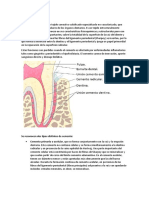
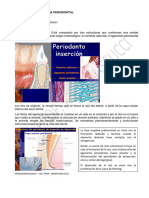
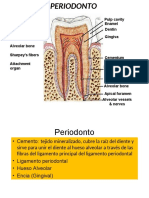
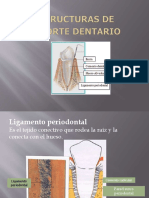
También podría gustarte
- 02 Estructura de Los Tej Periodntales en Indiv Sanos y EnferDocumento19 páginas02 Estructura de Los Tej Periodntales en Indiv Sanos y EnferFelipe YañezAún no hay calificaciones
- Histofisiologia PeriapicalDocumento14 páginasHistofisiologia PeriapicalLia YepezAún no hay calificaciones
- Ten Cate Oral Histology Development Structure and Function 9th Edition (Konkur - In) 448 450.en - EsDocumento3 páginasTen Cate Oral Histology Development Structure and Function 9th Edition (Konkur - In) 448 450.en - EsDenisse Márquez OrtegaAún no hay calificaciones
- Cemento Dental JMNRDocumento12 páginasCemento Dental JMNRJuan RoblesAún no hay calificaciones
- Ligamento Periodontal y CementoDocumento6 páginasLigamento Periodontal y CementodanielAún no hay calificaciones
- Cemento DentalDocumento24 páginasCemento DentalJuan Arturo80% (5)
- Bioquimica Dental CementoDocumento10 páginasBioquimica Dental CementoTIFFANY VANESSA DUEÑAS RADEAún no hay calificaciones
- Cemento RadicularDocumento16 páginasCemento RadicularDaniel Gamarra100% (1)
- Periodontología 2000Documento104 páginasPeriodontología 2000Porras Zamudio LeninAún no hay calificaciones
- Periodonto de InserciónDocumento11 páginasPeriodonto de InserciónAri MartinezAún no hay calificaciones
- El CementoDocumento9 páginasEl Cementoines valencia solanoAún no hay calificaciones
- Ten Cate Oral Histology Development Structure and Function 9th Edition (Konkur - In) 130 136.en - EsDocumento7 páginasTen Cate Oral Histology Development Structure and Function 9th Edition (Konkur - In) 130 136.en - EsDenisse Márquez OrtegaAún no hay calificaciones
- Periodoncio de InserciónDocumento16 páginasPeriodoncio de InserciónGianna ManziAún no hay calificaciones
- Práctica No. 6Documento4 páginasPráctica No. 6miguelsh0802Aún no hay calificaciones
- Formación Del Cemento Dental.Documento4 páginasFormación Del Cemento Dental.Brisa Espino TapiaAún no hay calificaciones
- Cuestionario PerioDocumento14 páginasCuestionario PerioSupercut StoreAún no hay calificaciones
- 10b Periodonto de Insercion 1Documento18 páginas10b Periodonto de Insercion 1Juli ViorelsAún no hay calificaciones
- Presentacion Periodonto ClasificacionesDocumento71 páginasPresentacion Periodonto ClasificacionesLeidi Diana MarlenneAún no hay calificaciones
- CEMENTO PeriodonciaDocumento5 páginasCEMENTO PeriodonciaNayAún no hay calificaciones
- Cementoblastos PDF InvestDocumento7 páginasCementoblastos PDF InvestLisa GarciaAún no hay calificaciones
- Histologia Cemento RadicularDocumento5 páginasHistologia Cemento Radicularrosa fariaAún no hay calificaciones
- Ligamento PeriodontalDocumento10 páginasLigamento Periodontalgenetica100% (33)
- Características Macroscópicas y Microscópicas de Los Tejidos de Inserción 2Documento11 páginasCaracterísticas Macroscópicas y Microscópicas de Los Tejidos de Inserción 2leidi40% (5)
- Resumen de Hueso y CementoDocumento9 páginasResumen de Hueso y CementoMigue MercuryAún no hay calificaciones
- Cemento, Ligamento Periodontal y Hueso AlveolarDocumento7 páginasCemento, Ligamento Periodontal y Hueso AlveolarVeronica Esteves100% (2)
- Informe de Hueso AlveolarDocumento14 páginasInforme de Hueso Alveolarzohan1889Aún no hay calificaciones
- Valoracion de Pilares en Protesis FijaDocumento161 páginasValoracion de Pilares en Protesis Fijacbustosw67% (12)
- Lectura 2 Anatomia Macroscopica Encia - LindheDocumento8 páginasLectura 2 Anatomia Macroscopica Encia - LindheKleiny Patricia Aranibar DextreAún no hay calificaciones
- 1 - Estructura de Los Tejidos Periodontales en El Individuo Sano y en El Individuo Enfermo PDFDocumento19 páginas1 - Estructura de Los Tejidos Periodontales en El Individuo Sano y en El Individuo Enfermo PDFsebis9Aún no hay calificaciones
- Expo Tejidos de SoporteDocumento39 páginasExpo Tejidos de SoporteArturo DuarteAún no hay calificaciones
- Cementogenesis GeneralidadesDocumento4 páginasCementogenesis GeneralidadesErika UrbinaAún no hay calificaciones
- Periodontología Clinica e Implantologia Odontologica.Documento48 páginasPeriodontología Clinica e Implantologia Odontologica.arturosanchezadss63% (49)
- Ligamento PeriodontalDocumento5 páginasLigamento PeriodontalfiorelitacabreraAún no hay calificaciones
- Cemento, Ligamento Periodontal, Hueso Alveolar y EncíaDocumento13 páginasCemento, Ligamento Periodontal, Hueso Alveolar y EncíaAndrea M.94% (16)
- Periodonto de SostenDocumento5 páginasPeriodonto de SostenJerolyn AvilaAún no hay calificaciones
- Pulpa NormalDocumento14 páginasPulpa NormalRomina MechehenAún no hay calificaciones
- Ligamento PeriodontalDocumento10 páginasLigamento PeriodontalNayAún no hay calificaciones
- No 6Documento6 páginasNo 6miguelsh0802Aún no hay calificaciones
- CementoDocumento31 páginasCementoDerly ContrerasAún no hay calificaciones
- Estructuras de Soporte DentalDocumento52 páginasEstructuras de Soporte DentalPilar ArellanoAún no hay calificaciones
- PeriiiiiDocumento4 páginasPeriiiiilauraRueda4Aún no hay calificaciones
- CementoDocumento50 páginasCementomiguelAún no hay calificaciones
- Cuestionario Tejidos PeriodontalesDocumento6 páginasCuestionario Tejidos PeriodontalesVerónica100% (1)
- Lectura LigamnetoDocumento7 páginasLectura LigamnetoAgustin guette oliverosAún no hay calificaciones
- BOrrador AvanceDocumento28 páginasBOrrador AvanceVictor ReyesAún no hay calificaciones
- ALVEOGENESISDocumento5 páginasALVEOGENESISKarla RojasAún no hay calificaciones
- Monografia CementoDocumento10 páginasMonografia CementoCesar Chigne100% (20)
- CementogenesisDocumento5 páginasCementogenesisCarlos HernandezAún no hay calificaciones
- Ligamento Periodontal RESUMENDocumento12 páginasLigamento Periodontal RESUMENRadhe GutiérrezAún no hay calificaciones
- Cementogénesis 098Documento9 páginasCementogénesis 098Luis Alejandro100% (4)
- 1punto HistoDocumento7 páginas1punto HistoMarlon CaillaguaAún no hay calificaciones
- Buco 2Documento10 páginasBuco 2Giovanni ArticaAún no hay calificaciones
- Elementos CelularesDocumento5 páginasElementos CelularesFrancescaFagianiAún no hay calificaciones
- Cuestionario 1 y 2 Periodoncia IIDocumento7 páginasCuestionario 1 y 2 Periodoncia IIMaria M Catacora MaldonadoAún no hay calificaciones
- Irrigantes locales en endodoncia de dientes primariosDe EverandIrrigantes locales en endodoncia de dientes primariosAún no hay calificaciones
- Reconstrucción de dientes endodonciados: Pautas de actuación clínicaDe EverandReconstrucción de dientes endodonciados: Pautas de actuación clínicaCalificación: 5 de 5 estrellas5/5 (4)
- Productos Finales de Glicación Avanzada y Estrés Oxidativo en La Diabetes Mellitus Tipo 2Documento11 páginasProductos Finales de Glicación Avanzada y Estrés Oxidativo en La Diabetes Mellitus Tipo 2Jhonatan RojasAún no hay calificaciones
- KiruDocumento5 páginasKiruJhonatan RojasAún no hay calificaciones
- MATRIZ DE CONSISTENCIA - Docx 2Documento2 páginasMATRIZ DE CONSISTENCIA - Docx 2Jhonatan RojasAún no hay calificaciones
- VariableDocumento1 páginaVariableJhonatan RojasAún no hay calificaciones
- Descripción de La Realidad Problemática - Docx ELABORACIÓN DE PROYECTOSDocumento25 páginasDescripción de La Realidad Problemática - Docx ELABORACIÓN DE PROYECTOSJhonatan RojasAún no hay calificaciones
- Periodontología Clinica e Implantologia Odontologica.Documento48 páginasPeriodontología Clinica e Implantologia Odontologica.arturosanchezadss63% (49)
- Traduccion OsteocitosDocumento4 páginasTraduccion OsteocitosJhonatan RojasAún no hay calificaciones
- Tercio Apical EndoDocumento21 páginasTercio Apical EndoJhonatan RojasAún no hay calificaciones
- Atención Estomatológica Del Paciente Con Asma BronquialDocumento1 páginaAtención Estomatológica Del Paciente Con Asma BronquialJhonatan RojasAún no hay calificaciones
- Primeros AuxiliosDocumento12 páginasPrimeros AuxiliosJhonatan RojasAún no hay calificaciones
- Factores Salivales Asociados A Prevalencia e Incremento de Caries Dental en Escolares RuralesDocumento1 páginaFactores Salivales Asociados A Prevalencia e Incremento de Caries Dental en Escolares RuralesJhonatan RojasAún no hay calificaciones
- Semana 24 - Ingles - SBSDocumento6 páginasSemana 24 - Ingles - SBSJhonatan RojasAún no hay calificaciones
- Aislamiento de Helicobacter Pylori en Sarro Dental de Pacientes Con Gastritis Del PoliclínicoDocumento2 páginasAislamiento de Helicobacter Pylori en Sarro Dental de Pacientes Con Gastritis Del PoliclínicoJhonatan RojasAún no hay calificaciones
- Fracturas EXPO ENFERMERIADocumento10 páginasFracturas EXPO ENFERMERIAJhonatan RojasAún no hay calificaciones
- Limpieza y DesinfeccionDocumento12 páginasLimpieza y DesinfeccionJhonatan RojasAún no hay calificaciones
- Asepsia y Antisepsia.Documento7 páginasAsepsia y Antisepsia.Jhonatan RojasAún no hay calificaciones
- Valores y MoralDocumento22 páginasValores y MoralJhonatan RojasAún no hay calificaciones
- Vocacion y Respo ProfesionalDocumento7 páginasVocacion y Respo ProfesionalJhonatan RojasAún no hay calificaciones
- PPR Tema 2 Componentes (Retenedores, Conectores, Bases y Dientes) PDFDocumento127 páginasPPR Tema 2 Componentes (Retenedores, Conectores, Bases y Dientes) PDFYaneth Cristina Morales BlasAún no hay calificaciones
- Semana 24 - Ingles - SBSDocumento6 páginasSemana 24 - Ingles - SBSJhonatan RojasAún no hay calificaciones
- Semana 24 - Ingles - SBSDocumento6 páginasSemana 24 - Ingles - SBSJhonatan RojasAún no hay calificaciones
- Revascularizacion - 2016Documento5 páginasRevascularizacion - 2016Jhonatan RojasAún no hay calificaciones
- Valores y MoralDocumento22 páginasValores y MoralJhonatan RojasAún no hay calificaciones
- Balotario Enao PDFDocumento125 páginasBalotario Enao PDFErling Beisaga EscobarAún no hay calificaciones
- 4.osteotomias Creaneomaxilofaciales Con UltrasonidoDocumento9 páginas4.osteotomias Creaneomaxilofaciales Con UltrasonidoJhonatan RojasAún no hay calificaciones
- PPR Tema 3 Paralelizado y Diseño PDFDocumento42 páginasPPR Tema 3 Paralelizado y Diseño PDFYaneth Cristina Morales Blas80% (25)
- PPR Tema 3 Paralelizado y Diseño PDFDocumento42 páginasPPR Tema 3 Paralelizado y Diseño PDFYaneth Cristina Morales Blas80% (25)
- Relacion Odontologo PacienteDocumento13 páginasRelacion Odontologo PacienteJhonatan RojasAún no hay calificaciones
- Introduccion AnatomiaDocumento100 páginasIntroduccion AnatomiaJhonatan RojasAún no hay calificaciones
- Protocolo de Servicio Inducto Conducción Del Trabajo de Parto A Término 1. IntroducciónDocumento12 páginasProtocolo de Servicio Inducto Conducción Del Trabajo de Parto A Término 1. IntroducciónGgr AsociadosAún no hay calificaciones
- Otros Estudiantes También Vieron: Células OseasDocumento25 páginasOtros Estudiantes También Vieron: Células OseasLizeth Gloria Jara PabloAún no hay calificaciones
- Tejido ConectivoDocumento2 páginasTejido ConectivoesnarmiAún no hay calificaciones
- Tarea 4Documento15 páginasTarea 4Wendy RuizAún no hay calificaciones
- Biologia Celular e Histologia 0Documento18 páginasBiologia Celular e Histologia 0Aranza AvAún no hay calificaciones
- Tejido OseoDocumento11 páginasTejido OseoSandro Fernandez ValerAún no hay calificaciones
- Tema-9 Tej Conjuntivo PDFDocumento12 páginasTema-9 Tej Conjuntivo PDFMelanieAún no hay calificaciones
- 21-09-2022 Tema 1 - Reparación Tisular - Omerling Varela (UNERG)Documento21 páginas21-09-2022 Tema 1 - Reparación Tisular - Omerling Varela (UNERG)cheot66Aún no hay calificaciones
- Cap1. La Celula Unidad de Salud y EnfermedadDocumento51 páginasCap1. La Celula Unidad de Salud y EnfermedadKevin Alejandro Arita ChinchillaAún no hay calificaciones
- Universidad Nacional de San Martin: Tejido CartilaginosoDocumento36 páginasUniversidad Nacional de San Martin: Tejido CartilaginosoCarolina DávilaAún no hay calificaciones
- Tejido ÓseoDocumento10 páginasTejido ÓseolourdessbravooAún no hay calificaciones
- Anat. (03) Tejido Conectivo 43 - 48Documento6 páginasAnat. (03) Tejido Conectivo 43 - 48claudia alvaAún no hay calificaciones
- Histologacartlago HuesoDocumento43 páginasHistologacartlago HuesoMartinez Martinez Edna Adaya100% (1)
- HISTOLOGÍADocumento23 páginasHISTOLOGÍACamila EscuderoAún no hay calificaciones
- Dossier Catalina CastroDocumento7 páginasDossier Catalina CastroCatalina Andrea Castro Montero 20825877Aún no hay calificaciones
- Capítulo 001Documento21 páginasCapítulo 001Deceno COLAún no hay calificaciones
- Tejidos TeoríaDocumento8 páginasTejidos TeoríaAaron Alexander Ceballos MorochoAún no hay calificaciones
- Tejido CartilaginosoDocumento10 páginasTejido CartilaginosoHERNÁNDEZ VEGA GIL ENRIQUE YOSEFAún no hay calificaciones
- A-Teorico U2 GlucidosDocumento19 páginasA-Teorico U2 GlucidosSebastian MoralesAún no hay calificaciones
- ReparaciónDocumento31 páginasReparaciónÁlvaro GarciaAún no hay calificaciones
- Portafolio HistoDocumento6 páginasPortafolio HistoReyes MilosAún no hay calificaciones
- Exposición de HistologíaDocumento3 páginasExposición de HistologíajavioletaAún no hay calificaciones
- Solemne I Histologia AvanzadaDocumento180 páginasSolemne I Histologia AvanzadaKarina SaezAún no hay calificaciones
- Catalogo Billabong Flows The Wave - 2023 09 28 08 17 37Documento30 páginasCatalogo Billabong Flows The Wave - 2023 09 28 08 17 37artfrankworkanaAún no hay calificaciones
- Traumatología y OrtopediaDocumento33 páginasTraumatología y OrtopediaRobertaAún no hay calificaciones
- Cuestionario de Histología N°3 - Tejido Conectivo No EspecializadoDocumento4 páginasCuestionario de Histología N°3 - Tejido Conectivo No Especializadosanmartinjosefa8Aún no hay calificaciones
- 8-Tejido Cartilaginoso.Documento41 páginas8-Tejido Cartilaginoso.Zoe RivarolaAún no hay calificaciones
- Revision de La Reparacion Del Cartilago Posibilidades y ResultadosDocumento18 páginasRevision de La Reparacion Del Cartilago Posibilidades y ResultadosAl CaAún no hay calificaciones
- PRACTICA 7 Observación de Células y Tejidos Básicos2Documento18 páginasPRACTICA 7 Observación de Células y Tejidos Básicos2Anderson Huaripata RojasAún no hay calificaciones
- Planificacion Biologia Celular y MolecularDocumento16 páginasPlanificacion Biologia Celular y MolecularDalays100% (1)