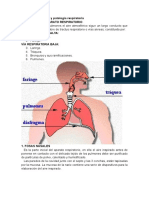
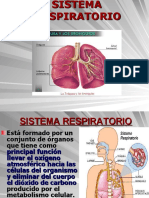
También podría gustarte
- Conceptos Y Estrategias En Terapia RespiratoriaDe EverandConceptos Y Estrategias En Terapia RespiratoriaCalificación: 4 de 5 estrellas4/5 (1)
- 2005, RCR, RX Torax UCIDocumento11 páginas2005, RCR, RX Torax UCIAngela TorresAún no hay calificaciones
- Tema 3 Aparato Respiratorio y FonadorDocumento10 páginasTema 3 Aparato Respiratorio y FonadorNatalia Grimaldi SánchezAún no hay calificaciones
- Intro Ducci OnDocumento7 páginasIntro Ducci OnMiguel Angel BaezAún no hay calificaciones
- Sistema Respiratorio ApuntesDocumento4 páginasSistema Respiratorio ApuntesJuan Manuel GuerreroAún no hay calificaciones
- Sistema RespiratorioDocumento5 páginasSistema RespiratorioLuz Marina Lozada RodriguezAún no hay calificaciones
- Bitacora de Viaje3Documento77 páginasBitacora de Viaje3LuluAún no hay calificaciones
- Sistema Respiratotio - Eduardo MariñosDocumento21 páginasSistema Respiratotio - Eduardo Mariñoscaleb huamanAún no hay calificaciones
- Sistema R, Exc, EndDocumento58 páginasSistema R, Exc, EndLeonel Huanacuni CotradoAún no hay calificaciones
- Biologia 3ro de SecundariaDocumento28 páginasBiologia 3ro de SecundariaShirley VanessaAún no hay calificaciones
- Sistema RespiratorioDocumento17 páginasSistema Respiratoriolomther111Aún no hay calificaciones
- Anatomía y Fisiologia Del Sistema RespiratorioDocumento4 páginasAnatomía y Fisiologia Del Sistema RespiratorioyajairaAún no hay calificaciones
- Morfología Del Sistema RespiratorioDocumento9 páginasMorfología Del Sistema RespiratorioAle Gómez LinaresAún no hay calificaciones
- Aparato RespiratorioDocumento18 páginasAparato RespiratorioMalhi Mariana Rios Huarcaya0% (1)
- Aparato RespiratorioDocumento23 páginasAparato RespiratorioYomira YulissaAún no hay calificaciones
- Fisiologia Sistema RespiratorioDocumento11 páginasFisiologia Sistema RespiratorioRichard PerezAún no hay calificaciones
- InformeDocumento14 páginasInformeGustavo Tabo MamaniAún no hay calificaciones
- Teoria RespiratorioDocumento10 páginasTeoria RespiratorioAnthony H-mAún no hay calificaciones
- Informe de Laboratorio RespiratorioDocumento8 páginasInforme de Laboratorio RespiratorioDANIELA ALEJANDRA RAMOS CASTAÑEDAAún no hay calificaciones
- Vias PDFDocumento9 páginasVias PDFPamela GavinoAún no hay calificaciones
- Vias Respiratorias Superiores e InferioresDocumento5 páginasVias Respiratorias Superiores e InferioresDannyel Flores Lott50% (2)
- Anatomia y Fisiologia DocumentoDocumento20 páginasAnatomia y Fisiologia Documentojesus guerraAún no hay calificaciones
- Informe Del Sistema RespiratorioDocumento7 páginasInforme Del Sistema RespiratorioWil SmithAún no hay calificaciones
- Anatomía y Fisiologia RespiratoriaDocumento23 páginasAnatomía y Fisiologia RespiratoriaScott McgeeAún no hay calificaciones
- Sistema RepiratorioDocumento14 páginasSistema Repiratoriolilianamonica100% (1)
- MORFOFISIOLOGIA Tarea 4 - Armonización CorporalDocumento10 páginasMORFOFISIOLOGIA Tarea 4 - Armonización CorporalAndres GutierrezAún no hay calificaciones
- Sistema Respiratorio 479074 Downloadable 1072077Documento5 páginasSistema Respiratorio 479074 Downloadable 1072077Jose OllarbeAún no hay calificaciones
- Informe PulmonDocumento8 páginasInforme PulmonLaurita Aspi50% (4)
- Practica de Disección de PulmónDocumento9 páginasPractica de Disección de PulmónAdri PanduroAún no hay calificaciones
- Definición Del Sistema RespiratorioDocumento11 páginasDefinición Del Sistema RespiratorioMannu ReysAún no hay calificaciones
- Anatomía y Fisiología PulmonarDocumento13 páginasAnatomía y Fisiología PulmonarSonia chubAún no hay calificaciones
- Presentacion Sistema Respiratorio Morfofisiologia Sin FotosDocumento26 páginasPresentacion Sistema Respiratorio Morfofisiologia Sin FotosJose David Casillas MuñozAún no hay calificaciones
- Aparato RespiratoriooooDocumento14 páginasAparato RespiratoriooooDaneska PeñaAún no hay calificaciones
- Aparato Respiratorio y FonadorDocumento6 páginasAparato Respiratorio y FonadorMaria Y Sus Manos McvAún no hay calificaciones
- Anatomia Y Fisiologia RespiratoriaDocumento52 páginasAnatomia Y Fisiologia Respiratoriacarolina DAún no hay calificaciones
- El Sistema RespiratorioDocumento8 páginasEl Sistema RespiratorioEnos DEGBEAún no hay calificaciones
- Anatomía TareaDocumento16 páginasAnatomía TareaGaby Puc TecAún no hay calificaciones
- Aparato RespiratorioDocumento10 páginasAparato RespiratorioRivera GabrielaAún no hay calificaciones
- Istema Espiratorio: Modul ODocumento13 páginasIstema Espiratorio: Modul Ochristian LopezAún no hay calificaciones
- Unidad 4. Aparato RespiratorioDocumento6 páginasUnidad 4. Aparato RespiratoriokarmaAún no hay calificaciones
- Tema 2 Aparato RespiratorioDocumento4 páginasTema 2 Aparato RespiratorioivanAún no hay calificaciones
- Tema 2 Aparato RespiratorioDocumento4 páginasTema 2 Aparato RespiratorioivanAún no hay calificaciones
- Aparato Respiratorio de Los AnimalesDocumento6 páginasAparato Respiratorio de Los AnimalesElizabeth MachucaAún no hay calificaciones
- 46 FISIO Clase Nº9 SIST - RESPIRATORIODocumento43 páginas46 FISIO Clase Nº9 SIST - RESPIRATORIOMario PeñalozaAún no hay calificaciones
- Morfofisiologia Tarea 4 - Armonización Corporal - Repiratorio - MillerDocumento11 páginasMorfofisiologia Tarea 4 - Armonización Corporal - Repiratorio - MillerAndres GutierrezAún no hay calificaciones
- Fisio 1 - Unidad 3 - LecturaDocumento9 páginasFisio 1 - Unidad 3 - LecturaFelicia RebolloAún no hay calificaciones
- Anatomía Tema 2 - El Aparato RespiratorioDocumento6 páginasAnatomía Tema 2 - El Aparato RespiratorioCristina Benítez SánchezAún no hay calificaciones
- Hoja Tecnica Del Sistema RespiratorioDocumento3 páginasHoja Tecnica Del Sistema RespiratorioFabián Alejandro Lezama LeonAún no hay calificaciones
- InvestigacionDocumento18 páginasInvestigacionAriana TzintzireAún no hay calificaciones
- 1 - Aparato RespiratorioDocumento8 páginas1 - Aparato RespiratorioLuciana BasseAún no hay calificaciones
- Anatomía. Fisiologia y Fsiopatologia de Los PulmonesDocumento31 páginasAnatomía. Fisiologia y Fsiopatologia de Los PulmonesMargarita TorresAún no hay calificaciones
- Fisiología Módulo 4Documento8 páginasFisiología Módulo 4Santiago RodriguezAún no hay calificaciones
- Aparato RespiratorioDocumento14 páginasAparato Respiratorio72938214Aún no hay calificaciones
- Aparato RespiratorioDocumento7 páginasAparato RespiratorioFranco FurnariAún no hay calificaciones
- 1-0 Recuerdo Anatomofisiologco Ap Respiratorio CampusDocumento18 páginas1-0 Recuerdo Anatomofisiologco Ap Respiratorio CampusBelén SilvaAún no hay calificaciones
- Sistema RespiratorioDocumento21 páginasSistema Respiratorionico.cristalliAún no hay calificaciones
- Anatomía y Fisiología RespiratoriaDocumento19 páginasAnatomía y Fisiología RespiratoriaMARYVON100% (1)
- Función de Las Estructuras Del Sistema RespiratorioDocumento7 páginasFunción de Las Estructuras Del Sistema RespiratorioJorge AquinoAún no hay calificaciones
- Aparato RESPIRATORIODocumento4 páginasAparato RESPIRATORIOInfocentroCharapotoAún no hay calificaciones
- Resúmen Sobre La Anatomia Cardio RespiratoriaDocumento28 páginasResúmen Sobre La Anatomia Cardio Respiratoriadanielaroque335Aún no hay calificaciones
- Sistema Respiratorio para Segundo Grado de SecundariaDocumento6 páginasSistema Respiratorio para Segundo Grado de SecundariaFernando Panta MoranAún no hay calificaciones
- Normas Oficiales Mexicanas para Instalaciones HidraulicasDocumento2 páginasNormas Oficiales Mexicanas para Instalaciones HidraulicasVictor CalvilloAún no hay calificaciones
- Programa Operativo Anual 2022Documento3 páginasPrograma Operativo Anual 2022Victor CalvilloAún no hay calificaciones
- Reporte de Práctica # 1Documento3 páginasReporte de Práctica # 1Victor CalvilloAún no hay calificaciones
- Maquinaria ExpoDocumento74 páginasMaquinaria ExpoVictor CalvilloAún no hay calificaciones
- Mecanismo de La RespiracionDocumento1 páginaMecanismo de La RespiracionVictor CalvilloAún no hay calificaciones
- Unidad 8Documento1 páginaUnidad 8Victor CalvilloAún no hay calificaciones
- Tarea U2 PDFDocumento1 páginaTarea U2 PDFVictor CalvilloAún no hay calificaciones
- Corte Directo PDFDocumento2 páginasCorte Directo PDFVictor CalvilloAún no hay calificaciones
- Unidad 4 Escenario EconomicoDocumento35 páginasUnidad 4 Escenario EconomicoVictor CalvilloAún no hay calificaciones
- Trabajo FinalDocumento17 páginasTrabajo FinalVictor CalvilloAún no hay calificaciones
- ExpoDocumento54 páginasExpoVictor CalvilloAún no hay calificaciones
- MAQUINARIA PARA EXCAVACION Eq#2Documento73 páginasMAQUINARIA PARA EXCAVACION Eq#2Victor CalvilloAún no hay calificaciones
- Unidad 3 RendimientoDocumento15 páginasUnidad 3 RendimientoVictor Calvillo100% (1)
- ComprobacionesDocumento8 páginasComprobacionesVictor CalvilloAún no hay calificaciones
- Galván Cervantes - Trabajo Individual MDI PDFDocumento15 páginasGalván Cervantes - Trabajo Individual MDI PDFVictor CalvilloAún no hay calificaciones
- Volumen 5 Tomo Ii Instalacion Hidraulica 2022Documento21 páginasVolumen 5 Tomo Ii Instalacion Hidraulica 2022Victor CalvilloAún no hay calificaciones
- Practica 11 PDFDocumento14 páginasPractica 11 PDFVictor CalvilloAún no hay calificaciones
- Volumen 5 Tomo Ii Instalacion Hidraulica 2022-13-19Documento7 páginasVolumen 5 Tomo Ii Instalacion Hidraulica 2022-13-19Victor CalvilloAún no hay calificaciones
- Estabilización de Suelos Con CementoDocumento4 páginasEstabilización de Suelos Con CementoVictor CalvilloAún no hay calificaciones
- Instalacion de Agua PotableDocumento10 páginasInstalacion de Agua PotableVictor CalvilloAún no hay calificaciones
- Descripción General de Los PiojosDocumento5 páginasDescripción General de Los PiojosJhancarla Nina GalarzaAún no hay calificaciones
- Circulacion Pulmonar, Edema Pulmonar, Liquido Pleural Cap 38Documento22 páginasCirculacion Pulmonar, Edema Pulmonar, Liquido Pleural Cap 38Rosângela PaixãoAún no hay calificaciones
- Conductos Alveolares Sacos AlveolaresDocumento9 páginasConductos Alveolares Sacos AlveolaresRuthsita MoretaAún no hay calificaciones
- ADAPTACIÓNDocumento4 páginasADAPTACIÓNNatalia Laguado RestrepoAún no hay calificaciones
- Apuntes Sistema CardiovascularDocumento6 páginasApuntes Sistema CardiovascularBritney OñateAún no hay calificaciones
- Sistema de Investigacion Venas y OdontologicoDocumento20 páginasSistema de Investigacion Venas y Odontologicomelissa lopezAún no hay calificaciones
- Paquete de Atencion Escolar MaisDocumento15 páginasPaquete de Atencion Escolar MaisJubencio Anticona CuevaAún no hay calificaciones
- Aparato CirculatorioDocumento16 páginasAparato CirculatorioPaula Andrey CESPEDES DIAZAún no hay calificaciones
- MicrocirculacionDocumento32 páginasMicrocirculacionmeli mur100% (1)
- Aparato CirculatorioDocumento3 páginasAparato CirculatorioMiriam Méndez BeasterAún no hay calificaciones
- Resumen Capitulo 41 Transporte O2 y Co2Documento5 páginasResumen Capitulo 41 Transporte O2 y Co2Lii LiiAún no hay calificaciones
- Artículo TraducidoDocumento13 páginasArtículo Traducidovargas070603Aún no hay calificaciones
- Difusion de GasesDocumento19 páginasDifusion de GasesDaniela Bedoya Pérez100% (1)
- Capacitación Mc. Airam by VitalherDocumento20 páginasCapacitación Mc. Airam by VitalherOscar Fortech100% (1)
- Circulación Sanguínea, Arterias, Venas y LinfáticosDocumento21 páginasCirculación Sanguínea, Arterias, Venas y LinfáticosAlejandro Villegas AmadorAún no hay calificaciones
- Tratamiento de Aguas - Sistema Actual y Nuevas SolucionesDocumento68 páginasTratamiento de Aguas - Sistema Actual y Nuevas SolucionesjucascribAún no hay calificaciones
- MicocirculaciónDocumento6 páginasMicocirculaciónAnonymous alzR2b1pqAún no hay calificaciones
- Punto de Fusion PracticaDocumento10 páginasPunto de Fusion PracticaJeanniel CamposAún no hay calificaciones
- Fisiologia - Respiratoria - III TRANSPORTE DE GASESDocumento29 páginasFisiologia - Respiratoria - III TRANSPORTE DE GASEScristian duqueAún no hay calificaciones
- Práctica Sistema RespiratorioDocumento4 páginasPráctica Sistema RespiratorioHender Iker Llauce ChapoñanAún no hay calificaciones
- Fisiología v1.0Documento43 páginasFisiología v1.0DIANA MARCELA ERAZO LASSOAún no hay calificaciones
- Cuidado Del Sistema Nervioso y Sistema Inmune y Los SentidosDocumento89 páginasCuidado Del Sistema Nervioso y Sistema Inmune y Los SentidosJeisson CarvajalAún no hay calificaciones
- Gema - Canales - 20191900150 2Documento7 páginasGema - Canales - 20191900150 2Gema CanalesAún no hay calificaciones
- Xabc de TraumaDocumento20 páginasXabc de TraumaPao PérezAún no hay calificaciones
- Transferencia de Masa en Aplicaciones BiomedicasDocumento9 páginasTransferencia de Masa en Aplicaciones BiomedicasMarco Estuardo Solórzano HernándezAún no hay calificaciones
- UNIDAD 4.FarmacocineÌ - Tica - DistribucioÌ - NDocumento43 páginasUNIDAD 4.FarmacocineÌ - Tica - DistribucioÌ - NJuan Angel Varela BazánAún no hay calificaciones
- Clase 1. Introduccion A Cardiologia PDFDocumento12 páginasClase 1. Introduccion A Cardiologia PDForianacgoAún no hay calificaciones
- HistologiaDocumento35 páginasHistologiaCarla PortuguezAún no hay calificaciones
- MicrohematocritoDocumento6 páginasMicrohematocritoOk AmisitaAún no hay calificaciones