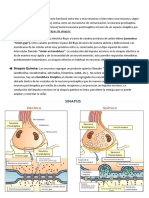
También podría gustarte
- Practica # 2Documento4 páginasPractica # 2Sonia C. Ojeda0% (1)
- Demostracion de Cuerpos Cetonicos en La OrinaDocumento21 páginasDemostracion de Cuerpos Cetonicos en La OrinaYiselda LisetheAún no hay calificaciones
- Actividad en Clase de Cuerpos CetonicosDocumento5 páginasActividad en Clase de Cuerpos CetonicosEzequiel SalazarAún no hay calificaciones
- Unidad 1 6to Medicina-2020Documento6 páginasUnidad 1 6to Medicina-2020Kalendra Miniaturas50% (2)
- Infográfia Diferencia Entre Cloroplastos, Mitocondrías y Los Procesos EnergeticosDocumento1 páginaInfográfia Diferencia Entre Cloroplastos, Mitocondrías y Los Procesos EnergeticosDahiana CastroAún no hay calificaciones
- 2 Parcial Corrillo 8 CetogenesisDocumento5 páginas2 Parcial Corrillo 8 Cetogenesiskaren.pina050703Aún no hay calificaciones
- Tarea 5Documento6 páginasTarea 5Aurora Dannae Véliz GonzálezAún no hay calificaciones
- TP 12 - Cetogénesis y Cetolisis. Cetoacidosis - 2015Documento6 páginasTP 12 - Cetogénesis y Cetolisis. Cetoacidosis - 2015docsamu78Aún no hay calificaciones
- Biosíntesis de Cuerpos CetónicosDocumento14 páginasBiosíntesis de Cuerpos CetónicosClaris Ersz Vradika100% (1)
- 6 - Beta OxidacionDocumento10 páginas6 - Beta OxidacionValeria GrandeAún no hay calificaciones
- Aminoácidos EsencialesDocumento6 páginasAminoácidos EsencialesjavierAún no hay calificaciones
- CETOGÉNESISDocumento5 páginasCETOGÉNESISvianyse100% (1)
- Metabolismo Del Colesterol y Cuerpos Cetonicos.Documento4 páginasMetabolismo Del Colesterol y Cuerpos Cetonicos.alguiensoy9512Aún no hay calificaciones
- CetogénesisDocumento6 páginasCetogénesisEdwin Juvenal LlamoccaAún no hay calificaciones
- Metabolismo de Los Cuerpos CetonicosDocumento22 páginasMetabolismo de Los Cuerpos CetonicosMaría TobíasAún no hay calificaciones
- Cuerpos CetonicosDocumento2 páginasCuerpos CetonicosALEXIS APOLO OCHOAAún no hay calificaciones
- Metabolismo Cuerpos CetonicosDocumento19 páginasMetabolismo Cuerpos CetonicosLili VanAún no hay calificaciones
- Cetogenesis y CetolisisDocumento4 páginasCetogenesis y CetolisisLeonardo RestrepoAún no hay calificaciones
- Objeto PQV #Documento8 páginasObjeto PQV #Diana MAún no hay calificaciones
- Cuerpos Cetonicos 401637 Downloadable 4736415Documento4 páginasCuerpos Cetonicos 401637 Downloadable 4736415Deniska MalpicaAún no hay calificaciones
- Informe BioquimicaDocumento7 páginasInforme BioquimicaAnthony ChiliquingaAún no hay calificaciones
- 15.3 Metabolismo de Los Cuerpos CetónicosDocumento1 página15.3 Metabolismo de Los Cuerpos CetónicosJose Leonardo Vega OchoaAún no hay calificaciones
- Cuerpos Cetónicos - ResumenDocumento2 páginasCuerpos Cetónicos - ResumenDenisse MedinaAún no hay calificaciones
- Lípidos 1Documento25 páginasLípidos 1Abrahamalberto HernandezAún no hay calificaciones
- Explicación de La CetogénesisDocumento3 páginasExplicación de La CetogénesisAna MarcelaAún no hay calificaciones
- Resumen Completo de Los Cuerpos CetónicosDocumento2 páginasResumen Completo de Los Cuerpos CetónicosCarolina DuranAún no hay calificaciones
- Goloria BioquimicaDocumento13 páginasGoloria BioquimicaWillam Cadima garciaAún no hay calificaciones
- Cuerpos CetonicosDocumento6 páginasCuerpos CetonicosVirginia MullisacaAún no hay calificaciones
- Metabolismo de Los Cuerpo CetónicoDocumento7 páginasMetabolismo de Los Cuerpo CetónicoJordy Anthony Abanto RuizAún no hay calificaciones
- T-22 Cuerpos CetonicosDocumento19 páginasT-22 Cuerpos CetonicosDenis PeraltaAún no hay calificaciones
- Bioquimica Informe 2 Exposición (Cuerpos Cetónicos) - 2Documento4 páginasBioquimica Informe 2 Exposición (Cuerpos Cetónicos) - 2DavidArChambaAún no hay calificaciones
- CETOGÉNESISDocumento3 páginasCETOGÉNESISalessandra donoso ch.Aún no hay calificaciones
- CetogenesisDocumento9 páginasCetogenesisJessica PaolaAún no hay calificaciones
- Oxidacion de Acidos Grasos 2Documento5 páginasOxidacion de Acidos Grasos 2Marvin SantanaAún no hay calificaciones
- Cuerpos CetonicosDocumento6 páginasCuerpos CetonicosGaby Anicama FloresAún no hay calificaciones
- Trabaja de Investigacion-Cetogenesis-Estapas Enzimaticas-RegulacionDocumento13 páginasTrabaja de Investigacion-Cetogenesis-Estapas Enzimaticas-Regulacionpaolo bryan velasquez alvarezAún no hay calificaciones
- Bnu U2 Atr MarzDocumento5 páginasBnu U2 Atr Marzmarisol100% (1)
- Metabolismo Cuerpos Cetonicos 2014Documento17 páginasMetabolismo Cuerpos Cetonicos 2014Cristina RaittAún no hay calificaciones
- T-24 Cuerpos CetonicosDocumento14 páginasT-24 Cuerpos CetonicosAnonymous iIaMAfVAún no hay calificaciones
- Cuestionario 7 Cuerpos CetónicosDocumento7 páginasCuestionario 7 Cuerpos CetónicosALEJANDRO T. CARMONAAún no hay calificaciones
- w4 Clase Bioca Cetogenesis Cetolisis Xenob 20052020Documento32 páginasw4 Clase Bioca Cetogenesis Cetolisis Xenob 20052020Darreem Steve Villarreal PutnamAún no hay calificaciones
- AFPD2 Cómo Las Células Obtienen Energía de Los AlimentosDocumento21 páginasAFPD2 Cómo Las Células Obtienen Energía de Los AlimentosJIMENA ORTIZ SALDAÑAAún no hay calificaciones
- El Ciclo de KrebsDocumento7 páginasEl Ciclo de Krebsdario javier saltos quitoAún no hay calificaciones
- Tp10. Lipidos 1Documento26 páginasTp10. Lipidos 1ferminsAún no hay calificaciones
- Ciclo de KrebsDocumento12 páginasCiclo de KrebsGallardo RichardAún no hay calificaciones
- Principales Compuestos de Altas Energía Que Participan en Los Procesos MetabolitoDocumento5 páginasPrincipales Compuestos de Altas Energía Que Participan en Los Procesos MetabolitoJuan Diego RujanoAún no hay calificaciones
- Cuestionario BQMDocumento4 páginasCuestionario BQMBeatriz G.Aún no hay calificaciones
- CetogenesisDocumento8 páginasCetogenesisYuukiAuclair100% (1)
- CetogénesisDocumento20 páginasCetogénesisLIZBETH GERALDINE ESPINOZA RODRIGUEZ100% (1)
- Teorico 7 Metabolismo de Lipidos 2Documento27 páginasTeorico 7 Metabolismo de Lipidos 2Adriano pompeiAún no hay calificaciones
- Resumen MetabolismoDocumento9 páginasResumen MetabolismoFer RiveraAún no hay calificaciones
- Respiración Anaeróbica - AeróbicaDocumento7 páginasRespiración Anaeróbica - AeróbicaIrene Molenz100% (1)
- Ciclo de KrebsDocumento16 páginasCiclo de KrebsCarmen Flores100% (4)
- Mecanismo de Reaccion de Acido PalmiticoDocumento21 páginasMecanismo de Reaccion de Acido PalmiticoLetizia Flores LopezAún no hay calificaciones
- CicloK GeorgetteDocumento6 páginasCicloK Georgetteosmigdy mataAún no hay calificaciones
- Metabolismo de Los LípidosDocumento1 páginaMetabolismo de Los Lípidoscorey rubioAún no hay calificaciones
- Artículo Del Ciclo de Krebs Mayo 31 2018Documento5 páginasArtículo Del Ciclo de Krebs Mayo 31 2018Lesdy MarllyAún no hay calificaciones
- Cetolisis CetogenesisDocumento10 páginasCetolisis CetogenesisTania Huarcaya100% (1)
- TEMA 1 MetabolismoDocumento14 páginasTEMA 1 MetabolismoBerenize VidalAún no hay calificaciones
- Generalidades Del MetabolismoDocumento12 páginasGeneralidades Del MetabolismoMaria Paula GarciaAún no hay calificaciones
- La Ciencia Del Metabolismo: Metabolismo sin rodeosDe EverandLa Ciencia Del Metabolismo: Metabolismo sin rodeosCalificación: 5 de 5 estrellas5/5 (1)
- CosechaDocumento3 páginasCosechaDelgado Mejía BrandonAún no hay calificaciones
- MENTADocumento4 páginasMENTADelgado Mejía BrandonAún no hay calificaciones
- Metabolismo Del GlucogenoDocumento8 páginasMetabolismo Del GlucogenoDelgado Mejía BrandonAún no hay calificaciones
- Ciclo de Krebs y Vía AnapleróticasDocumento7 páginasCiclo de Krebs y Vía AnapleróticasDelgado Mejía BrandonAún no hay calificaciones
- Copia de Metabolismo Del GlucogenoDocumento25 páginasCopia de Metabolismo Del GlucogenoFitxXluisXx xAún no hay calificaciones
- Guia 3ro 2011Documento68 páginasGuia 3ro 2011nautilus19859760Aún no hay calificaciones
- ECV IsquemicoDocumento38 páginasECV Isquemicodaniel vargasAún no hay calificaciones
- Fisiología De, Los FarmacosDocumento6 páginasFisiología De, Los FarmacosMacarena InzunzaAún no hay calificaciones
- TEMA No 3 (Sinapsis)Documento9 páginasTEMA No 3 (Sinapsis)LuisUrquiolaAún no hay calificaciones
- Metabolismo CerebralDocumento12 páginasMetabolismo CerebralFranklinYanes100% (3)
- Glándulas Del Sistema Endocrino, Hormonas Que Secretan y FunciónDocumento4 páginasGlándulas Del Sistema Endocrino, Hormonas Que Secretan y FunciónMITZY YATZYRI VICTOR ARAUJO0% (1)
- Choice EndocrinoDocumento12 páginasChoice Endocrinoborja florezAún no hay calificaciones
- Apunte Sindrome Coronario Agudo 2019 (Oficial)Documento12 páginasApunte Sindrome Coronario Agudo 2019 (Oficial)Jotam Muñoz M100% (1)
- 6 7 y 8 GuytonDocumento6 páginas6 7 y 8 Guytontonolo100% (1)
- TermorregulacionDocumento8 páginasTermorregulacionDome ChávezAún no hay calificaciones
- Vasopresores e InotropicosDocumento25 páginasVasopresores e Inotropicosdavid saenzAún no hay calificaciones
- Borderline Personality Disorder Traits in Adolescents With AN EsDocumento9 páginasBorderline Personality Disorder Traits in Adolescents With AN Estatiana CastañedaAún no hay calificaciones
- Guia de Procedimiento en La Toma de Presion ArterialDocumento6 páginasGuia de Procedimiento en La Toma de Presion ArterialAmy AmyAún no hay calificaciones
- NEONATOLOGIADocumento66 páginasNEONATOLOGIAvirgilio alejandro mamani ordoñezAún no hay calificaciones
- ElectricidadDocumento1 páginaElectricidadDereck ChavezAún no hay calificaciones
- Casos Clinicos de Acido BaseDocumento3 páginasCasos Clinicos de Acido Baseanthony marc geerlings visagaAún no hay calificaciones
- Suturas de CardioDocumento14 páginasSuturas de CardioLaura OrozcoAún no hay calificaciones
- Diagnostico Diferencial SindromicoDocumento82 páginasDiagnostico Diferencial Sindromicogabriel pachecoAún no hay calificaciones
- Mecanismos Moleculares de La EpilepsiaDocumento10 páginasMecanismos Moleculares de La EpilepsiaValentina Salazar QuezadaAún no hay calificaciones
- Ariahtna Rodriguez-CTA-Tarea-9 PDFDocumento2 páginasAriahtna Rodriguez-CTA-Tarea-9 PDFBernarp Calin Hernanez LeonAún no hay calificaciones
- Practica N°8 - Inhibidores Cadena Respiratoria.Documento12 páginasPractica N°8 - Inhibidores Cadena Respiratoria.George NGAún no hay calificaciones
- Documento A4 Receta de Cookies Orgánico Verde y MarrónDocumento2 páginasDocumento A4 Receta de Cookies Orgánico Verde y MarrónLuciana JimenezAún no hay calificaciones
- 2018 Categorias APTA GUIDE 3.0Documento8 páginas2018 Categorias APTA GUIDE 3.0ana millanAún no hay calificaciones
- Actividad Reconocer Los Organos Del Cuerpo Humano y Su Funcion Ver 1Documento3 páginasActividad Reconocer Los Organos Del Cuerpo Humano y Su Funcion Ver 1Karii CorderoAún no hay calificaciones
- Practica 7Documento4 páginasPractica 7Mario Ceron RuizAún no hay calificaciones
- RD #000051-2021-Dg-Insnsb Guia de Procedimiento de Electrocardiografía Ambulatoria de 24Documento19 páginasRD #000051-2021-Dg-Insnsb Guia de Procedimiento de Electrocardiografía Ambulatoria de 24perezpeltrochegAún no hay calificaciones
- Catéter Swan-Ganz - Publicaciones CientíficasDocumento6 páginasCatéter Swan-Ganz - Publicaciones CientíficasMaideth Diaz AcostaAún no hay calificaciones