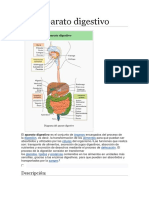
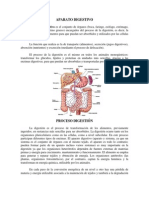
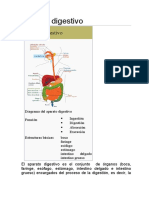
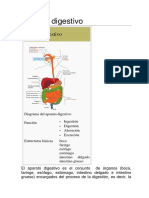
También podría gustarte
- Glándulas AnexasDocumento8 páginasGlándulas AnexasMartha DominguezAún no hay calificaciones
- Aparato DigestivoDocumento19 páginasAparato Digestivokareli FloresAún no hay calificaciones
- Sistema DigestivoDocumento4 páginasSistema Digestivocopiascarranza310Aún no hay calificaciones
- Sistema DigestivoDocumento14 páginasSistema DigestivoDaniela Fernanda Ochoa SanchezAún no hay calificaciones
- El Aparato DigestivoDocumento72 páginasEl Aparato DigestivoMore ParedesAún no hay calificaciones
- Aparato DigestivoDocumento16 páginasAparato DigestivoGabriela Diaz LaureanoAún no hay calificaciones
- Sistema Digestivo MonogástricosDocumento7 páginasSistema Digestivo MonogástricosFlavio CastañedaAún no hay calificaciones
- Plantilla para Artículos Normas APA Normasapa - Com 1Documento18 páginasPlantilla para Artículos Normas APA Normasapa - Com 1Vanesa NaranjoAún no hay calificaciones
- 7-Sistemas de Nutrición-Sistema DigestivoDocumento20 páginas7-Sistemas de Nutrición-Sistema DigestivoLautaro SolisAún no hay calificaciones
- Hemorragia DigestivaDocumento98 páginasHemorragia DigestivaEskarleth CoronelAún no hay calificaciones
- Aparato Digestivo-2021Documento11 páginasAparato Digestivo-2021gonzalo guachaminAún no hay calificaciones
- Protocolo de Fisiolog DigestivaDocumento24 páginasProtocolo de Fisiolog DigestivaRuth Esther Nuñez MaldonadoAún no hay calificaciones
- Apuntes Sistema Digestivo OKDocumento21 páginasApuntes Sistema Digestivo OKFernando LópezAún no hay calificaciones
- Aparato DigestivoDocumento4 páginasAparato Digestivohhjf7pww2yAún no hay calificaciones
- Aparato Digestivo - ResumenDocumento24 páginasAparato Digestivo - ResumenAdrienne LMAún no hay calificaciones
- A, H y F Del Tubo DigestivoDocumento22 páginasA, H y F Del Tubo DigestivoZora CaballeroAún no hay calificaciones
- Histologia Del Aparato DigestivoDocumento68 páginasHistologia Del Aparato DigestivoManuela Sarmiento GuarinAún no hay calificaciones
- Aparato DigestivoDocumento40 páginasAparato DigestivorocioAún no hay calificaciones
- Aparato DigestivoDocumento21 páginasAparato DigestivoHenry MistAún no hay calificaciones
- Aparato Digestivo Brunelli Fisiologia NutricionDocumento77 páginasAparato Digestivo Brunelli Fisiologia NutricionFiorela NavarroAún no hay calificaciones
- Sistema Digestivo II Organizacion General Del Tubo Digestivo y EsofagoDocumento5 páginasSistema Digestivo II Organizacion General Del Tubo Digestivo y EsofagoLiced Enriquez ChalcoAún no hay calificaciones
- Sistema DigestivoDocumento13 páginasSistema DigestivoAdolfo ArizaAún no hay calificaciones
- El Aparato DigestivoDocumento18 páginasEl Aparato DigestivoavalonchildAún no hay calificaciones
- Aparato DigestivoDocumento6 páginasAparato Digestivolauradani0% (1)
- GeriatríaDocumento34 páginasGeriatríaJime GuerreroAún no hay calificaciones
- Generalidades Del Sistema Digestivo AnatómicaDocumento7 páginasGeneralidades Del Sistema Digestivo Anatómicaagustinalguerrero2022Aún no hay calificaciones
- Aparato DigestivoDocumento13 páginasAparato DigestivoJaime PachecoAún no hay calificaciones
- Resumen Digestivo 2Documento15 páginasResumen Digestivo 2fernandaAún no hay calificaciones
- Sistema DigestivoDocumento9 páginasSistema DigestivoRocio AndiaAún no hay calificaciones
- Ores de La Bomba de Protones+GastroDocumento11 páginasOres de La Bomba de Protones+GastroGabriela GranadosAún no hay calificaciones
- Aparato Digestivo y RespiratorioDocumento30 páginasAparato Digestivo y RespiratorioWendy MartinezAún no hay calificaciones
- FinalDocumento19 páginasFinalValentin MartinezAún no hay calificaciones
- Sistema Digestivo Material de LecturaDocumento23 páginasSistema Digestivo Material de Lecturacristhian AyalaAún no hay calificaciones
- Aparato DigestivoDocumento13 páginasAparato DigestivoJaime PachecoAún no hay calificaciones
- Sistema Gastrointestinal - Digestivo - MorfofisiologíaDocumento31 páginasSistema Gastrointestinal - Digestivo - MorfofisiologíaVal NullAún no hay calificaciones
- Aparato DigestivoDocumento30 páginasAparato DigestivoEstefani DávilaAún no hay calificaciones
- Biología Tema 6 CoraDocumento27 páginasBiología Tema 6 Corakatalina Gutiérrez JimenezAún no hay calificaciones
- Sistema Digestivo Curso TephDocumento68 páginasSistema Digestivo Curso TephabelysAún no hay calificaciones
- 2.3 Aparato DogestivoDocumento6 páginas2.3 Aparato DogestivoCristhel RiosAún no hay calificaciones
- Aparato Digestivo KiwiDocumento7 páginasAparato Digestivo KiwiJACKPGALINDOAún no hay calificaciones
- Apuntes de Fisiologia DigestivaDocumento45 páginasApuntes de Fisiologia DigestivaRonald Mamani Huañapaco50% (4)
- Sistema Digestivo. Anatomía y FisiologíaDocumento40 páginasSistema Digestivo. Anatomía y FisiologíaJesica MachadoAún no hay calificaciones
- El Aparato Digestivo Es El Conjunto de ÓrganosDocumento3 páginasEl Aparato Digestivo Es El Conjunto de ÓrganosMayamayaayAún no hay calificaciones
- Tubo Digestivo HistologiaDocumento27 páginasTubo Digestivo HistologiaLaraBeatrizMoutinhoSantosAún no hay calificaciones
- Sistema Digestivo PDFDocumento11 páginasSistema Digestivo PDFRamiro MadriagaAún no hay calificaciones
- Anatomía Digestivo.Documento49 páginasAnatomía Digestivo.Carolina GomezAún no hay calificaciones
- Vaciamiento Gastrico y Absorción IntestinalDocumento24 páginasVaciamiento Gastrico y Absorción IntestinalAndy CarranzaAún no hay calificaciones
- Aparato DigestivoDocumento2 páginasAparato DigestivoCarla Xiomara Garcia ChacaliazaAún no hay calificaciones
- Tema 7 Anatomia Del Aparato DigestivoDocumento15 páginasTema 7 Anatomia Del Aparato DigestivoYodyAún no hay calificaciones
- Sistema DigestivoDocumento20 páginasSistema Digestivooguich6Aún no hay calificaciones
- Seminario 54Documento50 páginasSeminario 54tareasontimeyaAún no hay calificaciones
- Sistema DigestivoDocumento4 páginasSistema Digestivoescobarmabel.epsgrauAún no hay calificaciones
- Morfologia Del Tubo DigestivoDocumento14 páginasMorfologia Del Tubo DigestivoSan Sandry100% (1)
- Pag 32 Folleto de HistologiaDocumento6 páginasPag 32 Folleto de Histologiajennifer cuadradoAún no hay calificaciones
- DIGESTIVO AlDocumento10 páginasDIGESTIVO AlDIBL 96Aún no hay calificaciones
- Tema 2Documento95 páginasTema 2isabellacoronel2113Aún no hay calificaciones
- Cuestionario Sistema Digestivo HistoDocumento4 páginasCuestionario Sistema Digestivo Histoesmeralda vargasmonroyAún no hay calificaciones
- Motilidad GastroIntestinal IDocumento6 páginasMotilidad GastroIntestinal IcopiastopoAún no hay calificaciones
- Digestivo Parte IIDocumento8 páginasDigestivo Parte IIErika VertuanAún no hay calificaciones
- Aparato DigestivoDocumento8 páginasAparato DigestivoyorgelisAún no hay calificaciones
- Autorregulacion TermicaDocumento4 páginasAutorregulacion Termicavero_aga100% (1)
- ¿Que Es La DiabetesDocumento81 páginas¿Que Es La Diabetesteresa lourdes ramos yatacoAún no hay calificaciones
- Semana 13Documento11 páginasSemana 13josmet josokishy atalaya fajardoAún no hay calificaciones
- Fisiologia Endocrina:: Regulación de La GlicemiaDocumento38 páginasFisiologia Endocrina:: Regulación de La GlicemiafabianAún no hay calificaciones
- Fibrosis QuísticaDocumento2 páginasFibrosis QuísticaJeanneth JamiAún no hay calificaciones
- Guía de Sistema Digestivo NAOMI SALTOSDocumento16 páginasGuía de Sistema Digestivo NAOMI SALTOSNAOMI REBECA SALTOS CANDELAAún no hay calificaciones
- Marco ConceptualDocumento8 páginasMarco ConceptualEvi BravAún no hay calificaciones
- Pancreatitis Aguda UCIQDocumento19 páginasPancreatitis Aguda UCIQLuis Mosqueira MuñozAún no hay calificaciones
- Cistogastrostomia LaparoscopicaDocumento20 páginasCistogastrostomia LaparoscopicaEdu Pastten CarrascoAún no hay calificaciones
- Dosaje de La Amilasa Serica y Urinaria Pumacayo Perez BrighithDocumento24 páginasDosaje de La Amilasa Serica y Urinaria Pumacayo Perez BrighithBrighith PerezAún no hay calificaciones
- 10Documento4 páginas10LIZETHAún no hay calificaciones
- Chacras y ArcángelesDocumento8 páginasChacras y ArcángelesLoucía Ilamacuauhtli100% (1)
- Fármacos CardiotónicosDocumento7 páginasFármacos CardiotónicosDaniel Figueroa0% (1)
- Gen CFTR y Como Ocasiona La Fibrosis QuísticaDocumento31 páginasGen CFTR y Como Ocasiona La Fibrosis QuísticaADRIANA CRISTINA VILCHEZ ARISPEAún no hay calificaciones
- Sistema Digestivo: PerrosDocumento11 páginasSistema Digestivo: PerrosDylan Henriquez IturrietaAún no hay calificaciones
- Secreción Gástrica y PancreáticaDocumento9 páginasSecreción Gástrica y PancreáticaBelen BerruttiAún no hay calificaciones
- Documento Sin TítuloDocumento20 páginasDocumento Sin TítuloMarvin ChavezAún no hay calificaciones
- Cap 15 EmbrioDocumento14 páginasCap 15 EmbrioBELINDA XCARET RIVERA GONZALEZAún no hay calificaciones
- Práctica de LipasaDocumento2 páginasPráctica de LipasaSamantha Delgado0% (1)
- Resumen de Sistema EndocrinoDocumento7 páginasResumen de Sistema EndocrinoRoland M ReyesAún no hay calificaciones
- Preguntas Sobre Sistema Endocrino Bio 1Documento6 páginasPreguntas Sobre Sistema Endocrino Bio 1pipoAún no hay calificaciones
- Preguntas Amilasa - Grupo 4Documento2 páginasPreguntas Amilasa - Grupo 4Mariana LassoAún no hay calificaciones
- Sistema Digestivo GlandularDocumento8 páginasSistema Digestivo GlandularJorgelis CuevasAún no hay calificaciones
- Cuestionario Ahu Intestinos Bazo Pancreas GIULIANADocumento7 páginasCuestionario Ahu Intestinos Bazo Pancreas GIULIANAGIULIANA ELENA TICONA PARIAún no hay calificaciones
- Manual de Laboratorio Bio - 102 2012Documento36 páginasManual de Laboratorio Bio - 102 2012Oriol PolancoAún no hay calificaciones
- Cáncer de Páncreas - Síntomas y Causas - Mayo ClinicDocumento1 páginaCáncer de Páncreas - Síntomas y Causas - Mayo ClinicnoeliaAún no hay calificaciones
- DigestiDocumento6 páginasDigestiRoCo Brawn'sAún no hay calificaciones
- Criterios de AtlantaDocumento25 páginasCriterios de AtlantaMalú Ramos100% (2)
- Absorcion de NutrientesDocumento12 páginasAbsorcion de NutrientesSandy Gonzalez100% (1)