También podría gustarte
- Ceneval Odonto PDFDocumento49 páginasCeneval Odonto PDFGeraldo Pozos Vazquez100% (1)
- Nitrox Manual - Iantd SpainDocumento42 páginasNitrox Manual - Iantd SpainSanti Ramos Martin100% (2)
- Epigenética y Ser Humano PDFDocumento7 páginasEpigenética y Ser Humano PDFDaniel Felipe Marin VanegasAún no hay calificaciones
- Burton y RoseDocumento102 páginasBurton y RoseJem Severino VillacísAún no hay calificaciones
- IntroDocumento4 páginasIntroCatie P ParedesAún no hay calificaciones
- Microbios Intestinales Moldean El Comportamiento ESPDocumento8 páginasMicrobios Intestinales Moldean El Comportamiento ESPKing GeorgeAún no hay calificaciones
- Eje Microbiota Intestino CerebroDocumento3 páginasEje Microbiota Intestino CerebroLuciAngelik VaMe100% (2)
- Sherwin2017 Es - UnlockedDocumento41 páginasSherwin2017 Es - UnlockedMaria José OchoaAún no hay calificaciones
- Eje Microbiota-Intestino-CerebroDocumento3 páginasEje Microbiota-Intestino-CerebroBryan Echeverria Vasquez100% (1)
- Ane223e UnlockedDocumento9 páginasAne223e UnlockedLuismi DocAún no hay calificaciones
- Eje Cerebro Intestino MicrobiotaDocumento11 páginasEje Cerebro Intestino Microbiotaestefania.valencia02Aún no hay calificaciones
- Cómo Afecta La Microbiota en La Enfermedad Del ParkinsonDocumento4 páginasCómo Afecta La Microbiota en La Enfermedad Del ParkinsonRosy NaNoAún no hay calificaciones
- Capitulo Iii MarianaDocumento5 páginasCapitulo Iii MarianaMariana BermudezAún no hay calificaciones
- Bioquimica Del AlzheimerDocumento8 páginasBioquimica Del AlzheimerLilia González parianAún no hay calificaciones
- BBM1 U2 A2 Vabc.Documento6 páginasBBM1 U2 A2 Vabc.Vane SchatzAún no hay calificaciones
- El Papel de Los Ácidos Grasos de Cadena Corta de La Microbiota Intestinal en La Comunicación IntestinoDocumento6 páginasEl Papel de Los Ácidos Grasos de Cadena Corta de La Microbiota Intestinal en La Comunicación Intestinonest borjasAún no hay calificaciones
- Neurobiología de La AlimentaciónDocumento9 páginasNeurobiología de La AlimentaciónitzelAún no hay calificaciones
- Factores Nucleares de Los NutrientesDocumento16 páginasFactores Nucleares de Los NutrientesRegina GuzmanAún no hay calificaciones
- CronobiologiaDocumento4 páginasCronobiologiaEdwin LopezAún no hay calificaciones
- Así Afecta La Microbiota Al Sistema Nervioso CentralDocumento4 páginasAsí Afecta La Microbiota Al Sistema Nervioso CentralJacqueline AndradeAún no hay calificaciones
- Obesidad: Analisis Etiopatogenico y FisiopatogenicoDocumento8 páginasObesidad: Analisis Etiopatogenico y FisiopatogenicoGustavo Adolfo Aldana PerezAún no hay calificaciones
- 4 - 2012 El Papel de La Mitocondria en El Dolor de La Neuropatía Diabética PDFDocumento8 páginas4 - 2012 El Papel de La Mitocondria en El Dolor de La Neuropatía Diabética PDFc l a u d i a ZAún no hay calificaciones
- Informe de Porque Envejecemos PsicogerontologiaDocumento11 páginasInforme de Porque Envejecemos PsicogerontologiaDmitri JqhehAún no hay calificaciones
- Tarea de Tefi Epigenetica 2020Documento5 páginasTarea de Tefi Epigenetica 2020Estefany Brown GuevaraAún no hay calificaciones
- Entre Discursos - El Cuerpo de La MedicinaDocumento20 páginasEntre Discursos - El Cuerpo de La MedicinaMANDYAún no hay calificaciones
- Neuroquimica de Los Trastornos Bipolares PDFDocumento13 páginasNeuroquimica de Los Trastornos Bipolares PDFgamerhobbistaAún no hay calificaciones
- Rsan 21 2 65Documento6 páginasRsan 21 2 65LuIthurrartAún no hay calificaciones
- Circuito de PapezsDocumento10 páginasCircuito de PapezsJoellmyAún no hay calificaciones
- Terapeutica en Fonoestomatologia TMA-TNA PDFDocumento5 páginasTerapeutica en Fonoestomatologia TMA-TNA PDFMarianela PajonAún no hay calificaciones
- Microbiota IntestinalDocumento7 páginasMicrobiota IntestinalDaisy Jennifer Paulino AriasAún no hay calificaciones
- Fisioterapia en NeurologiaDocumento10 páginasFisioterapia en NeurologiaMichel Berenguela Aravena100% (1)
- Bases Neurobiológ TCADocumento3 páginasBases Neurobiológ TCACarolina PintoAún no hay calificaciones
- Conducta - Motivada Ust PsicologíaDocumento7 páginasConducta - Motivada Ust PsicologíaAmelia Pinto SaavedraAún no hay calificaciones
- 3 MicrobiotaDocumento6 páginas3 MicrobiotaMet LuAún no hay calificaciones
- 6 Articulo Final Sueño AntartidaDocumento14 páginas6 Articulo Final Sueño AntartidaNancyAún no hay calificaciones
- Informe de Psicogastroenterologia de Dinamica de Los Grupos ESTRUCDocumento4 páginasInforme de Psicogastroenterologia de Dinamica de Los Grupos ESTRUCAnderson RamosAún no hay calificaciones
- Pia Articulos de Rev - TSDN - PDFDocumento22 páginasPia Articulos de Rev - TSDN - PDFOla Conde VIIAún no hay calificaciones
- Biologia Del AdipocitoDocumento10 páginasBiologia Del AdipocitoSocorro Gutiérrez de JiménezAún no hay calificaciones
- BioquimicaDocumento2 páginasBioquimicaJatziri OrtizAún no hay calificaciones
- 737 2018 Article 889.en - Es PDFDocumento11 páginas737 2018 Article 889.en - Es PDFConsulta externa Clinica montserratAún no hay calificaciones
- Autismo Equio 123Documento32 páginasAutismo Equio 123Nallely P. IbarraAún no hay calificaciones
- PsiconeuroendocrinologiaDocumento7 páginasPsiconeuroendocrinologiaAlina AntayAún no hay calificaciones
- MIGRAÑADocumento14 páginasMIGRAÑAHernan CappelloAún no hay calificaciones
- Eje Intestino-Cerebro - En.esDocumento15 páginasEje Intestino-Cerebro - En.esRafael Antonio Rosas100% (1)
- NUTRICION EN AUTISMO GrabacionDocumento40 páginasNUTRICION EN AUTISMO Grabacionheverzoncamacaro.praxisAún no hay calificaciones
- Ijms-22-10028 en EsDocumento19 páginasIjms-22-10028 en Esdaniela altamiranoAún no hay calificaciones
- Pa1 - Neurociencias - 2 - Salirrosas Huarniz Jean DiderDocumento6 páginasPa1 - Neurociencias - 2 - Salirrosas Huarniz Jean DiderJEAN DIDIER SALIRROSAS HUARNIZAún no hay calificaciones
- Redalic Paralisis CerebralDocumento10 páginasRedalic Paralisis CerebralLuis Enrique Toledo MuñozAún no hay calificaciones
- Cronobiologia FISIOLOGIADocumento7 páginasCronobiologia FISIOLOGIAPauleth EspinozaAún no hay calificaciones
- Inflamacion en Tea 2019 EspañolDocumento18 páginasInflamacion en Tea 2019 EspañolMaria Gabriela SilvaAún no hay calificaciones
- Estimulación Cerebral No Invasiva Combinada Con Neuroimagen: Hacia Una Medicina de Precisión en El Tratamiento de Las AdiccionesDocumento6 páginasEstimulación Cerebral No Invasiva Combinada Con Neuroimagen: Hacia Una Medicina de Precisión en El Tratamiento de Las AdiccioneslucìaAún no hay calificaciones
- Sindrome de Intestino IrritableDocumento46 páginasSindrome de Intestino IrritableCarolina Noemi Gomez GomezAún no hay calificaciones
- Microbiota Intestinal y Trastornos Del Comportamiento MentalDocumento15 páginasMicrobiota Intestinal y Trastornos Del Comportamiento MentalJesusMonjeAún no hay calificaciones
- Resumen Los Anticuerpos Más Allá de La Defensa Contra MicroorganismosDocumento4 páginasResumen Los Anticuerpos Más Allá de La Defensa Contra MicroorganismosCeli HuarcayaAún no hay calificaciones
- Vias Biológicas Del Impacto de Los Factores Psicosociales Sobre La SaludDocumento19 páginasVias Biológicas Del Impacto de Los Factores Psicosociales Sobre La SaludMauricio CabreraAún no hay calificaciones
- Actualizacion en ObesidadDocumento8 páginasActualizacion en ObesidadadelaidaanguenvemengueAún no hay calificaciones
- EFECTO DE LA APLICACIÓN DE UN PROGRAMA DE BIOFILIA Puerto MaldonadoDocumento20 páginasEFECTO DE LA APLICACIÓN DE UN PROGRAMA DE BIOFILIA Puerto MaldonadoVALENTINA SANTA MARIA ASTUHUAMANAún no hay calificaciones
- Obesidad, Pnie - Trabajo Walter PortalDocumento13 páginasObesidad, Pnie - Trabajo Walter PortalWalter Angel Portal ReateguiAún no hay calificaciones
- Avances Del PnieDocumento43 páginasAvances Del PnieFreeddyy Sthephano Carcasi ChipanaAún no hay calificaciones
- Transtornos Funcionales Del ColonDocumento36 páginasTranstornos Funcionales Del ColonKaren Alexandra Flores GilmayoAún no hay calificaciones
- Epilepsia y La NutriciónDocumento9 páginasEpilepsia y La NutriciónCarinka Vidaños100% (1)
- Metabolismo del sistema digestivo, del hígado, de la vesícula y de las vías biliares: En condiciones de salud y en las enfermedadesDe EverandMetabolismo del sistema digestivo, del hígado, de la vesícula y de las vías biliares: En condiciones de salud y en las enfermedadesAún no hay calificaciones
- Resumen Y Guía De Estudio - La Relación Mente-Intestino: Cómo Impacta La Conversación Secreta, Al Interior De Nuestros Cuerpos En Nuestro Animo, Nuestras Elecciones Y Nuestra Salud En GeneralDe EverandResumen Y Guía De Estudio - La Relación Mente-Intestino: Cómo Impacta La Conversación Secreta, Al Interior De Nuestros Cuerpos En Nuestro Animo, Nuestras Elecciones Y Nuestra Salud En GeneralAún no hay calificaciones
- El Sistema CardiovascularDocumento22 páginasEl Sistema CardiovascularEstefanny ForeroAún no hay calificaciones
- Embriología y Genética 1ro Tema2Documento62 páginasEmbriología y Genética 1ro Tema2Daniela MirandaAún no hay calificaciones
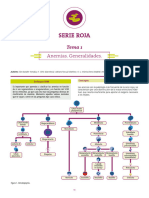
- Hematología AMIR 14va Edición-13-36Documento24 páginasHematología AMIR 14va Edición-13-36Mauro VelascoAún no hay calificaciones
- Anatomía Semana 1 PDFDocumento5 páginasAnatomía Semana 1 PDFMaria Paredes PeresAún no hay calificaciones
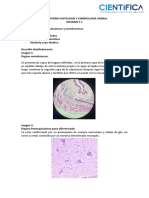
- Informes 5 - 9 (Guia Lab 2) .Docx - CompressedDocumento11 páginasInformes 5 - 9 (Guia Lab 2) .Docx - CompressedalexaAún no hay calificaciones
- Juarroz Poesia 12Documento24 páginasJuarroz Poesia 12León PedrouzoAún no hay calificaciones
- Glosario de Términos PDFDocumento5 páginasGlosario de Términos PDFLeandro MateoAún no hay calificaciones
- Cuadro ResumenDocumento1 páginaCuadro ResumenNATALIA ARIANA ALBUJAR RODRIGUEZAún no hay calificaciones
- El Intestino El Segundo CerebroDocumento5 páginasEl Intestino El Segundo CerebroMartha_ArAún no hay calificaciones
- Block Normado 19x19x39Documento1 páginaBlock Normado 19x19x39RickeyLopezAún no hay calificaciones
- Circulación en El HombreDocumento3 páginasCirculación en El Hombrepaula DelgadoAún no hay calificaciones
- ROTACION II-EXAMEN TEÓRICO PEDIATRIA-respuestasDocumento4 páginasROTACION II-EXAMEN TEÓRICO PEDIATRIA-respuestasAngel CedeñoAún no hay calificaciones
- Qué Es La SensopercepciónDocumento12 páginasQué Es La SensopercepciónMaricarmen AguilarAún no hay calificaciones
- 07 Anabolizantes Beneficios e MaleficiosDocumento2 páginas07 Anabolizantes Beneficios e MaleficiosKlysmann ReisAún no hay calificaciones
- Línea de Tiempo Del Periodo FetalDocumento1 páginaLínea de Tiempo Del Periodo FetalAdriana BibAún no hay calificaciones
- El Sistema CirculatorioDocumento4 páginasEl Sistema CirculatoriomgarciacalasanzAún no hay calificaciones
- Guía 2-CN-701,702-Sistema muscular-Yery-Período 3Documento4 páginasGuía 2-CN-701,702-Sistema muscular-Yery-Período 3ybar09121973Aún no hay calificaciones
- Clasificacion de LesionesDocumento48 páginasClasificacion de LesionesVanessa CruZzAún no hay calificaciones
- SISTEMA OSEO 3ero PDFDocumento144 páginasSISTEMA OSEO 3ero PDFSebastian EspinozaAún no hay calificaciones
- Todo Sobre El Cerebro HumanoDocumento2 páginasTodo Sobre El Cerebro HumanoESTEFANIAún no hay calificaciones
- Eje, Hipotálamo, Hipófisis OvariotestículoDocumento14 páginasEje, Hipotálamo, Hipófisis Ovariotestículoconrado.angel11Aún no hay calificaciones
- Sistema NerviosoDocumento5 páginasSistema NerviosoRuben Carlos Molina OlavarriaAún no hay calificaciones
- Solemne 2 Biología CelularDocumento5 páginasSolemne 2 Biología CelularVictoria BriceñoAún no hay calificaciones
- Sesión 14 y 15 Direccionamiento ProteicoDocumento60 páginasSesión 14 y 15 Direccionamiento ProteicoAnai ValenciaAún no hay calificaciones
- TERMORREGULACIÓNDocumento4 páginasTERMORREGULACIÓNSanti PalomequeAún no hay calificaciones
- ManualDocumento15 páginasManualyessica edith villanueva floresAún no hay calificaciones
- 2do Grado Examen DiciembreDocumento12 páginas2do Grado Examen DiciembrepepeAún no hay calificaciones