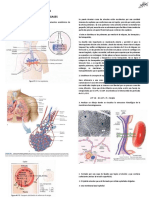
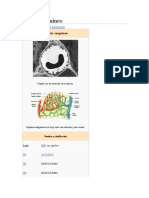
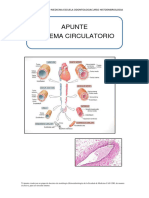
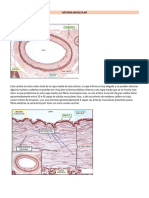
También podría gustarte
- CapilaresDocumento7 páginasCapilaresJhon Ji MaAún no hay calificaciones
- La Circulacion de La PulpaDocumento22 páginasLa Circulacion de La Pulpafernanda belalcazar100% (1)
- Intercambio y Transporte de Gases Tema 26Documento3 páginasIntercambio y Transporte de Gases Tema 26Miguel AmbrocioAún no hay calificaciones
- Micro Circ U Laci OnDocumento13 páginasMicro Circ U Laci OnrubenAún no hay calificaciones
- GadgasdgDocumento11 páginasGadgasdgwalmerAún no hay calificaciones
- Capilar Sanguíneo (Wikipedia)Documento3 páginasCapilar Sanguíneo (Wikipedia)Luis Soria TorresAún no hay calificaciones
- Vasos CapilaresDocumento3 páginasVasos CapilaresOrlando CalderaAún no hay calificaciones
- Perfusion Tisular en El Paciente Critico PDFDocumento13 páginasPerfusion Tisular en El Paciente Critico PDFAndyGcAún no hay calificaciones
- Características Histologicas de La Membrana ADocumento3 páginasCaracterísticas Histologicas de La Membrana AGaby Aldama100% (1)
- TEMA 2, Neurohistologia IDocumento9 páginasTEMA 2, Neurohistologia ICamila CastañoAún no hay calificaciones
- Célula EndotelialDocumento22 páginasCélula EndotelialKarla Cruz 06Aún no hay calificaciones
- CapilaresDocumento11 páginasCapilaresJosh MoralesAún no hay calificaciones
- Cadena RespiratoriaDocumento11 páginasCadena RespiratoriaAngelAún no hay calificaciones
- Endotelio Clase 04Documento10 páginasEndotelio Clase 04Ivonne Mercedes Gutierrez AtapaucarAún no hay calificaciones
- Tema 15 Sistema RespiratorioDocumento25 páginasTema 15 Sistema Respiratoriokim sun heeAún no hay calificaciones
- Umbral LácticoDocumento8 páginasUmbral LácticoManuel Ignacio Peña Urzua100% (1)
- Difusion RespiratoriaDocumento8 páginasDifusion RespiratoriaMaraby100% (1)
- Microcirculación e Intercambio CapilarDocumento21 páginasMicrocirculación e Intercambio CapilarEmilia GanzoAún no hay calificaciones
- TP PieuDocumento4 páginasTP PieuWalter E. GaviAún no hay calificaciones
- Cardiorespiratorio MicrocirculaciónDocumento43 páginasCardiorespiratorio MicrocirculaciónAndrew Emilio Castillo PachecoAún no hay calificaciones
- Tema 9 Circulación Capilar y Circulación LinfáticaDocumento12 páginasTema 9 Circulación Capilar y Circulación LinfáticaRosa SotoAún no hay calificaciones
- Capítulo 16 - La Microcirculación y El Sistema LinfáticoDocumento11 páginasCapítulo 16 - La Microcirculación y El Sistema LinfáticoMirianCamposAún no hay calificaciones
- Importancia Del Endotelio CapilarDocumento16 páginasImportancia Del Endotelio CapilarGlenn0% (2)
- Prueba de Biologia Posibles RespuestasDocumento9 páginasPrueba de Biologia Posibles Respuestass.lassagaAún no hay calificaciones
- Mecanismos+de+Funcionamiento+alveolar en EsDocumento16 páginasMecanismos+de+Funcionamiento+alveolar en EsDaniel Delgadillo CruzAún no hay calificaciones
- Esquema General de La Funcion PulmonarDocumento34 páginasEsquema General de La Funcion Pulmonaryessy32lAún no hay calificaciones
- 48 Circul OrganizacionDocumento5 páginas48 Circul OrganizacionEvangelina RuizAún no hay calificaciones
- Los EritrocitosDocumento9 páginasLos EritrocitosSisalima Nallely LilianaAún no hay calificaciones
- CapilarDocumento5 páginasCapilaranon_330975744Aún no hay calificaciones
- Micro Circ U Laci OnDocumento13 páginasMicro Circ U Laci Onkarla7barrenecheaAún no hay calificaciones
- Que Es La FisiologiaDocumento9 páginasQue Es La FisiologiaYulissa Bañales CastañedaAún no hay calificaciones
- Trastornos Vasculares y Trombosis - Patologia General VeterinariaDocumento21 páginasTrastornos Vasculares y Trombosis - Patologia General VeterinariaBerna CelestinoAún no hay calificaciones
- Capilar SanguíneoDocumento3 páginasCapilar SanguíneoLaura ElizabethAún no hay calificaciones
- PRÁCTICA 6. Tejido Adiposo y Sistema CirculatorioDocumento11 páginasPRÁCTICA 6. Tejido Adiposo y Sistema CirculatorioPaulina CarmonaAún no hay calificaciones
- Sistema circulatorio: guía para estudiar las partes y funciones de la sangre, el corazón y los vasos sanguíneosDocumento5 páginasSistema circulatorio: guía para estudiar las partes y funciones de la sangre, el corazón y los vasos sanguíneosCruz Bisbal100% (1)
- Clinical Liver Disease - 2014 - Krishna - Anatom A Microsc Pica Del H GadoDocumento4 páginasClinical Liver Disease - 2014 - Krishna - Anatom A Microsc Pica Del H GadoMonserrat MacíasAún no hay calificaciones
- Sistema Circulatorio c21 - 240227 - 195308Documento11 páginasSistema Circulatorio c21 - 240227 - 195308Juan Antonio Parra OrtegaAún no hay calificaciones
- 3dep A2 KybrDocumento3 páginas3dep A2 KybrKyrs MarichieAún no hay calificaciones
- RESUMEN - Fisiología Respiratorio y GastroDocumento213 páginasRESUMEN - Fisiología Respiratorio y Gastrojoelito_77Aún no hay calificaciones
- Lectura Endotelio Vascular PDFDocumento6 páginasLectura Endotelio Vascular PDFSofiaMarmolejoAún no hay calificaciones
- Eritrocitos: transporte de O2 y CO2Documento2 páginasEritrocitos: transporte de O2 y CO2Paula VarelaAún no hay calificaciones
- PANCREASDocumento14 páginasPANCREASAlfredo BenitesAún no hay calificaciones
- Cuestionario de Sistema Respiratorio NAOMI SALTOSDocumento7 páginasCuestionario de Sistema Respiratorio NAOMI SALTOSNAOMI REBECA SALTOS CANDELAAún no hay calificaciones
- S4 (T12) - Barrera Hematoencefálica y Fisiología Del LCRDocumento6 páginasS4 (T12) - Barrera Hematoencefálica y Fisiología Del LCRMarcelo HuamanAún no hay calificaciones
- Fisiología Respiratoria BásicaDocumento26 páginasFisiología Respiratoria BásicaPeter Amaya-QuiñonesAún no hay calificaciones
- Capítulo 11 - Trastornos Cardiovasculares - Enfermedad VascularDocumento39 páginasCapítulo 11 - Trastornos Cardiovasculares - Enfermedad VascularChávez Ramírez Juana SofíaAún no hay calificaciones
- 1ra Expo Shock CirugiaDocumento98 páginas1ra Expo Shock CirugiaFabiany Valladares NoleAún no hay calificaciones
- Tipeo HistoDocumento9 páginasTipeo HistoSofía GuzmánAún no hay calificaciones
- Cuestionario Sobre Diversidad Celular.Documento7 páginasCuestionario Sobre Diversidad Celular.Andreína HigueraAún no hay calificaciones
- Tejido ConectivoDocumento19 páginasTejido ConectivoJulianaSarmientoAún no hay calificaciones
- Capilares sanguíneos: características, tipos y funciónDocumento14 páginasCapilares sanguíneos: características, tipos y funciónDioni SarayaAún no hay calificaciones
- Histología (Segundo Parcial)Documento65 páginasHistología (Segundo Parcial)Meme Nella CastroAún no hay calificaciones
- Cuestionario Sistemas CardiovascularDocumento4 páginasCuestionario Sistemas CardiovascularJahaira CapellanAún no hay calificaciones
- T6 LaRespiraciónDocumento12 páginasT6 LaRespiraciónNikhil PaulAún no hay calificaciones
- Anemia Falciforme 2023Documento20 páginasAnemia Falciforme 2023CarlaAún no hay calificaciones
- Hhhemodinamica Sanguinea!!!!!!!!!Documento10 páginasHhhemodinamica Sanguinea!!!!!!!!!Daniela CarvajalAún no hay calificaciones
- Clase 7 EritropoyesisDocumento8 páginasClase 7 EritropoyesisLizy OdontoAún no hay calificaciones
- Sistema respiratorioDocumento6 páginasSistema respiratorioGabyAún no hay calificaciones
- Anestesia Secretos 4 EdicionDocumento20 páginasAnestesia Secretos 4 EdicionjuanAún no hay calificaciones
- aformularioregistro01_impl (1)Documento2 páginasaformularioregistro01_impl (1)Carlos VargasAún no hay calificaciones
- EVENTOS-OFICIALES-FOLKLORE-MUJERES-TITULARES-Y-SUPLENTESDocumento6 páginasEVENTOS-OFICIALES-FOLKLORE-MUJERES-TITULARES-Y-SUPLENTESCarlos VargasAún no hay calificaciones
- SIMEN ASCENSORES 17194Documento1 páginaSIMEN ASCENSORES 17194Carlos VargasAún no hay calificaciones
- Mant Gaston Faltantes-1Documento1 páginaMant Gaston Faltantes-1Carlos VargasAún no hay calificaciones
- Ups Ult 1000 3000VTDocumento2 páginasUps Ult 1000 3000VTCarlos VargasAún no hay calificaciones
- ManualUPSPolarisXION650 850Documento12 páginasManualUPSPolarisXION650 850Carlos Vargas100% (1)
- Ensayo de AntivaloresDocumento2 páginasEnsayo de AntivaloresJennifer Pope100% (2)
- Pragma-dialectical Approach to Argument AnalysisDocumento13 páginasPragma-dialectical Approach to Argument AnalysisPedro PosadaAún no hay calificaciones
- Procedimientos de la Unidad Educativa Adventista Franz TamayoDocumento8 páginasProcedimientos de la Unidad Educativa Adventista Franz TamayoCloverwatch TV0% (1)
- Intervención Fonoaudiológica en Pacientes Adultos Con Déficit Intelectual.Documento6 páginasIntervención Fonoaudiológica en Pacientes Adultos Con Déficit Intelectual.Sofía MardonesAún no hay calificaciones
- Qué Aire NecesitoDocumento2 páginasQué Aire NecesitoMalka Sarmiento100% (1)
- Momento DipolarDocumento9 páginasMomento DipolarLaura GuidoAún no hay calificaciones
- Psicopatología: definición, objetivos y manuales de clasificaciónDocumento2 páginasPsicopatología: definición, objetivos y manuales de clasificaciónMariangel StredelAún no hay calificaciones
- MentalismoDocumento3 páginasMentalismoGiancarlo López Velásquez100% (1)
- GEOMETRÍA-ÁNGULOS-TRIÁNGULOSDocumento15 páginasGEOMETRÍA-ÁNGULOS-TRIÁNGULOSEric BustamanteAún no hay calificaciones
- 1s1 Diodos y Rectificadores Apuntes Parte1Documento5 páginas1s1 Diodos y Rectificadores Apuntes Parte1goku saiyanAún no hay calificaciones
- Tensión NominalDocumento3 páginasTensión NominaljuanitoAún no hay calificaciones
- Capitulo 8 GerenciaDocumento3 páginasCapitulo 8 GerenciaAlejandra OlivaresAún no hay calificaciones
- Hilado de lana guarniciones flexibles y rígidasDocumento3 páginasHilado de lana guarniciones flexibles y rígidasArturo ParedesAún no hay calificaciones
- Estrategias marketing industria venadoDocumento96 páginasEstrategias marketing industria venadoTatti MendozaAún no hay calificaciones
- Lengua 3Documento4 páginasLengua 3mariajesus0921Aún no hay calificaciones
- Diagrama Del Proceso Del CompostDocumento1 páginaDiagrama Del Proceso Del CompostMARIA MUNERAAún no hay calificaciones
- 6to de Primaria Corazon (Cartilla)Documento16 páginas6to de Primaria Corazon (Cartilla)doris morandoAún no hay calificaciones
- Lainvestigacionetnografica 140903205458 Phpapp01Documento18 páginasLainvestigacionetnografica 140903205458 Phpapp01JazminPeñaAún no hay calificaciones
- Movimiento Feminista en ChileDocumento12 páginasMovimiento Feminista en ChileMonse Rocío100% (2)
- Convocatoria Familias ResponsablesDocumento3 páginasConvocatoria Familias ResponsablesTere VelasquezAún no hay calificaciones
- El Discurso de Graduación.Documento6 páginasEl Discurso de Graduación.Milagro Gracia0% (1)
- Corrientes Pedagogicas Contemporaneas 2022 Modulo 2Documento19 páginasCorrientes Pedagogicas Contemporaneas 2022 Modulo 2Mayra GuzmanAún no hay calificaciones
- Coronavirus peligro salud CorelDraw herramientasDocumento3 páginasCoronavirus peligro salud CorelDraw herramientasPablo MontesAún no hay calificaciones
- Muña o Inka Muña o Minthostachys MollisDocumento2 páginasMuña o Inka Muña o Minthostachys MollisKenyo Matencio AtaoAún no hay calificaciones
- Estrategias de Intervención PsicopedagógicasDocumento31 páginasEstrategias de Intervención PsicopedagógicasMaria Fatima PereiraAún no hay calificaciones
- Presentacion Esquematica de La Cadena de AbastecimientoDocumento29 páginasPresentacion Esquematica de La Cadena de AbastecimientoDeivid Leonardo Gallo PascagazaAún no hay calificaciones
- Cosmovision de Ruptura y ExperimentacionDocumento4 páginasCosmovision de Ruptura y Experimentacionalexia carjuzaaAún no hay calificaciones
- Preguntas sobre conceptos básicos de sistemas de informaciónDocumento56 páginasPreguntas sobre conceptos básicos de sistemas de informaciónBruno GuerreroAún no hay calificaciones
- Seis Pasos para Implementar Mantenimiento PlanificadoDocumento9 páginasSeis Pasos para Implementar Mantenimiento PlanificadoNarciso VasquezAún no hay calificaciones
- Programa Mensual de Reuniones de Inicio de GuardiaDocumento2 páginasPrograma Mensual de Reuniones de Inicio de GuardiaLuis Julio LimacheAún no hay calificaciones
- El libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)De EverandEl libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)Calificación: 3 de 5 estrellas3/5 (2)
- Terapia cognitiva: Conceptos básicos y profundizaciónDe EverandTerapia cognitiva: Conceptos básicos y profundizaciónCalificación: 5 de 5 estrellas5/5 (1)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- TDAH en Adultos. Cómo Reconocer y Tratar a un Adulto con TDAH en 30 Fáciles PasosDe EverandTDAH en Adultos. Cómo Reconocer y Tratar a un Adulto con TDAH en 30 Fáciles PasosCalificación: 4 de 5 estrellas4/5 (8)
- Sana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónDe EverandSana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónCalificación: 5 de 5 estrellas5/5 (4)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- Fisiopatología de las enfermedades cardiovascularesDe EverandFisiopatología de las enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- Dieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasDe EverandDieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasCalificación: 4 de 5 estrellas4/5 (9)
- Psicópatas seriales: Un recorrido por su oscura e inquietante naturalezaDe EverandPsicópatas seriales: Un recorrido por su oscura e inquietante naturalezaCalificación: 4 de 5 estrellas4/5 (3)
- Muchas Vidas, Muchos Sabios (Many Lives, Many Masters): (Many Lives, Many Masters)De EverandMuchas Vidas, Muchos Sabios (Many Lives, Many Masters): (Many Lives, Many Masters)Calificación: 4 de 5 estrellas4/5 (475)
- Prescripción de ejercico físico para la saludDe EverandPrescripción de ejercico físico para la saludCalificación: 5 de 5 estrellas5/5 (1)
- La Dieta Mediterránea Para Principiantes, Guía Paso A Paso Con Recetas Para Comer Mejor Y Bajar De PesoDe EverandLa Dieta Mediterránea Para Principiantes, Guía Paso A Paso Con Recetas Para Comer Mejor Y Bajar De PesoCalificación: 5 de 5 estrellas5/5 (2)
- Gua Sha: Guía de autotratamiento completoDe EverandGua Sha: Guía de autotratamiento completoCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Psicoterapia breve con niños y adolescentesDe EverandPsicoterapia breve con niños y adolescentesCalificación: 4.5 de 5 estrellas4.5/5 (15)
- Psiconeuroinmunología para la práctica clínicaDe EverandPsiconeuroinmunología para la práctica clínicaCalificación: 5 de 5 estrellas5/5 (4)
- Cardiología y enfermedades cardiovascularesDe EverandCardiología y enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- El arte del descanso: Descubre el método para dormir bien y descansar mejorDe EverandEl arte del descanso: Descubre el método para dormir bien y descansar mejorCalificación: 4.5 de 5 estrellas4.5/5 (4)
- Cómo curar la artritis: Curación natural de la artritis, la artrosis, la gota y la fibromialgiaDe EverandCómo curar la artritis: Curación natural de la artritis, la artrosis, la gota y la fibromialgiaCalificación: 5 de 5 estrellas5/5 (17)
- El daño cerebral invisible (3ª edición, revisada y actualizada): Alteraciones cognitivas en TCE, ictus y otras lesiones cerebralesDe EverandEl daño cerebral invisible (3ª edición, revisada y actualizada): Alteraciones cognitivas en TCE, ictus y otras lesiones cerebralesCalificación: 4.5 de 5 estrellas4.5/5 (9)
- Más Aristóteles y menos Concerta®: Las cuatro causas del TDAHDe EverandMás Aristóteles y menos Concerta®: Las cuatro causas del TDAHCalificación: 5 de 5 estrellas5/5 (2)
- ABC de los procedimientos médicos básicos: Una guía de aprendizaje y enseñanza para profesionales de medicinaDe EverandABC de los procedimientos médicos básicos: Una guía de aprendizaje y enseñanza para profesionales de medicinaCalificación: 4 de 5 estrellas4/5 (4)