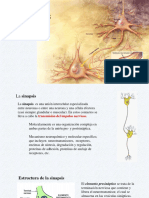
También podría gustarte
- Histología y Embrología del Sitema NerviosoDe EverandHistología y Embrología del Sitema NerviosoCalificación: 4 de 5 estrellas4/5 (4)
- 1 5 TejidoMuscularDocumento17 páginas1 5 TejidoMuscularOmar Sare CruzAún no hay calificaciones
- Inmunidad InnataDocumento14 páginasInmunidad InnataMelissa RodriguezAún no hay calificaciones
- FisioDocumento81 páginasFisioMafe MafeAún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
- Como Está Organizado El Cuerpo Humano 1Documento14 páginasComo Está Organizado El Cuerpo Humano 1Alexis AlonsoAún no hay calificaciones
- FISIOLOGIADocumento37 páginasFISIOLOGIAH DDAún no hay calificaciones
- Capítulo 4 de GuytonDocumento7 páginasCapítulo 4 de GuytonRosario GarciaAún no hay calificaciones
- Parcial 1Documento12 páginasParcial 1gaston lujanAún no hay calificaciones
- 1er SEMINARIO BIOPOTENCIALESDocumento15 páginas1er SEMINARIO BIOPOTENCIALESHannah Barlow100% (1)
- Barreto. Taller 1 Fisiología CelularDocumento19 páginasBarreto. Taller 1 Fisiología CelularOscar BarretoAún no hay calificaciones
- Celula 1Documento13 páginasCelula 1Florencia9 GhioAún no hay calificaciones
- 3 La Membrana Celularpara PresentarDocumento60 páginas3 La Membrana Celularpara PresentaryaninaAún no hay calificaciones
- DirseuHernández - 304 - Ensayo - Fisiología de La Membrana El Nervio y El MúsculoDocumento1 páginaDirseuHernández - 304 - Ensayo - Fisiología de La Membrana El Nervio y El MúsculoDirseu VillalobosAún no hay calificaciones
- Cap 2Documento36 páginasCap 2Alfredo CalispaAún no hay calificaciones
- Preguntas Fisiologia 3Documento12 páginasPreguntas Fisiologia 3Tiago AngrisaniAún no hay calificaciones
- Neuro TrabajoDocumento13 páginasNeuro TrabajoMau TorresAún no hay calificaciones
- Guia Ordinario FisioDocumento17 páginasGuia Ordinario Fisio2030 Mondragón Castillo María FernandaAún no hay calificaciones
- FisiologíaDocumento4 páginasFisiologíaARIAS ZELA DANIELA ALEJANDRAAún no hay calificaciones
- Tejidos ExcitablesDocumento4 páginasTejidos ExcitablesCandice CalderonAún no hay calificaciones
- Bomba de Sodio PotasioDocumento8 páginasBomba de Sodio PotasioEdemi PerezAún no hay calificaciones
- Resumen Cap1 5 6 7 y 8 Guyton e Hall Fisiologia Medica 13 EdDocumento20 páginasResumen Cap1 5 6 7 y 8 Guyton e Hall Fisiologia Medica 13 EdarielexAún no hay calificaciones
- 1transmisión NeuromuscularDocumento5 páginas1transmisión NeuromuscularJean Pierre Jiron SosaAún no hay calificaciones
- Fisiología de La Membrana, El Nervio y El Músculo.Documento14 páginasFisiología de La Membrana, El Nervio y El Músculo.Gabriel GuedezAún no hay calificaciones
- Fisio 2Documento17 páginasFisio 2Ana JimenezAún no hay calificaciones
- Sinapsis e Impulso NerviosoDocumento40 páginasSinapsis e Impulso NerviosogregorioaaAún no hay calificaciones
- Final Biología - HolaDocumento63 páginasFinal Biología - HolaFrancisco SauritAún no hay calificaciones
- Ensayo Brice Ñ oDocumento8 páginasEnsayo Brice Ñ oMaria Estefany GotaAún no hay calificaciones
- RibosomaDocumento5 páginasRibosomaYessica Mendoza MaqueraAún no hay calificaciones
- Seminario de PotencialesDocumento14 páginasSeminario de PotencialesAlexanther Palomino ReateguiAún no hay calificaciones
- FPB EsquemaDocumento52 páginasFPB EsquemamontseruizlopAún no hay calificaciones
- Núcleo y Ácidos NucleicosDocumento26 páginasNúcleo y Ácidos NucleicosAlexka Silva CamachoAún no hay calificaciones
- Potenciales de Accion y de MembranaDocumento6 páginasPotenciales de Accion y de MembranaModering PimentelAún no hay calificaciones
- Bases Biofísicas Del ElectrocardiogramaDocumento15 páginasBases Biofísicas Del Electrocardiogramaneomartin666Aún no hay calificaciones
- FinalDocumento63 páginasFinalAinara zubillagaAún no hay calificaciones
- La CélulaDocumento10 páginasLa CélulaJudith cordero peñaAún no hay calificaciones
- HistoDocumento16 páginasHistoVictoria RodriguezAún no hay calificaciones
- Fisiologia de Los Tejidos ExcitablesDocumento6 páginasFisiologia de Los Tejidos Excitablesjoceline.duenasAún no hay calificaciones
- Potencial de Acción, Sinápsis y Neurotransmisores-Guias de Estudio PsicobiologíaDocumento11 páginasPotencial de Acción, Sinápsis y Neurotransmisores-Guias de Estudio PsicobiologíaLaura RoaAún no hay calificaciones
- Informe Set 1 FisiologiaDocumento10 páginasInforme Set 1 FisiologiaAlejandro RomeroAún no hay calificaciones
- 04 Membrana PlasmáticaDocumento14 páginas04 Membrana PlasmáticaCamila RamirezAún no hay calificaciones
- Apuntes Bioelectromagnetismo PDFDocumento75 páginasApuntes Bioelectromagnetismo PDFsjhg13Aún no hay calificaciones
- Estructura Musculo EsqueléticoDocumento5 páginasEstructura Musculo EsqueléticoAlondraAún no hay calificaciones
- Fisiología 1 Trabajo 2 Susan AndreaDocumento10 páginasFisiología 1 Trabajo 2 Susan AndreaSUSAN ANDREA DORADOAún no hay calificaciones
- Expo 1 Fisio MuscularDocumento24 páginasExpo 1 Fisio MuscularYamilee NikoleAún no hay calificaciones
- El CitoesqueletoDocumento8 páginasEl CitoesqueletoShirley CastilloAún no hay calificaciones
- Bio Potencial EsDocumento23 páginasBio Potencial EsRossaly Perez NiñoAún no hay calificaciones
- 0) Guia Definitiva de Excitabilidad CelularDocumento10 páginas0) Guia Definitiva de Excitabilidad CelularRodrigo PujadasAún no hay calificaciones
- FISIOLOGIADocumento5 páginasFISIOLOGIARosivell RivasAún no hay calificaciones
- Membrana de Las CelulasDocumento4 páginasMembrana de Las CelulasDailton AntunezAún no hay calificaciones
- Fisio 1 y 2Documento62 páginasFisio 1 y 2Alejandra MuñozAún no hay calificaciones
- Secuencias Segunda Etapa Del AñoDocumento35 páginasSecuencias Segunda Etapa Del AñoÁlvaro ZapataAún no hay calificaciones
- T7 Membranas BiologicasDocumento5 páginasT7 Membranas Biologicasruben pacheco alonsoAún no hay calificaciones
- Fundamentos de La Actividad Fisiológica (La Célula)Documento64 páginasFundamentos de La Actividad Fisiológica (La Célula)IviAún no hay calificaciones
- Membrana Celular Mapa Conceptual..... Norma 5..Documento8 páginasMembrana Celular Mapa Conceptual..... Norma 5..Jamirton Erledy GarciaAún no hay calificaciones
- T7 La Menbrana Plasmática, EtcDocumento20 páginasT7 La Menbrana Plasmática, Etcdiana.redondo2004Aún no hay calificaciones
- Apuntes BIOELECTROMAGNETISMO - 1 - 2010Documento76 páginasApuntes BIOELECTROMAGNETISMO - 1 - 2010ayaloAún no hay calificaciones
- Cuadernillo de Fisio 1Documento23 páginasCuadernillo de Fisio 1Carlos LeguizaAún no hay calificaciones
- Transporte Y MusculosDocumento1 páginaTransporte Y MusculosJose Antonio Alcantara SelfaAún no hay calificaciones
- LinfopoyesisDocumento12 páginasLinfopoyesisMar MeléndezAún no hay calificaciones
- Mapas Conceptuales BiomoleculasDocumento7 páginasMapas Conceptuales Biomoleculasjahaziel urrutiaAún no hay calificaciones
- Apuntes Conocimiento Del Medio Tema 1Documento3 páginasApuntes Conocimiento Del Medio Tema 1RubeenMaAún no hay calificaciones
- Anatomia Muscular Rumientes PDFDocumento3 páginasAnatomia Muscular Rumientes PDFlaura garciaAún no hay calificaciones
- Teor 9 Hemostasis 2019Documento44 páginasTeor 9 Hemostasis 2019Flavia AntúnezAún no hay calificaciones
- Tarea 1 - Reconocer Los Principios de La Teoría CelularDocumento9 páginasTarea 1 - Reconocer Los Principios de La Teoría CelularrAún no hay calificaciones
- Preguntas Generadoras Tatiana T3Documento4 páginasPreguntas Generadoras Tatiana T3Felipe CastañedaAún no hay calificaciones
- Diagrama de Tipos de ClonacionDocumento2 páginasDiagrama de Tipos de ClonacionVictor RequejoAún no hay calificaciones
- Clase 6 Guía Fisiología Muscular y VentilaciónDocumento20 páginasClase 6 Guía Fisiología Muscular y VentilaciónAlejandro Andrés Rocha NarváezAún no hay calificaciones
- Informe OsmosisDocumento10 páginasInforme Osmosissanti0% (1)
- Difusion Osmosis PDFDocumento19 páginasDifusion Osmosis PDFAngel OlivaAún no hay calificaciones
- Codigo 20genetico 20y 20Traduccià N 20de 20proteinasDocumento14 páginasCodigo 20genetico 20y 20Traduccià N 20de 20proteinasSantiago Delgado Chumioque0% (1)
- Trip TicoDocumento1 páginaTrip TicoCarolina ChoezAún no hay calificaciones
- Orgánulos No Membranosos 2018Documento5 páginasOrgánulos No Membranosos 2018María Dolores Muñoz CastroAún no hay calificaciones
- Sistema TegumentarioDocumento31 páginasSistema TegumentarioCarlos Moreno RuizAún no hay calificaciones
- 34-B-24!04!19 Continuancion Vitamina B1-Coe Pal - mp3Documento3 páginas34-B-24!04!19 Continuancion Vitamina B1-Coe Pal - mp3Mosanjo Chavez ElbertAún no hay calificaciones
- Biología Clase 13-14Documento29 páginasBiología Clase 13-14ASMAún no hay calificaciones
- 7 Biología.Documento9 páginas7 Biología.ANA MILENA ANTEQUERA MENDOZAAún no hay calificaciones
- Exposición S8 - Moléculas de Relación CelularDocumento17 páginasExposición S8 - Moléculas de Relación CelularAlejandro SuaresAún no hay calificaciones
- Artículo 7Documento6 páginasArtículo 7Cele Lbo JaraAún no hay calificaciones
- Respuesta Inmune Frente A Infecciones ViralesDocumento3 páginasRespuesta Inmune Frente A Infecciones ViralesFabiola CornielesAún no hay calificaciones
- Composición Química Del Tejido MuscularDocumento2 páginasComposición Química Del Tejido MuscularVictor Corp100% (1)
- Tipo de Orden Número ReqDocumento4 páginasTipo de Orden Número ReqlcamineroAún no hay calificaciones
- Transducción de SeñalesDocumento17 páginasTransducción de SeñalesKenia ContrerasAún no hay calificaciones
- Marcadores TumoralesDocumento45 páginasMarcadores TumoralesYazmin L. FlowersAún no hay calificaciones
- Regeneración AutonomaDocumento18 páginasRegeneración AutonomaVelazquez AngelAún no hay calificaciones
- Nutricion ClinicaDocumento138 páginasNutricion ClinicaGaston Elias GimenezAún no hay calificaciones
- Formato 2015 ANATO - 1Documento6 páginasFormato 2015 ANATO - 1johancarvalAún no hay calificaciones