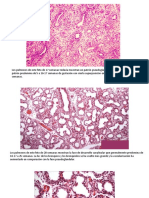
También podría gustarte
- Embarazo, parto y lactancia del bebéDe EverandEmbarazo, parto y lactancia del bebéAún no hay calificaciones
- Cuestionario 2do Parcial EmbrioDocumento22 páginasCuestionario 2do Parcial EmbrioDiana CastilloAún no hay calificaciones
- Informe 10 de EmbriologiaDocumento8 páginasInforme 10 de EmbriologiaYessenia Tasilla Huaripata100% (1)
- Sistema RespiratorioDocumento84 páginasSistema RespiratorioAntony Varela MontielAún no hay calificaciones
- Cavidades Corporales Mesenterios y DiafragmaDocumento51 páginasCavidades Corporales Mesenterios y Diafragmacaleb apaza aruquipa100% (1)
- Desarrollo de Las Cavidades CorcoporalesDocumento33 páginasDesarrollo de Las Cavidades CorcoporalesDiego IsaacAún no hay calificaciones
- Cavidades CorporalesDocumento4 páginasCavidades Corporales2003paolajimenezAún no hay calificaciones
- CAPITULO 8 - Cavidades Corporales, Mesenterios y DiafragmaDocumento3 páginasCAPITULO 8 - Cavidades Corporales, Mesenterios y DiafragmaTodoMePuedeAlvAún no hay calificaciones
- CAPITULO 8 - Cavidades Corporales, Mesenterios y DiafragmaDocumento4 páginasCAPITULO 8 - Cavidades Corporales, Mesenterios y DiafragmaTodoMePuedeAlvAún no hay calificaciones
- CELOMADocumento27 páginasCELOMAFrancinete Gomes de AraújoAún no hay calificaciones
- Desarrollo de las cavidades corporales embrionariasDocumento43 páginasDesarrollo de las cavidades corporales embrionariascarlosmpm4604Aún no hay calificaciones
- Cavidades CorporalesDocumento3 páginasCavidades CorporalesSara Lucía AchoyAún no hay calificaciones
- Resumen Tubo Intestinal y Las Cavidades Corporales 1Documento5 páginasResumen Tubo Intestinal y Las Cavidades Corporales 1Aidaly Carolina Viera Ponce De LeónAún no hay calificaciones
- CAPITULO 7 ResumenDocumento5 páginasCAPITULO 7 ResumenMiriam Martinez LópezAún no hay calificaciones
- Cavidades CorporalesDocumento18 páginasCavidades Corporalesperliux1406Aún no hay calificaciones
- Diafragma y Cavidad TorácicaDocumento48 páginasDiafragma y Cavidad TorácicaUlises García ArellanoAún no hay calificaciones
- Cavidades Corporales, Mesenterio y DiafragmaDocumento7 páginasCavidades Corporales, Mesenterio y DiafragmaMARK TORREZ DEHEZAAún no hay calificaciones
- Cavidades Corporales Mesenterios y DiafragmaDocumento34 páginasCavidades Corporales Mesenterios y DiafragmaGraciela SaénzAún no hay calificaciones
- Resumen EmbriologíaDocumento6 páginasResumen EmbriologíaKatherine Manriquez TrenadoAún no hay calificaciones
- Cavidades CorporalesDocumento4 páginasCavidades CorporalesLana KluyAún no hay calificaciones
- Musculatura Del TroncoDocumento26 páginasMusculatura Del TroncoNatacha QuevedoAún no hay calificaciones
- Cavidades CorporalesDocumento9 páginasCavidades CorporalesBelen Yapu100% (1)
- Cavidades Corporales PDFDocumento3 páginasCavidades Corporales PDFcaleb apaza aruquipaAún no hay calificaciones
- Clase 10. Formacion de Cavidades CorporalesDocumento36 páginasClase 10. Formacion de Cavidades CorporalesClaudia IquizeAún no hay calificaciones
- Cavidades Corporales, Mesenterios y DiafragmaDocumento51 páginasCavidades Corporales, Mesenterios y DiafragmaKaren Hernàndez67% (3)
- Embriología del diafragma y las cavidades corporalesDocumento20 páginasEmbriología del diafragma y las cavidades corporalesNatalia MendezAún no hay calificaciones
- Formación de las cavidades corporales y membranas serosasDocumento31 páginasFormación de las cavidades corporales y membranas serosasJhonatan Rubio GarcíaAún no hay calificaciones
- Desarrollo cavidad corporal embrionariaDocumento3 páginasDesarrollo cavidad corporal embrionariaCristian CordovaAún no hay calificaciones
- EMBRIOLOGÍA CAVIDADES CORPORALES y RESPIRATORIODocumento13 páginasEMBRIOLOGÍA CAVIDADES CORPORALES y RESPIRATORIOannabella del vecchioAún no hay calificaciones
- Cavidades EmbrionariasDocumento41 páginasCavidades EmbrionariasMaria ReyesAún no hay calificaciones
- MESENTERIOSDocumento2 páginasMESENTERIOSJorge Cornelio Mainato ShagñayAún no hay calificaciones
- 1 Cavidades Corporales y OsteomuscularDocumento48 páginas1 Cavidades Corporales y OsteomuscularShalom ApérrigueAún no hay calificaciones
- El Tubo Intestinal y Las Cavidades CorporalesDocumento22 páginasEl Tubo Intestinal y Las Cavidades CorporalesMarcos DantasAún no hay calificaciones
- Cavidadescorporalesmesenteriosydiafragma 110609002529 Phpapp02Documento34 páginasCavidadescorporalesmesenteriosydiafragma 110609002529 Phpapp02Ale Alexandra Yanque MuñozAún no hay calificaciones
- Mesos, Celoma y sus anomalíasDocumento5 páginasMesos, Celoma y sus anomalíasAdolfo Antonio Ríos AlcortaAún no hay calificaciones
- Desarrollo de cavidades corporales y defectos de paredDocumento104 páginasDesarrollo de cavidades corporales y defectos de paredMateo ChurtaAún no hay calificaciones
- Cap. 7 El Tubo Intestinal y Las Cavidades CorporalesDocumento15 páginasCap. 7 El Tubo Intestinal y Las Cavidades CorporalesGuido Walter Vargas ApazaAún no hay calificaciones
- Cavidades, Diafragma y MesenterioDocumento7 páginasCavidades, Diafragma y MesenterioLuz Moscoso Aguirre100% (1)
- El Tubo Intestinal y Las Cavidades Corporales..Documento5 páginasEl Tubo Intestinal y Las Cavidades Corporales..Denilson Mena Tito ZabalaAún no hay calificaciones
- CavidadesDocumento3 páginasCavidadesMayrisAún no hay calificaciones
- Las Cavidades CorporalesDocumento7 páginasLas Cavidades CorporalesShia PinoAún no hay calificaciones
- Resumen de Sistema Digestivo Embriologia de Langman !!!!Documento6 páginasResumen de Sistema Digestivo Embriologia de Langman !!!!Cedrid MoralesAún no hay calificaciones
- Tema 13 - Cavidades Corporales y SerosasDocumento9 páginasTema 13 - Cavidades Corporales y SerosasAdriana Izquierdo CalvoAún no hay calificaciones
- Cavidades Corporales Mesenterios y DiafragmaDocumento27 páginasCavidades Corporales Mesenterios y DiafragmaMaria MartinezAún no hay calificaciones
- Embriología 13 A Las Cavidades CorporalesDocumento51 páginasEmbriología 13 A Las Cavidades CorporalesCARMITA ESPERANZA VALDIVIEZO PORTOCARREROAún no hay calificaciones
- Cavidadescorporalesmesenteriosydiafragma 110609002529 Phpapp02Documento34 páginasCavidadescorporalesmesenteriosydiafragma 110609002529 Phpapp02Rodrigo Randy Aliaga HuancaAún no hay calificaciones
- Cavidad Corporal, Mesentericos y DiafragmaDocumento12 páginasCavidad Corporal, Mesentericos y DiafragmaenriqueAún no hay calificaciones
- Tubo DigestivoDocumento56 páginasTubo DigestivoEmiliano CardenasAún no hay calificaciones
- Morfología Humana Con Orientación Clínica IIDocumento25 páginasMorfología Humana Con Orientación Clínica IIJosé CarlosAún no hay calificaciones
- Embriología cavidadesDocumento19 páginasEmbriología cavidadesDavenia SaldañaAún no hay calificaciones
- 10 - Cavidades CelomáticasDocumento4 páginas10 - Cavidades Celomáticasbio.agus92Aún no hay calificaciones
- Embriología de las mamas, pulmones y corazónDocumento53 páginasEmbriología de las mamas, pulmones y corazónAnggie Lissette García ViciosoAún no hay calificaciones
- Desarrollo de Las Cavidades CorporalesDocumento5 páginasDesarrollo de Las Cavidades CorporalesAlicia Hernández PérezAún no hay calificaciones
- Aparato Digestivo 2 Embrio 2023Documento4 páginasAparato Digestivo 2 Embrio 2023RaoulAún no hay calificaciones
- Anatomia y Embriologia Del IntestinoDocumento31 páginasAnatomia y Embriologia Del IntestinoCamilo Tiu GrijalvaAún no hay calificaciones
- Aparato DigestivoDocumento10 páginasAparato DigestivoJoyce ZambranoAún no hay calificaciones
- Cavidades Del Cuerpo HumanoDocumento1 páginaCavidades Del Cuerpo HumanoJIMMY RICARDO PAZMIÑO ARROBAAún no hay calificaciones
- Formación del diafragma y cavidades corporalesDocumento48 páginasFormación del diafragma y cavidades corporalesUlises García Arellano100% (7)
- Anatomia Generalidades de EsplacnologiaDocumento6 páginasAnatomia Generalidades de EsplacnologiaDiana RengelAún no hay calificaciones
- Estomago 100112164159 Phpapp01Documento23 páginasEstomago 100112164159 Phpapp01Moreno MoralesAún no hay calificaciones
- Capitulo 7 - Langman - Cavidades CorporalesDocumento3 páginasCapitulo 7 - Langman - Cavidades Corporalesmaria jose sanchez gilAún no hay calificaciones
- RESUMEN de Juan GaviotaDocumento6 páginasRESUMEN de Juan GaviotaRuth Simón VergaraAún no hay calificaciones
- Afectividad y PercepcionDocumento59 páginasAfectividad y PercepcionRuth Simón VergaraAún no hay calificaciones
- 3ra Clase ppt3-1Documento31 páginas3ra Clase ppt3-1Ruth Simón VergaraAún no hay calificaciones
- Modelo - ArgumentoDocumento2 páginasModelo - ArgumentoRuth Simón VergaraAún no hay calificaciones
- Uso de AlcanosDocumento6 páginasUso de AlcanosRuth Simón Vergara100% (1)
- EstereoquímicaDocumento32 páginasEstereoquímicaRuth Simón VergaraAún no hay calificaciones
- Entalpía reaccionesDocumento6 páginasEntalpía reaccionesRuth Simón VergaraAún no hay calificaciones
- 4 - 4.2 EntalpiaDocumento3 páginas4 - 4.2 EntalpiaRuth Simón VergaraAún no hay calificaciones
- Ley Que Modifica La Ley 27972 Ley Organica de Municipalid Ley N 31079 1907435 2Documento3 páginasLey Que Modifica La Ley 27972 Ley Organica de Municipalid Ley N 31079 1907435 2Ruth Simón VergaraAún no hay calificaciones
- Entalpía reaccionesDocumento6 páginasEntalpía reaccionesRuth Simón VergaraAún no hay calificaciones
- Flora Es El Conjunto de Las Plantas Que Pueblan Un HábitatDocumento2 páginasFlora Es El Conjunto de Las Plantas Que Pueblan Un HábitatRuth Simón VergaraAún no hay calificaciones
- Experiencias de Aprendizaje 3er GradoDocumento67 páginasExperiencias de Aprendizaje 3er GradoAlejandroRamirez60% (5)
- Antomia y Fisiologia de La VentilacionDocumento36 páginasAntomia y Fisiologia de La VentilacionSaeed DiazAún no hay calificaciones
- Respiratorio IiDocumento31 páginasRespiratorio IianonymoAún no hay calificaciones
- Paramixovirus: agentes causantes de enfermedades respiratoriasDocumento65 páginasParamixovirus: agentes causantes de enfermedades respiratoriasJANAINA DA SILVA LIMAAún no hay calificaciones
- Copia de Copia de DradioDocumento11 páginasCopia de Copia de DradioLuis IguaranAún no hay calificaciones
- Desarrollo Embrionario Del Sistema Respiratorio - Línea Del TiempoDocumento5 páginasDesarrollo Embrionario Del Sistema Respiratorio - Línea Del TiempoSalma LaraAún no hay calificaciones
- Glosario Histología FinalDocumento13 páginasGlosario Histología FinalDanai GonzalezAún no hay calificaciones
- 3ro de SecundariaDocumento13 páginas3ro de SecundariaCarlos Ramiro Uriona TapiaAún no hay calificaciones
- Informe 09 - Morales - BrendaDocumento17 páginasInforme 09 - Morales - BrendaKaterin HilarioAún no hay calificaciones
- Practica 1 PulmonDocumento18 páginasPractica 1 PulmonLizeth Rejas MendozaAún no hay calificaciones
- Semio Radiología - Trabajo GrupalDocumento12 páginasSemio Radiología - Trabajo Grupaljohn moraAún no hay calificaciones
- Sistema RespiratorioDocumento33 páginasSistema RespiratorioBryan PozoAún no hay calificaciones
- Informe Histología S.respiratorio.Documento7 páginasInforme Histología S.respiratorio.Katalina Acevedo DiazAún no hay calificaciones
- Vias Aereas Sup e InfDocumento16 páginasVias Aereas Sup e InfDaniel EscobarAún no hay calificaciones
- Respirator I oDocumento12 páginasRespirator I oRené MuñozAún no hay calificaciones
- Apuntes TC Torax y CerebroDocumento50 páginasApuntes TC Torax y CerebroYami SeguinAún no hay calificaciones
- Cap 1 Modulo 1Documento26 páginasCap 1 Modulo 1emycatahanaAún no hay calificaciones
- Histo - 2° CuatrimestreDocumento158 páginasHisto - 2° CuatrimestremaruorioloAún no hay calificaciones
- El Sistema Digestivo y RespiratorioDocumento11 páginasEl Sistema Digestivo y RespiratorioJovanny Tolentino RamosAún no hay calificaciones
- Aparato RespiratorioDocumento20 páginasAparato RespiratorioCHRISTIAN MATIAS GARCIAAún no hay calificaciones
- GUÍA DE HISTOEMBRIOLOGÍA 2023 Práctica 7 Piel, Cardiovascular, RespiratorioDocumento8 páginasGUÍA DE HISTOEMBRIOLOGÍA 2023 Práctica 7 Piel, Cardiovascular, RespiratorioalbieriaqperuAún no hay calificaciones
- Aparato Respiratorio en La Ifancia BuenoDocumento13 páginasAparato Respiratorio en La Ifancia BuenoYesica QuintanaAún no hay calificaciones
- Patología Animal II: Lina M Palomino V MVZ, MSCDocumento354 páginasPatología Animal II: Lina M Palomino V MVZ, MSCGeraldine sanchezAún no hay calificaciones
- Manual de Temas de Charla de 10 Minutos Enero - 2020Documento34 páginasManual de Temas de Charla de 10 Minutos Enero - 2020joseluisescalanteAún no hay calificaciones
- Histología Pulmón PROYECTODocumento6 páginasHistología Pulmón PROYECTOGerardo PicazoAún no hay calificaciones
- Ada 4 Diseccion - Equipo2Documento32 páginasAda 4 Diseccion - Equipo2Yuliani PerezAún no hay calificaciones
- Lìnea Del Tiempo - Sistema Respiratorio - PràcticaDocumento37 páginasLìnea Del Tiempo - Sistema Respiratorio - PràcticaMatha Janet Cruz RiosAún no hay calificaciones
- Repaso 2do SemestreDocumento15 páginasRepaso 2do Semestremariana ruvaAún no hay calificaciones
- Sistema Respiratorio MioDocumento92 páginasSistema Respiratorio MioRoberto UcAún no hay calificaciones
- 5 Cie PL CTDocumento21 páginas5 Cie PL CTÁMBAR ANTONELLA JIMÉNEZ MORELAún no hay calificaciones