También podría gustarte
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Tema 12Documento6 páginasTema 12sandraAún no hay calificaciones
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- 02a Transcripción en ProcariontesDocumento9 páginas02a Transcripción en ProcariontesMaFe CamachoAún no hay calificaciones
- Transcripcion y TraduccionDocumento22 páginasTranscripcion y TraduccionChristian Correa VerdeAún no hay calificaciones
- 12a-Síntesis de Proteínas-Transcripción y TraduccDocumento48 páginas12a-Síntesis de Proteínas-Transcripción y TraduccJose HerediaAún no hay calificaciones
- Tema 2 GeneticaDocumento29 páginasTema 2 GeneticaSara MendezAún no hay calificaciones
- TRANSCRIPCIÓNDocumento19 páginasTRANSCRIPCIÓNjhon_4548Aún no hay calificaciones
- Clase Transcripción y Traduccion MaestriaDocumento215 páginasClase Transcripción y Traduccion MaestriaRobert CaballeroAún no hay calificaciones
- Síntesis de Proteínas (Transcripción y Traducción) y Código GenéticoDocumento33 páginasSíntesis de Proteínas (Transcripción y Traducción) y Código GenéticoMaria RosseAún no hay calificaciones
- Proceso de TranscripciónDocumento17 páginasProceso de TranscripciónKarina Alejandra Cervantes RoblesAún no hay calificaciones
- ARN y Sus CaracterísticasDocumento41 páginasARN y Sus CaracterísticasNazareth EspinozaAún no hay calificaciones
- Transcripcion Del DnaDocumento3 páginasTranscripcion Del DnaBrayan MercadoAún no hay calificaciones
- Tema 16.transcripcion PDFDocumento37 páginasTema 16.transcripcion PDFJoaquín Castro Orellana100% (1)
- Metabolismo Del ARNDocumento20 páginasMetabolismo Del ARNDavid AnlaAún no hay calificaciones
- Clase 9. Transcripción ARNDocumento52 páginasClase 9. Transcripción ARNChristian Robalino TorresAún no hay calificaciones
- Flujo de La Información GenéticaDocumento90 páginasFlujo de La Información GenéticaDayana Giraldo GAún no hay calificaciones
- TranscripciónDocumento8 páginasTranscripciónCarolina Rivera ReyAún no hay calificaciones
- TranscripcionDocumento9 páginasTranscripcionlina victoria garciaAún no hay calificaciones
- Transcripción y TraducciónDocumento80 páginasTranscripción y TraducciónThalia Ludeña VelardeAún no hay calificaciones
- Qué Es Un GenomaDocumento9 páginasQué Es Un GenomaemilyAún no hay calificaciones
- TranscripciónDocumento28 páginasTranscripciónAurelia Gonzales Calle100% (2)
- Repaso de Biología MolecularDocumento39 páginasRepaso de Biología MolecularhielitousAún no hay calificaciones
- Transcripción y Biosintesis Del ArnDocumento12 páginasTranscripción y Biosintesis Del ArnPaul CapellanAún no hay calificaciones
- Tema 8 TranscripcionDocumento15 páginasTema 8 Transcripcionjulian andresAún no hay calificaciones
- Síntesis y Maduración Del RnaDocumento7 páginasSíntesis y Maduración Del RnaJessicaChumbesAún no hay calificaciones
- Metabolismo Del RNADocumento3 páginasMetabolismo Del RNAJordy EdwardAún no hay calificaciones
- 2 Parcial de Genã©ticaDocumento17 páginas2 Parcial de Genã©ticarey reyesAún no hay calificaciones
- Semana 12 Expresión GenéticaDocumento34 páginasSemana 12 Expresión GenéticaArian Pilar Huarcaya CarlosAún no hay calificaciones
- TranscripcionDocumento8 páginasTranscripcionDeboraAún no hay calificaciones
- Resumen Del Capitulo 7 de Biologia MolecularDocumento7 páginasResumen Del Capitulo 7 de Biologia MolecularAlison TerrazasAún no hay calificaciones
- TranscripcionDocumento47 páginasTranscripcionFABIANA ANDREA DAZA ROCHAAún no hay calificaciones
- Unidad 3 TranscripcionDocumento31 páginasUnidad 3 TranscripcionAngelAún no hay calificaciones
- Transcripcion en ProcariotasDocumento16 páginasTranscripcion en ProcariotasSERGIO MANCERAAún no hay calificaciones
- Transcripción. Traducción. ReplicaciónDocumento93 páginasTranscripción. Traducción. ReplicaciónAlexa RodríguezAún no hay calificaciones
- Clase 5 y 6Documento64 páginasClase 5 y 6MatíasAún no hay calificaciones
- TRANSCRIPCIONDocumento20 páginasTRANSCRIPCIONJorgeLaTorreNavarroAún no hay calificaciones
- Biología MolecularDocumento8 páginasBiología MolecularErikaAún no hay calificaciones
- Transcripcion y TraduccionDocumento64 páginasTranscripcion y TraduccionFABIANA ANDREA DAZA ROCHAAún no hay calificaciones
- TEMA 7 DesgrabadoDocumento17 páginasTEMA 7 DesgrabadoCamila ObregonAún no hay calificaciones
- Semana 12 TeoriaDocumento38 páginasSemana 12 TeoriaLeonardo MarrufoAún no hay calificaciones
- Tema 10 - La Genética Molecular II. Expresión y Regulación de La Información GenéticaDocumento32 páginasTema 10 - La Genética Molecular II. Expresión y Regulación de La Información GenéticaInés JimenezAún no hay calificaciones
- Taller 2 - Transc Traducción.2019Documento20 páginasTaller 2 - Transc Traducción.2019LOL X Repleys ElmoshtaykersAún no hay calificaciones
- Informe TranscripcionDocumento12 páginasInforme TranscripcionAngieSagumaMoreno100% (1)
- Equipo 3 GeneticaaDocumento45 páginasEquipo 3 GeneticaaSeñor PavoAún no hay calificaciones
- TranscripciónDocumento13 páginasTranscripciónBibiana VarónAún no hay calificaciones
- Transcripcion Del AdnDocumento38 páginasTranscripcion Del AdnElenaBarbosaAún no hay calificaciones
- TRANSCRIPCIÓNDocumento21 páginasTRANSCRIPCIÓNDaniel CarpioAún no hay calificaciones
- Metabolismo de RNADocumento40 páginasMetabolismo de RNACesar Pariona100% (1)
- Licenciatura en NutricionDocumento8 páginasLicenciatura en NutricionDenisse G Tamay YervesAún no hay calificaciones
- Clase 6 Genetica Molecular IIDocumento8 páginasClase 6 Genetica Molecular IIcyber pgAún no hay calificaciones
- Transcripcion y TraduccionDocumento54 páginasTranscripcion y Traduccionjj1064Aún no hay calificaciones
- Diferencias Entre Rna y DnaDocumento7 páginasDiferencias Entre Rna y DnaCata ValenciaAún no hay calificaciones
- Tema 10Documento9 páginasTema 10Laura Xi Baratto PérezAún no hay calificaciones
- TrasncripciónDocumento25 páginasTrasncripciónGabriel TorresAún no hay calificaciones
- TRANSCRIPCIÓNDocumento5 páginasTRANSCRIPCIÓNrommy danferAún no hay calificaciones
- Transcripcion Del DNADocumento8 páginasTranscripcion Del DNAAngelique CassabAún no hay calificaciones
- Transcripcion Del Adn - ExposicionDocumento31 páginasTranscripcion Del Adn - ExposicionJuly Lalangui Rivera100% (4)
- Obj. 2 - TranscripciónDocumento5 páginasObj. 2 - TranscripciónCristinaAún no hay calificaciones
- ArnDocumento28 páginasArnSharini_Mu_oz_137Aún no hay calificaciones
- Elongacion MuscularDocumento12 páginasElongacion MuscularJuliEstradaAún no hay calificaciones
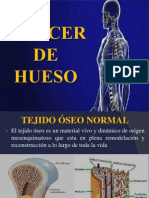
- Cáncer de HuesoDocumento24 páginasCáncer de HuesoVanessa Tapia Quiroz100% (1)
- CTO Endocrinología CushingDocumento6 páginasCTO Endocrinología CushingDaniel MitreAún no hay calificaciones
- Examen General de HecesDocumento23 páginasExamen General de HecesLuz FloresAún no hay calificaciones
- Microsoft Word Solicitud Informe Externo DefinitivoDocumento3 páginasMicrosoft Word Solicitud Informe Externo DefinitivoferAún no hay calificaciones
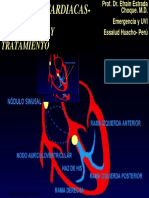
- Cardio ArritmiasDocumento85 páginasCardio ArritmiasFabio Hernàndez SantiagoAún no hay calificaciones
- EpidemiologiaDocumento4 páginasEpidemiologiaJhon100% (1)
- Antibioticos en OdontologiaDocumento11 páginasAntibioticos en Odontologiakarla0% (1)
- Educacion EspecialDocumento6 páginasEducacion EspecialZeroTheBesTTAún no hay calificaciones
- Introducción A La EmbriologíaDocumento19 páginasIntroducción A La EmbriologíaLiz Calderon MolloAún no hay calificaciones
- Triptico Voz PDFDocumento2 páginasTriptico Voz PDFLucas BenjamínAún no hay calificaciones
- Patologia de La Via BiliarDocumento74 páginasPatologia de La Via BiliarCarloos SemperAún no hay calificaciones
- MicrobiosDocumento1 páginaMicrobiosAlee AppAún no hay calificaciones
- Segundo LugarDocumento21 páginasSegundo LugarYulizaFT100% (1)
- Las MacromoléculasDocumento2 páginasLas MacromoléculasFrancisco FelixAún no hay calificaciones
- Rueda de La Resiliencia Escolar y Ejemplos de AplicacionDocumento6 páginasRueda de La Resiliencia Escolar y Ejemplos de Aplicacionpablo190381Aún no hay calificaciones
- Mastitis Montana SADocumento30 páginasMastitis Montana SALeodanRiojasHuamanAún no hay calificaciones
- Informe 1. Fitopatología. Rueda, Delgado y GalvisDocumento9 páginasInforme 1. Fitopatología. Rueda, Delgado y GalvisLaura Johanna Galvis RuizAún no hay calificaciones
- Evoluciones de Psicologia Leddys Lopez AutismoDocumento10 páginasEvoluciones de Psicologia Leddys Lopez AutismoINGRID PUMAREJOAún no hay calificaciones
- Práctica 10Documento11 páginasPráctica 10Aranzy Judith Flores LeosAún no hay calificaciones
- Unidad 1 Importancia de La Psicobiologc3adaDocumento13 páginasUnidad 1 Importancia de La Psicobiologc3adaWENDY MENDOZAAún no hay calificaciones
- Gastroenterología 2014 - 01 PDFDocumento30 páginasGastroenterología 2014 - 01 PDFJose RamonAún no hay calificaciones
- Ejercicio Bloque 3 Tema 3.1.1Documento2 páginasEjercicio Bloque 3 Tema 3.1.1ConsueloAún no hay calificaciones
- Caso 1Documento4 páginasCaso 1Frida VAAún no hay calificaciones
- El Aparato ExcretorDocumento20 páginasEl Aparato ExcretorfioriAún no hay calificaciones
- Reino ProtistaDocumento8 páginasReino ProtistagreiberAún no hay calificaciones
- DIGESA - Recomendaciones para Comprar JuguetesDocumento2 páginasDIGESA - Recomendaciones para Comprar JuguetesR. S.Aún no hay calificaciones
- Estimulacion TempranaDocumento16 páginasEstimulacion TempranaSolcito Bü100% (1)
- Rias 20 MarzoDocumento47 páginasRias 20 MarzoAnonymous rTaxZIjmKxAún no hay calificaciones
- Primer Laboratorio Enzimologiìa RetroDocumento7 páginasPrimer Laboratorio Enzimologiìa RetroMariana BecerrilAún no hay calificaciones
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Homo antecessor: El nacimiento de una especieDe EverandHomo antecessor: El nacimiento de una especieCalificación: 5 de 5 estrellas5/5 (1)
- Liberación del trauma: Perdón y temblor es el caminoDe EverandLiberación del trauma: Perdón y temblor es el caminoCalificación: 4 de 5 estrellas4/5 (4)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Flujo de fluidos e intercambio de calorDe EverandFlujo de fluidos e intercambio de calorCalificación: 1 de 5 estrellas1/5 (1)
- Cerebro y silencio: Las claves de la creatividad y la serenidadDe EverandCerebro y silencio: Las claves de la creatividad y la serenidadCalificación: 5 de 5 estrellas5/5 (2)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- Cuerpo Tóxico: Como Liberar Tu Cuerpo De Las Toxinas Externas E Internas, Y Evitar Asi Los Efectos De Los Radicales LibresDe EverandCuerpo Tóxico: Como Liberar Tu Cuerpo De Las Toxinas Externas E Internas, Y Evitar Asi Los Efectos De Los Radicales LibresCalificación: 5 de 5 estrellas5/5 (2)
- Anatomía del caballo: Guía práctica ilustradaDe EverandAnatomía del caballo: Guía práctica ilustradaCalificación: 4 de 5 estrellas4/5 (9)
- Sesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesDe EverandSesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Psicología forense: Estudio de la mente criminalDe EverandPsicología forense: Estudio de la mente criminalCalificación: 4.5 de 5 estrellas4.5/5 (18)
- Las Enfermedades comienzan y terminan en tu mente: Una guía para la autosanaciónDe EverandLas Enfermedades comienzan y terminan en tu mente: Una guía para la autosanaciónCalificación: 4 de 5 estrellas4/5 (4)
- Las matemáticas de la biología: De las celdas de las abejas a las simetrías de los virusDe EverandLas matemáticas de la biología: De las celdas de las abejas a las simetrías de los virusCalificación: 4 de 5 estrellas4/5 (1)
- Química orgánica: ejercicios de aplicaciónDe EverandQuímica orgánica: ejercicios de aplicaciónCalificación: 5 de 5 estrellas5/5 (3)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Grasas inteligentes: Come más grasa. Pierde más peso. Mantente más sanoDe EverandGrasas inteligentes: Come más grasa. Pierde más peso. Mantente más sanoCalificación: 4 de 5 estrellas4/5 (10)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- El nacimiento del pensamiento científico: Anaximandro de MiletoDe EverandEl nacimiento del pensamiento científico: Anaximandro de MiletoCalificación: 4.5 de 5 estrellas4.5/5 (4)
- Contacto & Relación en Psicoterapia: Reflexiones sobre terapia GestaltDe EverandContacto & Relación en Psicoterapia: Reflexiones sobre terapia GestaltCalificación: 3 de 5 estrellas3/5 (2)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Un científico en el lavadero: Manchas, olores, ciencia, tecnología y suciedadDe EverandUn científico en el lavadero: Manchas, olores, ciencia, tecnología y suciedadCalificación: 5 de 5 estrellas5/5 (2)
- UF0085 - Diagnóstico estético facial y corporalDe EverandUF0085 - Diagnóstico estético facial y corporalCalificación: 4 de 5 estrellas4/5 (5)
- Epigenoma: para cuidar tu cuerpo y tu vidaDe EverandEpigenoma: para cuidar tu cuerpo y tu vidaCalificación: 4 de 5 estrellas4/5 (5)