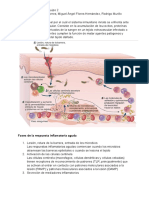
También podría gustarte
- Celulas Del Sistema InmuneDocumento14 páginasCelulas Del Sistema InmuneAntonio RodriguezAún no hay calificaciones
- Inflamación aguda y crónicaDocumento18 páginasInflamación aguda y crónicaYanina Pdv90% (10)
- PMN y MNDocumento8 páginasPMN y MNLili Moreta100% (1)
- NeutrofilosDocumento6 páginasNeutrofilosMeleMeleninAún no hay calificaciones
- Inflamación AgudaDocumento4 páginasInflamación AgudaOriana RegaladoAún no hay calificaciones
- 4.clase 2 - 9 Inmunidad Innata IIIDocumento15 páginas4.clase 2 - 9 Inmunidad Innata IIIElías Damián Martínez TerénAún no hay calificaciones
- Sistema InmunitarioDocumento11 páginasSistema InmunitarioJoaquin Biglia AvanciniAún no hay calificaciones
- Fagocitosis, proceso de ingestión de patógenosDocumento10 páginasFagocitosis, proceso de ingestión de patógenosJuliana AmayaAún no hay calificaciones
- Glóbulos BlancosDocumento9 páginasGlóbulos BlancosLisbet ZuhuAún no hay calificaciones
- fisiologia resistencia a la infecciónDocumento11 páginasfisiologia resistencia a la infecciónDani Slayer IriarteAún no hay calificaciones
- Inmunologia Células 1Documento13 páginasInmunologia Células 1Jaquii100% (1)
- Linfocitos B y TDocumento11 páginasLinfocitos B y TLourdes Tuanama JaraAún no hay calificaciones
- Sistema LeucocitarioDocumento12 páginasSistema LeucocitarioNoelia NocedaAún no hay calificaciones
- Capitulo 3 Robbins Inflamacion aguda_Documento15 páginasCapitulo 3 Robbins Inflamacion aguda_nadiaAún no hay calificaciones
- InflamaciónDocumento9 páginasInflamaciónIGNACIO SALVADOR FERNANDEZ GIRONAún no hay calificaciones
- 06 LeucocitosDocumento15 páginas06 LeucocitosKaren RamosAún no hay calificaciones
- Sistema InmunitarioDocumento25 páginasSistema InmunitarioCamila Quispe VilchezAún no hay calificaciones
- 1 Serie BlancaDocumento9 páginas1 Serie BlancaCarly Tarqui YujraAún no hay calificaciones
- Inmunologia Capitulo 2Documento6 páginasInmunologia Capitulo 2Daniela Medina PelaezAún no hay calificaciones
- Células y Moléculas Que Intervienen en La InflamacioónDocumento5 páginasCélulas y Moléculas Que Intervienen en La InflamacioónDenisse Morante RuizAún no hay calificaciones
- Tarea 3.1 - FagocitosisDocumento9 páginasTarea 3.1 - FagocitosisRosmery Sánchez TejadaAún no hay calificaciones
- Repaso tema 5^Documento6 páginasRepaso tema 5^orlynmagdalysAún no hay calificaciones
- LEUCOCITOSDocumento5 páginasLEUCOCITOSAnkelly CeballoAún no hay calificaciones
- Celulas Sistema InmuneDocumento5 páginasCelulas Sistema InmuneLízbeth RojasAún no hay calificaciones
- Leucocitos RossDocumento3 páginasLeucocitos RossBelen DuceAún no hay calificaciones
- Globulos BlancosDocumento45 páginasGlobulos BlancosHernán Padilla100% (2)
- Guia de InmunologiaDocumento4 páginasGuia de InmunologiaMarina OlmosAún no hay calificaciones
- Fisiologia LeucocitariaDocumento7 páginasFisiologia LeucocitariaJaviera Alejandra Montoya AcevedoAún no hay calificaciones
- RESISTENCIA DEL ORGANISMO A LA INFECCIÓN: LEUCOCITOS Y FAGOCITOSISDocumento46 páginasRESISTENCIA DEL ORGANISMO A LA INFECCIÓN: LEUCOCITOS Y FAGOCITOSISLizeth ManuAún no hay calificaciones
- Mecanismos Efectores de La Respuesta Inmune InnataDocumento7 páginasMecanismos Efectores de La Respuesta Inmune InnataEmilis MadridAún no hay calificaciones
- 1 . - Leucocitos y Resistencia Del Cuerpo A La Infección.Documento13 páginas1 . - Leucocitos y Resistencia Del Cuerpo A La Infección.deeppurple.478Aún no hay calificaciones
- 1 B . - Leucocitos y Resistencia Del Cuerpo A La InfecciónDocumento27 páginas1 B . - Leucocitos y Resistencia Del Cuerpo A La Infeccióndeeppurple.478Aún no hay calificaciones
- Serie Blanca IDocumento13 páginasSerie Blanca IBrenda ZalazarAún no hay calificaciones
- Componentes de La Inmunidad InnataDocumento33 páginasComponentes de La Inmunidad InnataPedro ReyesAún no hay calificaciones
- Células y Tejidos Del Sistema InmunitarioDocumento14 páginasCélulas y Tejidos Del Sistema InmunitarioGaby HenriquezAún no hay calificaciones
- EusinófilosDocumento4 páginasEusinófilosMirnaAún no hay calificaciones
- RespuestaInmunitariaInnata CarolineBarredoDocumento43 páginasRespuestaInmunitariaInnata CarolineBarredoCaroline BarredoAún no hay calificaciones
- Inflamación Aguda y CrónicaDocumento6 páginasInflamación Aguda y CrónicaFernando Villavicencio100% (1)
- Fagocitosis: definición y clasificaciónDocumento37 páginasFagocitosis: definición y clasificaciónJunior ValdiviezoAún no hay calificaciones
- Fisiologia de Los GranulositosDocumento10 páginasFisiologia de Los GranulositosDaniel De OcamposAún no hay calificaciones
- leucocitos fisiologíaDocumento38 páginasleucocitos fisiologíaJosé Luis RaymundoAún no hay calificaciones
- LEUCOCITOSDocumento6 páginasLEUCOCITOSBrayan BarajasAún no hay calificaciones
- Capitulo 34-SebastiansanchezDocumento2 páginasCapitulo 34-SebastiansanchezSebastian Sanchez MayaAún no hay calificaciones
- Anatomía y Fisiología Del Sistema InmuneDocumento39 páginasAnatomía y Fisiología Del Sistema InmuneCésar Morante ChavarryAún no hay calificaciones
- INMUNOLOGDocumento27 páginasINMUNOLOGortiz1081Aún no hay calificaciones
- LEUCOCITOSDocumento7 páginasLEUCOCITOSBelen DuceAún no hay calificaciones
- Resumen 2 (Introducción A La Inmunología Humana)Documento17 páginasResumen 2 (Introducción A La Inmunología Humana)Jacqueline Agustina ToledoAún no hay calificaciones
- Acontecimientos CelularesDocumento7 páginasAcontecimientos CelularesNico Blanco BarajasAún no hay calificaciones
- Comunicación CelularDocumento43 páginasComunicación CelularDiana AguilarAún no hay calificaciones
- Función de leucocitosDocumento31 páginasFunción de leucocitosRichard Gonzales DíazAún no hay calificaciones
- P13. Inmunidad. Miguel Vereau. 202120Documento57 páginasP13. Inmunidad. Miguel Vereau. 202120GESU PIERO LOPEZ MEJIAAún no hay calificaciones
- Sangre IIIDocumento29 páginasSangre IIIdavid esquivelAún no hay calificaciones
- Inflamacion y Funcion de Los Neutrofilos y MacrofagosDocumento9 páginasInflamacion y Funcion de Los Neutrofilos y MacrofagosGiovanniGuerreroAún no hay calificaciones
- Fagocitosis-proceso-defensaDocumento16 páginasFagocitosis-proceso-defensaJeisonAlejandroÁlvarezVergaraAún no hay calificaciones
- Resistencia Del Organismo A La InfeccionDocumento55 páginasResistencia Del Organismo A La InfeccionIsadora MirandaAún no hay calificaciones
- Universidad Veracruzana: Inflamación Y ReparaciónDocumento88 páginasUniversidad Veracruzana: Inflamación Y ReparaciónMireya MeseguerAún no hay calificaciones
- 1 Resumen de Neutrã FilosDocumento4 páginas1 Resumen de Neutrã FilosJhanKarloFloresFlores100% (1)
- Uniones y Señalizacion CelularDocumento8 páginasUniones y Señalizacion CelularmfvelascoAún no hay calificaciones
- Metabolismo de Los Leucocitos 1Documento14 páginasMetabolismo de Los Leucocitos 1Silhhean CruzAún no hay calificaciones
- Anemia Para Todos: Medicina Para TodosDe EverandAnemia Para Todos: Medicina Para TodosCalificación: 3 de 5 estrellas3/5 (1)
- Practico 3 EspecialesDocumento13 páginasPractico 3 EspecialesGonzales Pereira YennyAún no hay calificaciones
- 1 Ejercicio 1E A Abandono Formulac. Asientos 17.12.20.Documento1 página1 Ejercicio 1E A Abandono Formulac. Asientos 17.12.20.Gonzales Pereira YennyAún no hay calificaciones
- Ley 843 CompendioDocumento536 páginasLey 843 CompendioMiguel Chura CordovaAún no hay calificaciones
- Formato Nuevo de PsicotropicosDocumento16 páginasFormato Nuevo de PsicotropicosGonzales Pereira YennyAún no hay calificaciones
- Xi Nervio AccesorioDocumento14 páginasXi Nervio AccesorioGonzales Pereira YennyAún no hay calificaciones
- Procesos Cognitivos SesiònDocumento65 páginasProcesos Cognitivos SesiònCésar DelgadoAún no hay calificaciones
- Concepción Del Carmen Ahuja AguirreDocumento119 páginasConcepción Del Carmen Ahuja AguirreWillman Ramirez PomaAún no hay calificaciones
- Electrocardiograma FinalDocumento23 páginasElectrocardiograma FinalLaura SanabriaAún no hay calificaciones
- Palmer - Et - Al-2015 ICSH Recommendations For The Standardization of Nomenclature - En.esDocumento17 páginasPalmer - Et - Al-2015 ICSH Recommendations For The Standardization of Nomenclature - En.esAngeles Toala100% (1)
- Psicologia Ambiental Leticia2Documento13 páginasPsicologia Ambiental Leticia2Dyogo Arturo Mendez TrilloAún no hay calificaciones
- Morfologia de Insectos - Abdómen y ApéndicesDocumento10 páginasMorfologia de Insectos - Abdómen y ApéndicesKatherine CardenasAún no hay calificaciones
- Fisiología Sensitiva y Motora Del Aparato EstomatognáticoDocumento10 páginasFisiología Sensitiva y Motora Del Aparato Estomatognáticoimane imaneAún no hay calificaciones
- Factores Que Influyen Sobre El Proceso de FotosintesisDocumento18 páginasFactores Que Influyen Sobre El Proceso de FotosintesisANGEL DENILSON MONTOYA DAVILAAún no hay calificaciones
- Precios TerapiaDocumento1 páginaPrecios TerapiaPatricia MinickAún no hay calificaciones
- Semiologia Neurologica. SeminarioDocumento34 páginasSemiologia Neurologica. SeminariojesusAún no hay calificaciones
- ANATOMÍA DE LA LARINGE FMC DianaDocumento47 páginasANATOMÍA DE LA LARINGE FMC DianaJuliana Cano GómezAún no hay calificaciones
- Cuestionario BiologiaDocumento2 páginasCuestionario BiologiaLucho AldasAún no hay calificaciones
- Juan Alberto Peña-Infografia Macromoleculas PDFDocumento1 páginaJuan Alberto Peña-Infografia Macromoleculas PDFjuanAún no hay calificaciones
- Resultado LaboratorioDocumento3 páginasResultado LaboratorioDavid BritoAún no hay calificaciones
- Clasificación de proteínas en holoproteínas y heteroproteínasDocumento2 páginasClasificación de proteínas en holoproteínas y heteroproteínasJuan diego Zabala navarroAún no hay calificaciones
- Cuestionario 4Documento3 páginasCuestionario 4JORDY ANGELO SALVADOR SALAZARAún no hay calificaciones
- Anfibios: características, órdenes y ciclo de vidaDocumento5 páginasAnfibios: características, órdenes y ciclo de vidaMacarena HonekerAún no hay calificaciones
- Solucionario Canal 1 Examen Cepu 2022-IDocumento10 páginasSolucionario Canal 1 Examen Cepu 2022-IPablo Ramos100% (2)
- Embriogenésis Del Sistema Nervioso HumanoDocumento2 páginasEmbriogenésis Del Sistema Nervioso HumanoLOPEZ GUERRA Christopher GabrielAún no hay calificaciones
- TEC y Traumatismo Vertebro Medular: Cuidados de Enfermería y DiagnósticosDocumento38 páginasTEC y Traumatismo Vertebro Medular: Cuidados de Enfermería y DiagnósticosMichael Jefri VillarAún no hay calificaciones
- La CelulaDocumento3 páginasLa CelulaCristian LlanosAún no hay calificaciones
- Huesos Del TroncoDocumento15 páginasHuesos Del TroncoXaxelGamplaysXAún no hay calificaciones
- Aqui Hay Plomo - Keith LugerDocumento65 páginasAqui Hay Plomo - Keith LugerCenobioAún no hay calificaciones
- UNIDAD VIII-SistemaRespiratorioDocumento25 páginasUNIDAD VIII-SistemaRespiratorioVictor CalvilloAún no hay calificaciones
- Cartílago y hueso: tipos, estructura y funciónDocumento4 páginasCartílago y hueso: tipos, estructura y funciónArlyn DiazAún no hay calificaciones
- Fichas NerviosDocumento7 páginasFichas NerviosFernanda CentellasAún no hay calificaciones
- Aparato EndocrinoDocumento1 páginaAparato EndocrinoDanis VaccaAún no hay calificaciones
- Hta en La Insuficiencia RenalDocumento6 páginasHta en La Insuficiencia RenalYovera Vivanco AbrahamAún no hay calificaciones
- Ele UveDocumento5 páginasEle Uvejuan felipeAún no hay calificaciones
- Ejemplos de FlavonoidesDocumento106 páginasEjemplos de FlavonoidesMARLENY MARIBEL QUISPE CAHUANAAún no hay calificaciones