También podría gustarte
- Uso del cannabis en el manejo del dolorDe EverandUso del cannabis en el manejo del dolorCalificación: 3 de 5 estrellas3/5 (1)
- Fisiologia Dolor PDFDocumento59 páginasFisiologia Dolor PDFMeyling García100% (1)
- Neurofisiologia Del DolorDocumento21 páginasNeurofisiologia Del DolorLety Catari100% (1)
- Dolor Lmixto Cambiando ParadigmasDocumento9 páginasDolor Lmixto Cambiando ParadigmasSindy AguilarAún no hay calificaciones
- Dolor Fisiologia - MergedDocumento5 páginasDolor Fisiologia - MergedYuliethmendezmAún no hay calificaciones
- El DolorDocumento16 páginasEl DolorAurora SosaAún no hay calificaciones
- Dolor CompletoDocumento9 páginasDolor CompletoMelissa RivasAún no hay calificaciones
- Fisiofarmacología en AnestesiologíaDocumento7 páginasFisiofarmacología en AnestesiologíaSandra Gil EspinozaAún no hay calificaciones
- Clase 5 FisiologíaDocumento15 páginasClase 5 FisiologíaAlonso molinaAún no hay calificaciones
- Neuroplasticidad y DolorDocumento5 páginasNeuroplasticidad y Dolorkine_hector100% (1)
- Transmisión Del Estimulo Del Dolor 1Documento9 páginasTransmisión Del Estimulo Del Dolor 1Nerly JarabaAún no hay calificaciones
- DOLOR LizethDocumento40 páginasDOLOR LizethLizeth Yulieth Obando MorenoAún no hay calificaciones
- Wa0104.Documento19 páginasWa0104.grupodeamistad20Aún no hay calificaciones
- Dolorr FisiopatologiaDocumento16 páginasDolorr FisiopatologiaJAIRO GALINDOAún no hay calificaciones
- Mapa Conceptual Neurofisiologia Del Dolor ListoDocumento2 páginasMapa Conceptual Neurofisiologia Del Dolor Listoyasmir veliz100% (1)
- TEMA Fisiopatologia Del Dolor CURSO Fisiopatologia DOCENTE G.O.Documento31 páginasTEMA Fisiopatologia Del Dolor CURSO Fisiopatologia DOCENTE G.O.Alejandra Isabel Vargas PaucarAún no hay calificaciones
- Vias Del DolorDocumento45 páginasVias Del DolorAvelar Rios Brenda Escarlette100% (1)
- Dolor Agudo y CronicoDocumento21 páginasDolor Agudo y Cronicoclubdelquimico.blogspot.com100% (3)
- El Dolor Estructuras Que IntervienenDocumento23 páginasEl Dolor Estructuras Que IntervienenMonica AlcantaraAún no hay calificaciones
- Fisiopatología Del DolorDocumento9 páginasFisiopatología Del DolorCami RomeroAún no hay calificaciones
- Dolor Dolor PDFDocumento4 páginasDolor Dolor PDFSebastián VilchesAún no hay calificaciones
- Dolor en Pediatría: Fisiopatología y ValoraciónDocumento10 páginasDolor en Pediatría: Fisiopatología y ValoraciónPapu FloresAún no hay calificaciones
- FISIOTERAPIA DEL DOLOR ParcialDocumento7 páginasFISIOTERAPIA DEL DOLOR ParcialAlejandra LAún no hay calificaciones
- Neurofisiologia Del DolorDocumento21 páginasNeurofisiologia Del DolorCHARLES100% (1)
- BASES FISIOPATOLÓGICAS DEL DOLOR Acta Médica Peruana, Mayo-Agosto, Año/vol. 24, Número 002Documento5 páginasBASES FISIOPATOLÓGICAS DEL DOLOR Acta Médica Peruana, Mayo-Agosto, Año/vol. 24, Número 002Paloma MendozaAún no hay calificaciones
- Tipos de Dolor y Nociceptores-1Documento23 páginasTipos de Dolor y Nociceptores-1Wendy AlcantaraAún no hay calificaciones
- Neurofisiologia DesarrolloDocumento19 páginasNeurofisiologia DesarrolloMilagros M. PaucarAún no hay calificaciones
- DOLORDocumento4 páginasDOLORMelissa RivasAún no hay calificaciones
- Documento Sin TítuloDocumento3 páginasDocumento Sin TítuloAudiAún no hay calificaciones
- Analgesia y Pre-AnestesiaDocumento55 páginasAnalgesia y Pre-AnestesiaAngie HernándezAún no hay calificaciones
- Dolor HarrisonDocumento178 páginasDolor Harrisondaniela cAún no hay calificaciones
- Aines FarmacologiaDocumento9 páginasAines FarmacologiaRTNAún no hay calificaciones
- 17.principios Basicos Acerca Del Manejo Del DolorDocumento27 páginas17.principios Basicos Acerca Del Manejo Del DolorTIPO C CATAMAYOAún no hay calificaciones
- Fisiopatología Del DolorDocumento24 páginasFisiopatología Del DoloririsvillalobosmorantAún no hay calificaciones
- Argente DolorDocumento29 páginasArgente DolorDAMARIS CRISTELL AGUILAR SANCHEZAún no hay calificaciones
- Universidad Latina de Panamá Facultad de Ciencias Médicas Y de La SaludDocumento18 páginasUniversidad Latina de Panamá Facultad de Ciencias Médicas Y de La Saluddaniela morenoAún no hay calificaciones
- Cuadro DolorDocumento1 páginaCuadro DolorDianaAún no hay calificaciones
- Fisiopatologia Del DolorDocumento3 páginasFisiopatologia Del DolorAngélica María AponteAún no hay calificaciones
- Steeds2009 en EsDocumento5 páginasSteeds2009 en EsrosangelaAún no hay calificaciones
- Dolor PDFDocumento39 páginasDolor PDFSaandy GC100% (1)
- Fisiologia Del DolorDocumento14 páginasFisiologia Del DolorSoAr JCAún no hay calificaciones
- 1 Dolor ConceptosDocumento5 páginas1 Dolor ConceptosAlix RodríguezAún no hay calificaciones
- Tema 3 - Fisiopatologia Del DolorDocumento6 páginasTema 3 - Fisiopatologia Del DolorRodrigo LeesAún no hay calificaciones
- Fisiopatologia Del DolorDocumento34 páginasFisiopatologia Del DolorAnto Flores100% (4)
- DolorDocumento18 páginasDolorVivi CardonaAún no hay calificaciones
- UBA UD 2 Neurofisio - Clase 3 - Sistema Sensorial. DolorDocumento63 páginasUBA UD 2 Neurofisio - Clase 3 - Sistema Sensorial. DolorMILAGROS BELEN GILAún no hay calificaciones
- FISIOLOGÍA DEL DOLOR PARA EL CURSO 2021.en - EsDocumento5 páginasFISIOLOGÍA DEL DOLOR PARA EL CURSO 2021.en - EsMauricio MarínAún no hay calificaciones
- Fisiopatología Del DolorDocumento4 páginasFisiopatología Del DolorLubego870% (1)
- Opioid EsDocumento5 páginasOpioid EsSHARON LENNY MAMANI TONCONIAún no hay calificaciones
- Teorias Del DolorDocumento13 páginasTeorias Del DolorHillary Vasquez100% (1)
- Clase 3 2022 - GcnaDocumento38 páginasClase 3 2022 - Gcnacecilia cortezAún no hay calificaciones
- Infografia DolorDocumento1 páginaInfografia DolorGERARDO ALEXIS MARTINEZ NATIVIDADAún no hay calificaciones
- Neurofisiologia Dolor PDFDocumento27 páginasNeurofisiologia Dolor PDFkine_hectorAún no hay calificaciones
- Fisiopatologia Del DolorDocumento20 páginasFisiopatologia Del DolorEdgarQuiñonesAún no hay calificaciones
- Receptores Pulpares, Periodontales y Articulares (Cristi)Documento21 páginasReceptores Pulpares, Periodontales y Articulares (Cristi)Angela Alvarado DelgadoAún no hay calificaciones
- PAPER DOLOR EspañolDocumento8 páginasPAPER DOLOR EspañolAlejandro TorresAún no hay calificaciones
- Dolor AgudoDocumento18 páginasDolor AgudoAide Gonzalez SandovalAún no hay calificaciones
- Sensaciones Percibidas Por El CuerpoDocumento13 páginasSensaciones Percibidas Por El CuerpoGisselle VelásquezAún no hay calificaciones
- DesarrolloDocumento1 páginaDesarrollomia sanchezAún no hay calificaciones
- Jimeno Crimen PasionalDocumento29 páginasJimeno Crimen PasionalCatalina GonzalezAún no hay calificaciones
- Tratamiento de Quemaduras Térmicas Menores - UpToDateDocumento17 páginasTratamiento de Quemaduras Térmicas Menores - UpToDateJose PiñaAún no hay calificaciones
- Test de Tipología de JungDocumento9 páginasTest de Tipología de JungGenesis Tavarez MoralwsAún no hay calificaciones
- Antropología de Las Emociones y Los SentimientosDocumento3 páginasAntropología de Las Emociones y Los SentimientososwaldosaenzAún no hay calificaciones
- Discriminación Película FRAGMENTADO Psicopatología de AtenciónDocumento3 páginasDiscriminación Película FRAGMENTADO Psicopatología de AtenciónJohana Corzo Rodriguez100% (1)
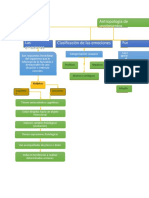
- Clasificacion Del Dolor Sistema Nervioso 1Documento9 páginasClasificacion Del Dolor Sistema Nervioso 1Jefferson Arones RodriguezAún no hay calificaciones
- Guia Manejo Del Dolor en EnfermeriaDocumento16 páginasGuia Manejo Del Dolor en EnfermeriaJean Paul Castro CárdenasAún no hay calificaciones
- Analgesia Multimodal EquinosDocumento65 páginasAnalgesia Multimodal EquinosAna Murillo Quevedo100% (1)
- Efectividad de La Neuroestimulación Eléctrica Percutánea en La Desactivación de Los Puntos Gatillo Miofasciales en La Lumbalgia CrónicaDocumento4 páginasEfectividad de La Neuroestimulación Eléctrica Percutánea en La Desactivación de Los Puntos Gatillo Miofasciales en La Lumbalgia CrónicaDaniel Alejandro Vargas PérezAún no hay calificaciones
- PDF 20230410 173505 0000Documento46 páginasPDF 20230410 173505 0000Katheryne GómezAún no hay calificaciones
- 2.sindrome de Espalda FallidaDocumento32 páginas2.sindrome de Espalda FallidaAlondra BenitesAún no hay calificaciones
- Bio Psico Socio Cultural CelosDocumento145 páginasBio Psico Socio Cultural Celoscesarpsychology100% (1)
- Cómo Ser FelizDocumento4 páginasCómo Ser FelizValentina FonsecaAún no hay calificaciones
- La Técnica Del EspejoDocumento2 páginasLa Técnica Del EspejoPatricia Edith Martinez PortilloAún no hay calificaciones
- Cuáles Son Los 17 Tipos de Sentimientos Del Ser HumanoDocumento19 páginasCuáles Son Los 17 Tipos de Sentimientos Del Ser HumanoMatilde ForeroAún no hay calificaciones
- Fernández-Abascal, E. (2010) - Procesamiento EmocionalDocumento47 páginasFernández-Abascal, E. (2010) - Procesamiento EmocionalEliana CaroAún no hay calificaciones
- Damasio La Mente HumanaDocumento11 páginasDamasio La Mente HumanaIgnacio VictorianoAún no hay calificaciones
- Balotario de Psicologia GeneralDocumento27 páginasBalotario de Psicologia GeneralMartin Müller50% (2)
- Programa Cuidados Paliativos - RAM INCan 2015Documento5 páginasPrograma Cuidados Paliativos - RAM INCan 2015Stanley Wu TaiAún no hay calificaciones
- MONOGRAFIADocumento51 páginasMONOGRAFIAgiovana chura100% (3)
- SedonaDocumento7 páginasSedonaapi-3854058Aún no hay calificaciones
- Inventario de Preferencias Personales de Edwards ADocumento42 páginasInventario de Preferencias Personales de Edwards AClaudioAún no hay calificaciones
- Sesion de ClasesDocumento14 páginasSesion de ClasesrobertomalpartidaleonAún no hay calificaciones
- Fisiopatologia Del DolorDocumento34 páginasFisiopatologia Del DolorAnto Flores100% (4)
- Sentimiento de CulpaDocumento4 páginasSentimiento de CulpaJonathan AraujoAún no hay calificaciones
- Monografía Con Índice.Documento13 páginasMonografía Con Índice.RONALD RUBEN TTITO MAMANIAún no hay calificaciones
- Dr. Dennis López Naranjo Neurología Pediátrica Hospital Infantil Federico GomezDocumento50 páginasDr. Dennis López Naranjo Neurología Pediátrica Hospital Infantil Federico GomezvictoriaAún no hay calificaciones
- Pienso, Siento y Actúo - Siento, Pienso, Actúo - o Actúo, Siento, PiensoDocumento9 páginasPienso, Siento y Actúo - Siento, Pienso, Actúo - o Actúo, Siento, PiensovatilanoAún no hay calificaciones
- Segundo Parcial de PsicopatologíaDocumento19 páginasSegundo Parcial de PsicopatologíaJosé Luis PadgettAún no hay calificaciones
- Cómo conversar con cualquiera: Descubre cómo ser la persona más interesante del lugar y a destruir la ansiedad social permanentementeDe EverandCómo conversar con cualquiera: Descubre cómo ser la persona más interesante del lugar y a destruir la ansiedad social permanentementeCalificación: 4 de 5 estrellas4/5 (81)
- Altas Dosis: Los efectos milagrosos de dosis extremadamente altas de vitamina D3. El gran secreto que la industria farmacéutica quiere ocultarle.De EverandAltas Dosis: Los efectos milagrosos de dosis extremadamente altas de vitamina D3. El gran secreto que la industria farmacéutica quiere ocultarle.Calificación: 5 de 5 estrellas5/5 (4)
- Terapia cognitiva del comportamiento: Descubre cómo despejar tu cerebro con la TCC. Aprender a superar el miedo y la ansiedad, la depresiónDe EverandTerapia cognitiva del comportamiento: Descubre cómo despejar tu cerebro con la TCC. Aprender a superar el miedo y la ansiedad, la depresiónCalificación: 5 de 5 estrellas5/5 (3)
- Sana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónDe EverandSana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónCalificación: 5 de 5 estrellas5/5 (4)
- Terapia cognitiva: Conceptos básicos y profundizaciónDe EverandTerapia cognitiva: Conceptos básicos y profundizaciónCalificación: 5 de 5 estrellas5/5 (1)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- Psicología y trastornos de los niños en la edad evolutiva: Qué son y cómo funcionanDe EverandPsicología y trastornos de los niños en la edad evolutiva: Qué son y cómo funcionanCalificación: 5 de 5 estrellas5/5 (1)
- Cómo estimular el cerebro del niño: 100 ejercicios para potenciar la concentración, la memoria y otras funciones ejecutivasDe EverandCómo estimular el cerebro del niño: 100 ejercicios para potenciar la concentración, la memoria y otras funciones ejecutivasCalificación: 4.5 de 5 estrellas4.5/5 (12)
- Autismo: guía para padres sobre el trastorno del espectro autista En españolDe EverandAutismo: guía para padres sobre el trastorno del espectro autista En españolCalificación: 4.5 de 5 estrellas4.5/5 (4)
- Dinámicas familiares a través de la vida de José: La túnica del PadreDe EverandDinámicas familiares a través de la vida de José: La túnica del PadreCalificación: 5 de 5 estrellas5/5 (2)
- Neurociencia para vencer la depresión: La esprial ascendenteDe EverandNeurociencia para vencer la depresión: La esprial ascendenteCalificación: 4.5 de 5 estrellas4.5/5 (10)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Cardiología y enfermedades cardiovascularesDe EverandCardiología y enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- Muchas Vidas, Muchos Sabios (Many Lives, Many Masters): (Many Lives, Many Masters)De EverandMuchas Vidas, Muchos Sabios (Many Lives, Many Masters): (Many Lives, Many Masters)Calificación: 4 de 5 estrellas4/5 (475)
- DMT: La molécula del espíritu: Las revolucionarias investigaciones de un médico sobre la biología de las experiencias místicas y cercanas a la muerteDe EverandDMT: La molécula del espíritu: Las revolucionarias investigaciones de un médico sobre la biología de las experiencias místicas y cercanas a la muerteCalificación: 4 de 5 estrellas4/5 (135)
- Shiatsu: Conozca las técnicas y tratamientos de una de las terapias más antiguasDe EverandShiatsu: Conozca las técnicas y tratamientos de una de las terapias más antiguasAún no hay calificaciones
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Mindfulness: Cómo hacer menos y ser más conscientes puede ayudarnos a vivir mejor en este mundo estresadoDe EverandMindfulness: Cómo hacer menos y ser más conscientes puede ayudarnos a vivir mejor en este mundo estresadoCalificación: 4.5 de 5 estrellas4.5/5 (9)
- Neuropsicología: Los fundamentos de la materiaDe EverandNeuropsicología: Los fundamentos de la materiaCalificación: 5 de 5 estrellas5/5 (1)
- Fisiopatología de las enfermedades cardiovascularesDe EverandFisiopatología de las enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- Prescripción de ejercico físico para la saludDe EverandPrescripción de ejercico físico para la saludCalificación: 5 de 5 estrellas5/5 (1)
- Psicópatas seriales: Un recorrido por su oscura e inquietante naturalezaDe EverandPsicópatas seriales: Un recorrido por su oscura e inquietante naturalezaCalificación: 4 de 5 estrellas4/5 (3)
- TDAH en Adultos. Cómo Reconocer y Tratar a un Adulto con TDAH en 30 Fáciles PasosDe EverandTDAH en Adultos. Cómo Reconocer y Tratar a un Adulto con TDAH en 30 Fáciles PasosCalificación: 4 de 5 estrellas4/5 (8)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)