También podría gustarte
- Recetario Tradicional VenezolanoDocumento62 páginasRecetario Tradicional Venezolanobrujie100% (1)
- La importancia de la nutrición en el paciente oncológicoDe EverandLa importancia de la nutrición en el paciente oncológicoAún no hay calificaciones
- Bases Moleculares Del CancerDocumento26 páginasBases Moleculares Del CancerAlan MendozaAún no hay calificaciones
- CarcinogénesisDocumento30 páginasCarcinogénesisMauricioKiraAún no hay calificaciones
- ONCOGENESDocumento93 páginasONCOGENESMarco Herrera Flores100% (1)
- Neoplasias Bases MolecularesDocumento27 páginasNeoplasias Bases MolecularesMaria Del Carmen Morante BallesterAún no hay calificaciones
- ONCOGENETICADocumento52 páginasONCOGENETICAKitleng TayAún no hay calificaciones
- Bases Moleculares Del CancerDocumento9 páginasBases Moleculares Del CancertogetaAún no hay calificaciones
- Absorción con filtro BK como terapia eficaz en el riñón del mieloma múltipleDe EverandAbsorción con filtro BK como terapia eficaz en el riñón del mieloma múltipleAún no hay calificaciones
- Uso de Derivados del Cannabis en Pacientes con CáncerDe EverandUso de Derivados del Cannabis en Pacientes con CáncerAún no hay calificaciones
- Bases CarcinogenesisDocumento75 páginasBases Carcinogenesiscit.valdez100% (9)
- Ejecuciones colectivasDocumento6 páginasEjecuciones colectivasFernando José Hernández Afre100% (1)
- ALTERACIONESDocumento18 páginasALTERACIONESChipiRosalesAún no hay calificaciones
- Genética Del Cáncer - Grupo 4Documento18 páginasGenética Del Cáncer - Grupo 4Emely RoussAún no hay calificaciones
- Genes Supresores de CancerDocumento3 páginasGenes Supresores de CancerCristina CachiAún no hay calificaciones
- Insensibilidad A La InhibicionDocumento27 páginasInsensibilidad A La Inhibicion2020 Barranco Hernandez Jesús JavierAún no hay calificaciones
- Introduccion A La Oncologia PediatricaDocumento3 páginasIntroduccion A La Oncologia PediatricaIsabel Soto AAún no hay calificaciones
- Insensibilidad A La Inhibición Del Crecimiento y EvasiónDocumento34 páginasInsensibilidad A La Inhibición Del Crecimiento y EvasiónMarita Mirelle Solis Rivero0% (1)
- Gene SsuresoresDocumento11 páginasGene SsuresoresMarcela MontillaAún no hay calificaciones
- Genes Supresores Tumorales y Oncogenes ArchivoDocumento28 páginasGenes Supresores Tumorales y Oncogenes ArchivoMiguelon OviedoAún no hay calificaciones
- Genética y Genómica Del CáncerDocumento39 páginasGenética y Genómica Del CáncerBryan JacomeAún no hay calificaciones
- Resumengeneticadelcancer VeraDocumento4 páginasResumengeneticadelcancer VeraJuan Jose VAAún no hay calificaciones
- Gen RBDocumento2 páginasGen RBOscar Leones GuerraAún no hay calificaciones
- 10 pasos claves de la CarcinogénesisDocumento10 páginas10 pasos claves de la CarcinogénesisLina SantiagoAún no hay calificaciones
- Alteraciones Celulares y Enfermedad PDFDocumento85 páginasAlteraciones Celulares y Enfermedad PDFVicky Stefany Montoya PachecoAún no hay calificaciones
- Genes Supresores Tumorales y Su Influencia en El Ciclo CelularDocumento2 páginasGenes Supresores Tumorales y Su Influencia en El Ciclo CelularLucilaAún no hay calificaciones
- Protooncogenes y OncogenesDocumento6 páginasProtooncogenes y OncogenessanchezcoronaAún no hay calificaciones
- Oncogenes y El CancerDocumento10 páginasOncogenes y El CancerCora AgüeroAún no hay calificaciones
- Genes Relacionados Con CáncerDocumento3 páginasGenes Relacionados Con CáncerJosé Antonio Villicaña ZúñigaAún no hay calificaciones
- Sem9 ONCOGENES Y CANCER-Carlo CroceDocumento9 páginasSem9 ONCOGENES Y CANCER-Carlo CroceRoger santa cruz zamoraAún no hay calificaciones
- Características y alteraciones del cáncerDocumento4 páginasCaracterísticas y alteraciones del cáncerNORBERTO VICTORINOAún no hay calificaciones
- Genes 1Documento8 páginasGenes 1Luisa MendozaAún no hay calificaciones
- CancerDocumento7 páginasCancerTania Elizabeth ToledanoAún no hay calificaciones
- Bases Moleculares Del CáncerDocumento32 páginasBases Moleculares Del CáncerJudit LopeAún no hay calificaciones
- Metabolismo de Las Celulas NeoplasicasDocumento4 páginasMetabolismo de Las Celulas NeoplasicasAannaxeAún no hay calificaciones
- Bases moleculares cáncer oncogenesDocumento18 páginasBases moleculares cáncer oncogenessamanthaAún no hay calificaciones
- OncogenesDocumento26 páginasOncogenesChoche Luis JodonAún no hay calificaciones
- Ud6 - Citogenética y CancerDocumento9 páginasUd6 - Citogenética y Cancerarino.martinez.monicaaAún no hay calificaciones
- Ciclo Celular y CancerDocumento5 páginasCiclo Celular y CancerAnette LunaAún no hay calificaciones
- CancerDocumento10 páginasCancerAndrik HernandezAún no hay calificaciones
- Clase 10. Agentes Carcinogenicos y ProtooncogenesDocumento85 páginasClase 10. Agentes Carcinogenicos y ProtooncogenesUSMP FN ARCHIVOSAún no hay calificaciones
- Genes Supresores de TumoresDocumento7 páginasGenes Supresores de TumoresJonathan Álvarez AndradeAún no hay calificaciones
- B37. Bioquímica Del CáncerDocumento18 páginasB37. Bioquímica Del CáncerMatilde Antero Mercado AlemanAún no hay calificaciones
- Genes Supresores TumoresDocumento4 páginasGenes Supresores TumoresJesus Yampier Diaz ZeladaAún no hay calificaciones
- Genes y Cáncer IIDocumento15 páginasGenes y Cáncer IIHernán UrbanoAún no hay calificaciones
- Los Genes Del Cancer 2-2Documento36 páginasLos Genes Del Cancer 2-2Sofia CillisAún no hay calificaciones
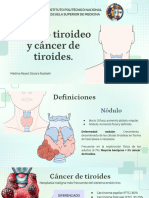
- Nódulo Tiroideo y Cáncer de TiroidesDocumento15 páginasNódulo Tiroideo y Cáncer de Tiroidesreymed.385Aún no hay calificaciones
- Cancer Del Sistema PulmonarDocumento33 páginasCancer Del Sistema Pulmonar000035995Aún no hay calificaciones
- Inmunidad frente a tumoresDocumento4 páginasInmunidad frente a tumoresOctavio ZmAún no hay calificaciones
- Inmunidad Frente A Los TumoresDocumento4 páginasInmunidad Frente A Los TumoresJacs SaulAún no hay calificaciones
- Genes y Cáncer: Vol. 11 N°2 Abril 2000Documento6 páginasGenes y Cáncer: Vol. 11 N°2 Abril 2000abaurreangelaAún no hay calificaciones
- Agentes Carcinogenicos y Protooncogenes 2014Documento78 páginasAgentes Carcinogenicos y Protooncogenes 2014Fatima Flores VelasquezAún no hay calificaciones
- PROTOONCOGENESDocumento6 páginasPROTOONCOGENESMalu MartinezAún no hay calificaciones
- Informe Seminario 22Documento21 páginasInforme Seminario 22VPA IFMSA UNTAún no hay calificaciones
- Examen Seminario CancerDocumento6 páginasExamen Seminario CancerMarbel Maydana MendietaAún no hay calificaciones
- Cáncer de CólonDocumento6 páginasCáncer de CólonAlexis RodriguezAún no hay calificaciones
- Cáncer Biología Molecular Ciclo Celular GenesDocumento2 páginasCáncer Biología Molecular Ciclo Celular GenesLulu CortésAún no hay calificaciones
- CÁNCER DE PRÓSTATADocumento69 páginasCÁNCER DE PRÓSTATAslipknotfan9613Aún no hay calificaciones
- Ciclo CelularDocumento26 páginasCiclo CelularMichelle Rocio Carvajal LaraAún no hay calificaciones
- Taller 2 VPH Y VHSDocumento10 páginasTaller 2 VPH Y VHSKaritho MontesAún no hay calificaciones
- Parte 2: Bases Moleculares Del Cáncer Muy Controlada de Eventos. Estos Eventos Dependen de Niveles Adecuados de Transcripción y Traducción deDocumento3 páginasParte 2: Bases Moleculares Del Cáncer Muy Controlada de Eventos. Estos Eventos Dependen de Niveles Adecuados de Transcripción y Traducción deMaria JoseAún no hay calificaciones
- PRB y CáncerDocumento10 páginasPRB y CáncerjoseluisolveracalderonAún no hay calificaciones
- SESION 14 Búsqueda Avanzada de Informacion en InternetDocumento33 páginasSESION 14 Búsqueda Avanzada de Informacion en InternetALIS PAMELA GUIVAR CHAMBERGOAún no hay calificaciones
- Foro Relacion Medico PacienteDocumento1 páginaForo Relacion Medico PacienteALIS PAMELA GUIVAR CHAMBERGOAún no hay calificaciones
- Clase Sem 10Documento2 páginasClase Sem 10ALIS PAMELA GUIVAR CHAMBERGOAún no hay calificaciones
- Clase Sem 10Documento3 páginasClase Sem 10ALIS PAMELA GUIVAR CHAMBERGOAún no hay calificaciones
- Guia A Colores Turistica de AmsterdamDocumento64 páginasGuia A Colores Turistica de AmsterdamhectorAún no hay calificaciones
- Resolucion Ministerial #483-2000-MTC-15.04Documento2 páginasResolucion Ministerial #483-2000-MTC-15.04mercedessalazarAún no hay calificaciones
- Introduccion Riddles of Existence EspanolDocumento2 páginasIntroduccion Riddles of Existence EspanolNatalia Luna LunaAún no hay calificaciones
- Reglamento General Campeonato Argentino de Pilotos. CAF ACTC - 2018Documento38 páginasReglamento General Campeonato Argentino de Pilotos. CAF ACTC - 2018DiegoAún no hay calificaciones
- Escritos para La Mesa de Fe - ROLY - Revision JDDocumento36 páginasEscritos para La Mesa de Fe - ROLY - Revision JDJuan Daniel De La Vega ValenzuelaAún no hay calificaciones
- Tesis 4-FranklimDocumento266 páginasTesis 4-FranklimLuis Lopez GonzalesAún no hay calificaciones
- Unidad 1 Estructura Del Cuerpo HumanoDocumento125 páginasUnidad 1 Estructura Del Cuerpo HumanoLida Yaneth Quimbayo VargasAún no hay calificaciones
- Satir, Virginia - Curso de Programación PNL en 4 LeccionesDocumento6 páginasSatir, Virginia - Curso de Programación PNL en 4 LeccionesAngelo FaggioliAún no hay calificaciones
- Derecho Empresarial EnsayoDocumento4 páginasDerecho Empresarial EnsayoIngrid Ipanaqué LamasAún no hay calificaciones
- Enfermeria ComunitariaDocumento201 páginasEnfermeria Comunitariamilus100100% (1)
- Visita A La Granja Avicola SantiagoDocumento6 páginasVisita A La Granja Avicola SantiagoYeni ArratiaAún no hay calificaciones
- Matemática Financiera Aspectos BásicosDocumento26 páginasMatemática Financiera Aspectos BásicosAbigail Jara VelaAún no hay calificaciones
- Tema 8 FisicaDocumento33 páginasTema 8 FisicaHeriberto Gonzalez BritoAún no hay calificaciones
- Carrera de Tecnico Superior en Enfermería Prácticas Pre-Profesionales 17 de Abril Al 31 de Mayo Del 2023Documento3 páginasCarrera de Tecnico Superior en Enfermería Prácticas Pre-Profesionales 17 de Abril Al 31 de Mayo Del 2023Nicolas BeltranAún no hay calificaciones
- Resumen Sach Cap 9Documento4 páginasResumen Sach Cap 9Angela Macarena Cerda GonzálezAún no hay calificaciones
- Pavimentos de Concreto Simple Con PasadoresDocumento8 páginasPavimentos de Concreto Simple Con PasadoresJorge LugoAún no hay calificaciones
- Algebra lineal taller 2Documento5 páginasAlgebra lineal taller 2ZACKS HOLOSAún no hay calificaciones
- Metabolismo de Los Carbohidratos, Proteinas y LipidosDocumento18 páginasMetabolismo de Los Carbohidratos, Proteinas y LipidosLESLI CAROL MAMANI HUMPIRIAún no hay calificaciones
- Metabolismo GlucidosDocumento25 páginasMetabolismo GlucidosJulian PérezAún no hay calificaciones
- Relato Individual de ExperienciaDocumento4 páginasRelato Individual de ExperienciafernandoAún no hay calificaciones
- Historia de La Topografia PDFDocumento13 páginasHistoria de La Topografia PDFMishell Angueta GiraldoAún no hay calificaciones
- 9-Informe de Laboratorio - Movimiento Armónico SimpleDocumento5 páginas9-Informe de Laboratorio - Movimiento Armónico Simpleandrea delgadoAún no hay calificaciones
- Marco Teórico Referencial MicroEmpresasDocumento9 páginasMarco Teórico Referencial MicroEmpresasHugo Tasbille0% (1)
- Agua Con Limon en El Embarazo - Buscar Con GoogleDocumento1 páginaAgua Con Limon en El Embarazo - Buscar Con GoogleSyrailiy SilatanAún no hay calificaciones
- Agilidad de Liderazgo Joiner Es 22719Documento7 páginasAgilidad de Liderazgo Joiner Es 22719columna74Aún no hay calificaciones
- HV Omar de La Cruz 2022Documento5 páginasHV Omar de La Cruz 2022LuisitoAún no hay calificaciones
- Cadena Respiratoria MitocondrialDocumento7 páginasCadena Respiratoria Mitocondrialana guadalupe rodriguez ariasAún no hay calificaciones