionJuly Lalangui Rivera100% (4)
- Resumen Síntesis de Novo Purinas y PirimidinasDocumento14 páginasResumen Síntesis de Novo Purinas y Pirimidinasconi22Aún no hay calificaciones
- 16 y 17. - Biosintesis de Proteinas - 2022Documento57 páginas16 y 17. - Biosintesis de Proteinas - 2022Keiry Stefany Hernandez LemusAún no hay calificaciones
- Obj. 2 - TranscripciónDocumento5 páginasObj. 2 - TranscripciónCristinaAún no hay calificaciones
- TRANSCRIPCIÓNDocumento21 páginasTRANSCRIPCIÓNDaniel CarpioAún no hay calificaciones
- TranscripcionDocumento4 páginasTranscripcionmafermoralesv1Aún no hay calificaciones
- Temario Tercer Parcial de BiomolDocumento7 páginasTemario Tercer Parcial de BiomolMARCO ANTONIO SOSA AGUILARAún no hay calificaciones
- Síntesis y Maduración Del RnaDocumento7 páginasSíntesis y Maduración Del RnaJessicaChumbesAún no hay calificaciones
- Guía Molecular. 3er ParcialDocumento18 páginasGuía Molecular. 3er ParcialMichelle RodríguezAún no hay calificaciones
- Transcripción y Traducción - GalánDocumento22 páginasTranscripción y Traducción - GalánJimena De Los AngelesAún no hay calificaciones
- Repaso de Biología MolecularDocumento39 páginasRepaso de Biología MolecularhielitousAún no hay calificaciones
- Parcial 4 - Bioquímica IIDocumento92 páginasParcial 4 - Bioquímica IIFanni Stefany Salazar PenadoAún no hay calificaciones
- Transcripcion y Traduccion en Celulas EucariotasDocumento5 páginasTranscripcion y Traduccion en Celulas EucariotasGenesis MacíasAún no hay calificaciones
- Biomol 2Documento5 páginasBiomol 2JOSE ARTURO LOZANO CRUZAún no hay calificaciones
- Tema 6 - Síntesis de Proteínas, TraducciónDocumento7 páginasTema 6 - Síntesis de Proteínas, TraducciónepepAún no hay calificaciones
- Maduracion RnaDocumento78 páginasMaduracion RnaPaula EchavarriaAún no hay calificaciones
- Transcripción Eucariotas y ProcariotasDocumento9 páginasTranscripción Eucariotas y ProcariotasAndrésAún no hay calificaciones
- Temario Biomol 3er ExmDocumento6 páginasTemario Biomol 3er ExmMARCO ANTONIO SOSA AGUILARAún no hay calificaciones
- TraducciónDocumento14 páginasTraducciónOSSIEL ALFREDO GARCIA ALVAREZAún no hay calificaciones
- Procesamiento Postranscripcional Del ArnDocumento26 páginasProcesamiento Postranscripcional Del ArnJorge Carlos Vazquez Sanchez100% (1)
- ARN y Sus CaracterísticasDocumento41 páginasARN y Sus CaracterísticasNazareth EspinozaAún no hay calificaciones
- Cuestionario 1. ¿Por Qué Se Ha Hecho Diferente Número de Cartas para Los Aminoácidos? ¿Existe Alguna Sustentación Biológica?Documento6 páginasCuestionario 1. ¿Por Qué Se Ha Hecho Diferente Número de Cartas para Los Aminoácidos? ¿Existe Alguna Sustentación Biológica?Gabriel MoralesAún no hay calificaciones
- 5 Clase de Mec de Traduccion LJ Ribosomas 2Documento17 páginas5 Clase de Mec de Traduccion LJ Ribosomas 2Mary BuzoAún no hay calificaciones
- Genética Mendeliana y ADNDocumento4 páginasGenética Mendeliana y ADNmireiaAún no hay calificaciones
- Traduccion Del Arn Procariota y EucariotaDocumento40 páginasTraduccion Del Arn Procariota y EucariotaUlises Infante Huaytalla100% (2)
- Resumen Segundo Parcial Biologia CelularDocumento7 páginasResumen Segundo Parcial Biologia CelularmoraAún no hay calificaciones
- Traducción Adn-ArnDocumento4 páginasTraducción Adn-ArnCindy FranceschetAún no hay calificaciones
- Sintesis de Proteinas y Acidos NucleicosDocumento5 páginasSintesis de Proteinas y Acidos NucleicosShannon HallAún no hay calificaciones
- Tema 6 - Sกntesis de proteกnas, traducciขnDocumento8 páginasTema 6 - Sกntesis de proteกnas, traducciขnJON LEGUINAAún no hay calificaciones
- Replicación, Transcripción y TraducciónDocumento4 páginasReplicación, Transcripción y TraducciónMarily Cabada50% (2)
- Ribosomal FrameshiftingDocumento37 páginasRibosomal FrameshiftingErika CoronadoAún no hay calificaciones
- Clase 9. Transcripción ARNDocumento52 páginasClase 9. Transcripción ARNChristian Robalino TorresAún no hay calificaciones
- BM11Documento46 páginasBM11Emerson Ynca PalominoAún no hay calificaciones
- Repaso Parcial de MolecularDocumento3 páginasRepaso Parcial de Molecularlaura QuinteroAún no hay calificaciones
- Transcripción GenéticaDocumento4 páginasTranscripción GenéticaAntonio TreminiAún no hay calificaciones
- Transcripcion Del AdnDocumento38 páginasTranscripcion Del AdnElenaBarbosaAún no hay calificaciones
- Bioquímica ResumenDocumento13 páginasBioquímica ResumenAlejandra LAún no hay calificaciones
- Taller TraduccionDocumento25 páginasTaller TraduccionJunior SuescunAún no hay calificaciones
- Traducción EucariotaDocumento9 páginasTraducción Eucariotadaniel rodriguez riosAún no hay calificaciones
- Tema 11Documento8 páginasTema 11boakye5353Aún no hay calificaciones
- ENARM Litiasis UrinariaDocumento67 páginasENARM Litiasis Urinaria4317322Aún no hay calificaciones
- Enarm TiroidesDocumento58 páginasEnarm Tiroides4317322100% (1)
- ENARM DiarreaDocumento13 páginasENARM Diarrea4317322Aún no hay calificaciones
- Urgencias Endocr EnarmDocumento25 páginasUrgencias Endocr Enarm4317322Aún no hay calificaciones
- Enarm FeocromocitomaDocumento7 páginasEnarm Feocromocitoma4317322Aún no hay calificaciones
- Enarm EapDocumento14 páginasEnarm Eap4317322Aún no hay calificaciones
- Enarm IamDocumento31 páginasEnarm Iam4317322Aún no hay calificaciones
- La PielDocumento111 páginasLa Piel4317322Aún no hay calificaciones
- ENARM Cardiopatia IsquemicaDocumento57 páginasENARM Cardiopatia Isquemica4317322Aún no hay calificaciones
- Intervenciones Obstetricas 1Documento38 páginasIntervenciones Obstetricas 14317322Aún no hay calificaciones
- Antihelminticos 2222222Documento32 páginasAntihelminticos 22222224317322Aún no hay calificaciones
- Mal Del PintoDocumento24 páginasMal Del Pinto43173220% (1)
- EMN Otorrino 2008Documento50 páginasEMN Otorrino 20084317322Aún no hay calificaciones
- Molecular ParcialDocumento14 páginasMolecular ParcialANDREA VALENTINA MAYORGA LOPEZAún no hay calificaciones
- 5to Informe PCRDocumento8 páginas5to Informe PCRKaren Esteves ZúñigaAún no hay calificaciones
- Fisiologia y Morfologia Del Núcleo CelularDocumento13 páginasFisiologia y Morfologia Del Núcleo CelularAbelardo Ortiz50% (4)
- Marcadas Por La Experiencia: Epigenética y Adaptación en PlantasDocumento2 páginasMarcadas Por La Experiencia: Epigenética y Adaptación en PlantasAnonymous vNgs3sAún no hay calificaciones
- Solucion Taller MitosisDocumento9 páginasSolucion Taller MitosisSergio Alejandro Castro100% (1)
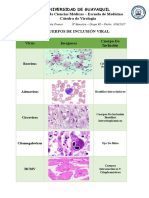
- Universidad de Guayaquil Facultad de Ciencias Médicas - Escuela de Medicina Cátedra de VirologíaDocumento4 páginasUniversidad de Guayaquil Facultad de Ciencias Médicas - Escuela de Medicina Cátedra de VirologíaGenesis GarciaAún no hay calificaciones
- Farmacia Plan 1987Documento10 páginasFarmacia Plan 1987Karen Magalí CruzAún no hay calificaciones
- Herencia MultifactorialDocumento35 páginasHerencia MultifactorialDaniel Sebastian0% (1)
- Organigrama de Enfermeria (Centro de Salud)Documento2 páginasOrganigrama de Enfermeria (Centro de Salud)So FiAún no hay calificaciones
- Celula TrabajoDocumento15 páginasCelula TrabajoMilagro MataAún no hay calificaciones
- Cuaderno Biologia y Microbologia Aplicada Cenco 3Documento5 páginasCuaderno Biologia y Microbologia Aplicada Cenco 3mauricio mallaraucoAún no hay calificaciones
- BACTERIASDocumento62 páginasBACTERIASPaola ChimbolemaAún no hay calificaciones
- BioprocesosDocumento13 páginasBioprocesosMariaAún no hay calificaciones
- Ensayo BiologiaDocumento4 páginasEnsayo BiologiaLida Luz BeltranAún no hay calificaciones
- Métodos TradicionalesDocumento37 páginasMétodos TradicionalesJurgen LunaAún no hay calificaciones
- FotosíntesisDocumento3 páginasFotosíntesisValentina MoraAún no hay calificaciones
- Danagene Spin Genomic Dna Kit PDFDocumento6 páginasDanagene Spin Genomic Dna Kit PDFCrist VillarAún no hay calificaciones
- MANUAL DE PRÁCTICAS BIOLOGIA Y QUIMICA Actualizadas....Documento83 páginasMANUAL DE PRÁCTICAS BIOLOGIA Y QUIMICA Actualizadas....thania zambranoAún no hay calificaciones
- Terapia AntibacterianaDocumento31 páginasTerapia AntibacterianaJhon Lauren's0% (1)
- Vigor HibridoDocumento18 páginasVigor Hibridofresh_2000Aún no hay calificaciones
- El VIH Un Virus Como Ningun OtroDocumento35 páginasEl VIH Un Virus Como Ningun OtroMAURICE022Aún no hay calificaciones
- Universidad Nacional Del Altiplano Puno GeneticaDocumento10 páginasUniversidad Nacional Del Altiplano Puno GeneticaNelson CariAún no hay calificaciones
- Presentation VIRUSDocumento24 páginasPresentation VIRUSIrene Catalá TrujilloAún no hay calificaciones
- 2.3 Factores de VirulenciaDocumento11 páginas2.3 Factores de VirulenciaOdontologia UnamAún no hay calificaciones
- La OsmosisDocumento6 páginasLa OsmosisJuan Felipe Villa DiegoAún no hay calificaciones
- La Clonación de La Oveja Dolly Explicada en ClaseDocumento2 páginasLa Clonación de La Oveja Dolly Explicada en ClaseUna Nueva Vida En NphAún no hay calificaciones
- Clase 13 CNDocumento6 páginasClase 13 CNTania Alvarez LlorenteAún no hay calificaciones
- Patologia Sylabo Upao VDocumento11 páginasPatologia Sylabo Upao VJuan Carlos SAún no hay calificaciones
- Clase 2 .Documento41 páginasClase 2 .kristell alvarado peñaAún no hay calificaciones
- Formato de Entrega Tarea 4Documento4 páginasFormato de Entrega Tarea 4alejandra guerreroAún no hay calificaciones