También podría gustarte
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Virus RabiaDocumento36 páginasVirus RabiaMarcelo RealAún no hay calificaciones
- Compartimientos de Replicación de Virus DNADocumento32 páginasCompartimientos de Replicación de Virus DNAMarcelo Collantes CheloAún no hay calificaciones
- Proteínas. Biología Molecular.Documento27 páginasProteínas. Biología Molecular.Sofía A. SánchezAún no hay calificaciones
- VirusDocumento12 páginasVirusGeysiAún no hay calificaciones
- Taller # 2 BioquimicaDocumento4 páginasTaller # 2 BioquimicaFelipe MartinezAún no hay calificaciones
- Diabetes Tipo 2Documento24 páginasDiabetes Tipo 2Estefania AcuñaAún no hay calificaciones
- Orto y ParamyxovirusDocumento52 páginasOrto y ParamyxovirusJb HdzAún no hay calificaciones
- Matlab Basico - ComandosDocumento56 páginasMatlab Basico - ComandosIsaac Manuel Rodriguez LavadoAún no hay calificaciones
- Clase 4 - Transcripción y TraducciónDocumento52 páginasClase 4 - Transcripción y TraducciónConsue HAPPYAún no hay calificaciones
- Virus de La InfluenzaDocumento42 páginasVirus de La InfluenzaAndrea AltamiranoAún no hay calificaciones
- t9 MV - Togavirus - RubeolaDocumento50 páginast9 MV - Togavirus - RubeolaAnge M. H. DávilaAún no hay calificaciones
- Patogenia Del Virus de La RabiaDocumento12 páginasPatogenia Del Virus de La RabiaSaul Sandoval0% (1)
- CAPÍTULO 42 - Rabia, Infecciones Por Virus Lentos y Enfermedades PriónicasDocumento16 páginasCAPÍTULO 42 - Rabia, Infecciones Por Virus Lentos y Enfermedades Priónicas2215237eAún no hay calificaciones
- Regulación de La Síntesis de Galactosidasa En: Escherichia ColiDocumento53 páginasRegulación de La Síntesis de Galactosidasa En: Escherichia ColiZarate Arrieta Erika VianeyAún no hay calificaciones
- Modificaciones PostranscripcionalesDocumento25 páginasModificaciones PostranscripcionaleslizbethAún no hay calificaciones
- Virus de La Rabia BovinaDocumento19 páginasVirus de La Rabia BovinaAndre Suarez FarfanAún no hay calificaciones
- Clase 13 Traducción - Regulación Expresión-BiolDocumento53 páginasClase 13 Traducción - Regulación Expresión-BiolKatherinne Perez SanchezAún no hay calificaciones
- Epigentica (1) .....Documento20 páginasEpigentica (1) .....Gonzalo LunacAún no hay calificaciones
- Sesión 13 - Replicación-Transcripción y TraducciónDocumento25 páginasSesión 13 - Replicación-Transcripción y TraducciónKaren Crisologo BuenoAún no hay calificaciones
- CLASE #3 Los Virus, Viroides y PrionesDocumento23 páginasCLASE #3 Los Virus, Viroides y PrionesMarleni SilvaAún no hay calificaciones
- TraducciónDocumento37 páginasTraducciónJaviera ArellanoAún no hay calificaciones
- Transcripción 1Documento37 páginasTranscripción 1Javiera ArellanoAún no hay calificaciones
- CAPÍTULO 42 - Rabia, Infecciones Por Virus Lentos y Enfermedades PriónicasDocumento16 páginasCAPÍTULO 42 - Rabia, Infecciones Por Virus Lentos y Enfermedades PriónicasJeraldiin BeltranAún no hay calificaciones
- ACFrOgAIlsAP0mHG2ezLfubBfI - 2iWQUg79u1EykduEwzfwDmIIscvG3meqaOehPlPPmTBLf4nWrR1EivV35KYh5INK3TQ ZN iYY4g2oc1rSCu - aCCK4wip4xTBqzhR 2JxsPrQ1QK07Documento95 páginasACFrOgAIlsAP0mHG2ezLfubBfI - 2iWQUg79u1EykduEwzfwDmIIscvG3meqaOehPlPPmTBLf4nWrR1EivV35KYh5INK3TQ ZN iYY4g2oc1rSCu - aCCK4wip4xTBqzhR 2JxsPrQ1QK07César HaroAún no hay calificaciones
- 3 Genetica ViralDocumento40 páginas3 Genetica ViralErnesto AmaroAún no hay calificaciones
- Proyectos 5tos 2023: Proyecto TODocumento22 páginasProyectos 5tos 2023: Proyecto TOSlandyAún no hay calificaciones
- S030028961531382XDocumento9 páginasS030028961531382XDiana LunaAún no hay calificaciones
- Biologia Molecular III - Dr. Martín Rodríguez 2023Documento70 páginasBiologia Molecular III - Dr. Martín Rodríguez 2023Maria Gabriela Urdaneta ZubalevichAún no hay calificaciones
- Prats Virus Rna Con EnvolturaDocumento10 páginasPrats Virus Rna Con EnvolturaAdrilupiss ArroyoAún no hay calificaciones
- Dogma CentralDocumento64 páginasDogma CentralNally JumboAún no hay calificaciones
- Reovirus y RotavirusDocumento3 páginasReovirus y RotavirusDalila CarreñoAún no hay calificaciones
- TAREA 1. Introducción GenómicaDocumento8 páginasTAREA 1. Introducción GenómicaMariana MezaAún no hay calificaciones
- Transcripcion Del AdnDocumento19 páginasTranscripcion Del AdnSayda Alisson Huacre TuctoAún no hay calificaciones
- Características Principales Del Genoma VIHDocumento4 páginasCaracterísticas Principales Del Genoma VIHRene VelasquezAún no hay calificaciones
- Unidad 3 TranscripcionDocumento31 páginasUnidad 3 TranscripcionAngelAún no hay calificaciones
- Exposición. Traducción GeneticaDocumento46 páginasExposición. Traducción GeneticaFairyox RavenAún no hay calificaciones
- Clase I, II y III Abril-Mayo 2016Documento70 páginasClase I, II y III Abril-Mayo 2016evanconstantine77Aún no hay calificaciones
- Tema 2 Replicación y Reparación Del DNADocumento82 páginasTema 2 Replicación y Reparación Del DNAAnabel GutierrezAún no hay calificaciones
- Respuestas Genética y Biología MolecularDocumento3 páginasRespuestas Genética y Biología MolecularSerCL1Aún no hay calificaciones
- Sesión 13 - Replicación-Transcripción y TraducciónDocumento24 páginasSesión 13 - Replicación-Transcripción y TraducciónJAIRO KICK ENRIQUEZ FLORESAún no hay calificaciones
- Patogenia Microbiana-Priones y VirusDocumento25 páginasPatogenia Microbiana-Priones y VirusGabriela CastroAún no hay calificaciones
- 6-Ac NucleicosDocumento56 páginas6-Ac NucleicosSofi VasquezAún no hay calificaciones
- Cuestionario de Antecedentes y Procedimiento Experimental RegulacionDocumento22 páginasCuestionario de Antecedentes y Procedimiento Experimental RegulacionIVON DE LA CRUZ MORENOAún no hay calificaciones
- Transcripción 17Documento19 páginasTranscripción 17Claudio ManríquezAún no hay calificaciones
- BC 2021 Seminario 4 Regulacion de La Expresion de GenesDocumento59 páginasBC 2021 Seminario 4 Regulacion de La Expresion de GenesEmilio Sáez ValdésAún no hay calificaciones
- Genoma 2021Documento65 páginasGenoma 2021carlos davilaAún no hay calificaciones
- SARS CoV 2Documento29 páginasSARS CoV 2Daniel Ricardo Rodriguez AlvaradoAún no hay calificaciones
- Tema 5.4. Transcripción Del ADN A ARNDocumento59 páginasTema 5.4. Transcripción Del ADN A ARNCRISTIAN ALONSO RODRIGUEZ GONZALEZAún no hay calificaciones
- Parcial 2A BQ IIDocumento2 páginasParcial 2A BQ IIMaia CucchianiAún no hay calificaciones
- Taller 3 FinalDocumento55 páginasTaller 3 FinalAlejandro García RigualAún no hay calificaciones
- Seminario 6Documento57 páginasSeminario 6Leydi BravoAún no hay calificaciones
- Expo Hepatitis BDocumento24 páginasExpo Hepatitis BKevin AlquingaAún no hay calificaciones
- Alberts Capítulo Mecanismos Genéticos Básicos - Replicación, Transcripción, TraducciónDocumento44 páginasAlberts Capítulo Mecanismos Genéticos Básicos - Replicación, Transcripción, Traducciónjj romeroAún no hay calificaciones
- Enfermedad Granulomatosa CrónicaDocumento10 páginasEnfermedad Granulomatosa CrónicaJuan Diego Felix CoronaAún no hay calificaciones
- 11 Teoría BIología Molecular AplicadaDocumento54 páginas11 Teoría BIología Molecular Aplicadaivany maricielo velasquez calderonAún no hay calificaciones
- Wuolah Free TEMA 7Documento3 páginasWuolah Free TEMA 7ceciliasaavedra1604Aún no hay calificaciones
- Clase 5 Del ADN A Las ProteinasDocumento116 páginasClase 5 Del ADN A Las ProteinasBenito VegaAún no hay calificaciones
- Transcripción y Traducción en Organismos ProcariotasDocumento2 páginasTranscripción y Traducción en Organismos ProcariotasLuis Mario MartinezAún no hay calificaciones
- VitaminasDocumento38 páginasVitaminasFranche OportoAún no hay calificaciones
- HerpesDocumento11 páginasHerpesFranche OportoAún no hay calificaciones
- Virus IntestinalesDocumento4 páginasVirus IntestinalesFranche OportoAún no hay calificaciones
- HipovitaminosisDocumento3 páginasHipovitaminosisFranche OportoAún no hay calificaciones
- HepatitisDocumento42 páginasHepatitisFranche OportoAún no hay calificaciones
- AmebiasisDocumento9 páginasAmebiasisFranche OportoAún no hay calificaciones
- GiardiasisDocumento8 páginasGiardiasisFranche OportoAún no hay calificaciones
- AdenovirusDocumento3 páginasAdenovirusFranche OportoAún no hay calificaciones
- La Definición de La Corrupción y Clase SocialDocumento17 páginasLa Definición de La Corrupción y Clase SocialSandra Milena ReinaAún no hay calificaciones
- RD Ci2 134 20Documento13 páginasRD Ci2 134 20Jose MendozaAún no hay calificaciones
- Salud y EducaciónDocumento3 páginasSalud y Educaciónalicia nieto postigoAún no hay calificaciones
- 1601-P-Asg-02-V2 Procedimiento Manejo de Novedades Base de Datos Regimen SubsidiadoDocumento6 páginas1601-P-Asg-02-V2 Procedimiento Manejo de Novedades Base de Datos Regimen Subsidiadojose l tous diazAún no hay calificaciones
- Modelo His TBDocumento4 páginasModelo His TBWalter Delgado RosasAún no hay calificaciones
- Articulo Cientifico CerealesDocumento15 páginasArticulo Cientifico Cerealesamicarelli100% (1)
- FT Queso Campesino PasteurizadoDocumento3 páginasFT Queso Campesino PasteurizadoWilthon F Urbano100% (3)
- Castillos en El AireDocumento54 páginasCastillos en El AireFlor MagnaAún no hay calificaciones
- 08 El Ser Humano La Reproduccion ProfesoradoDocumento8 páginas08 El Ser Humano La Reproduccion ProfesoradoJose Gregorio OrtegaAún no hay calificaciones
- Boe S 2021 237Documento17 páginasBoe S 2021 237Javier De La Mora MirandaAún no hay calificaciones
- Informe Avance Muestreo Plagas Maíz. Agosto 2016Documento14 páginasInforme Avance Muestreo Plagas Maíz. Agosto 2016Gustavo De LeónAún no hay calificaciones
- Informe de Imagenes de Ets-Victoria Jamanca AstoDocumento24 páginasInforme de Imagenes de Ets-Victoria Jamanca AstoSede Operativa Sanchez CarriónAún no hay calificaciones
- Platica de 5 Min - INCENDIOS 09 04 2021Documento2 páginasPlatica de 5 Min - INCENDIOS 09 04 2021Jesus ValdezAún no hay calificaciones
- Introduccion y ConclusionesDocumento2 páginasIntroduccion y Conclusionesyelko donairesAún no hay calificaciones
- Diaz S Duiii ExpoDocumento21 páginasDiaz S Duiii Expomainares67Aún no hay calificaciones
- Perfil Del Adolescente Infractor en El PerúDocumento14 páginasPerfil Del Adolescente Infractor en El PerúCM GalAún no hay calificaciones
- Taller Proyecto de InvestigacionDocumento8 páginasTaller Proyecto de InvestigacionDaniela AlejandraAún no hay calificaciones
- Cómo Equipar Una Sala de Juego TerapeuticoDocumento5 páginasCómo Equipar Una Sala de Juego TerapeuticoEmmanuel Maldonado.Aún no hay calificaciones
- FUROSEMIDADocumento3 páginasFUROSEMIDAoscarAún no hay calificaciones
- Ciclo Menstrual Persepcion y Rendimiento en Jugadoras de Futbol de ÉliteDocumento9 páginasCiclo Menstrual Persepcion y Rendimiento en Jugadoras de Futbol de ÉliteRicardo DelfosAún no hay calificaciones
- Sepsis en PediatríaDocumento23 páginasSepsis en PediatríaDaniela QuijadaAún no hay calificaciones
- Programa Del 13° Congreso de ASETDocumento44 páginasPrograma Del 13° Congreso de ASETEzequiel Monteforte100% (1)
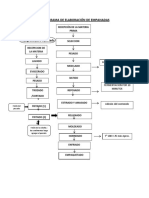
- Flujograma de Elaboración de EmpanadasDocumento5 páginasFlujograma de Elaboración de Empanadasflor cano padilla33% (3)
- Biopsicologia Resumen Parcial 1Documento45 páginasBiopsicologia Resumen Parcial 1Paula GPAún no hay calificaciones
- Promoción de Comportamientos SaludablesDocumento4 páginasPromoción de Comportamientos SaludablesJuan Esteban LopezAún no hay calificaciones
- Cátedra Farmacología. Trimestre Nov 12-Ene 13Documento230 páginasCátedra Farmacología. Trimestre Nov 12-Ene 13Jose Tomas NuñezAún no hay calificaciones
- Ennis Flint Glass Beads SDSDocumento6 páginasEnnis Flint Glass Beads SDSWilliams VázquezAún no hay calificaciones
- Control de Calidad en La Industria FarmaceuticaDocumento39 páginasControl de Calidad en La Industria FarmaceuticaRenzo Fernancarri100% (3)
- PED San Jose de Ocoa 2014 2024Documento100 páginasPED San Jose de Ocoa 2014 2024Jason TejedaAún no hay calificaciones
- Ingeniería Ambiental Uni Plan de EstudiosDocumento5 páginasIngeniería Ambiental Uni Plan de EstudiosJunior Marc Alcantara SantamariaAún no hay calificaciones
- Homo antecessor: El nacimiento de una especieDe EverandHomo antecessor: El nacimiento de una especieCalificación: 5 de 5 estrellas5/5 (1)
- Cerebro y silencio: Las claves de la creatividad y la serenidadDe EverandCerebro y silencio: Las claves de la creatividad y la serenidadCalificación: 5 de 5 estrellas5/5 (2)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- Liberación del trauma: Perdón y temblor es el caminoDe EverandLiberación del trauma: Perdón y temblor es el caminoCalificación: 4 de 5 estrellas4/5 (4)
- Muchas Vidas, Muchos Sabios (Many Lives, Many Masters): (Many Lives, Many Masters)De EverandMuchas Vidas, Muchos Sabios (Many Lives, Many Masters): (Many Lives, Many Masters)Calificación: 4 de 5 estrellas4/5 (475)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- Dieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasDe EverandDieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasCalificación: 4 de 5 estrellas4/5 (9)
- Sesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesDe EverandSesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesCalificación: 4.5 de 5 estrellas4.5/5 (13)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Cerebros rotos: Pacientes asombrosos que me enseñaron a vivirDe EverandCerebros rotos: Pacientes asombrosos que me enseñaron a vivirCalificación: 5 de 5 estrellas5/5 (3)
- Sana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónDe EverandSana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónCalificación: 5 de 5 estrellas5/5 (4)
- Historia del cuerpo humano: Evolución, salud y enfermedadDe EverandHistoria del cuerpo humano: Evolución, salud y enfermedadAún no hay calificaciones
- Lactancia humana y fonoaudiología: Guía para madres lactantesDe EverandLactancia humana y fonoaudiología: Guía para madres lactantesCalificación: 5 de 5 estrellas5/5 (4)
- Cambiar el pasado: Superar las experiencias traumáticas con la terapia estratégicaDe EverandCambiar el pasado: Superar las experiencias traumáticas con la terapia estratégicaCalificación: 5 de 5 estrellas5/5 (4)
- Guía práctica para la refracción ocularDe EverandGuía práctica para la refracción ocularCalificación: 5 de 5 estrellas5/5 (2)
- Hematología. Casos clínicos: preguntas y respuestasDe EverandHematología. Casos clínicos: preguntas y respuestasCalificación: 4 de 5 estrellas4/5 (4)
- El Libro de la Dieta Antiinflamatoria: Plan de 14 días para Sanar el Sistema inmunológico y Sentirte Mejor que NuncaDe EverandEl Libro de la Dieta Antiinflamatoria: Plan de 14 días para Sanar el Sistema inmunológico y Sentirte Mejor que NuncaCalificación: 4.5 de 5 estrellas4.5/5 (14)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Diseño, construcción y pruebas de horno prototipo semicontinuo para producir cerámicaDe EverandDiseño, construcción y pruebas de horno prototipo semicontinuo para producir cerámicaAún no hay calificaciones
- Ondas de choque extracorpóreas radiales: Aplicación en patologías músculo esqueléticasDe EverandOndas de choque extracorpóreas radiales: Aplicación en patologías músculo esqueléticasCalificación: 5 de 5 estrellas5/5 (3)
- Terapia de vidas pasadas: Un camino hacia la luz del alma. Técnica y prácticaDe EverandTerapia de vidas pasadas: Un camino hacia la luz del alma. Técnica y prácticaCalificación: 4.5 de 5 estrellas4.5/5 (11)