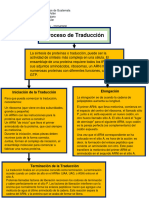
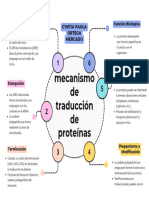
También podría gustarte
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- R T23 TraduccionDocumento3 páginasR T23 TraduccionGeraldine BordaAún no hay calificaciones
- Guia Repaso BiologiaDocumento10 páginasGuia Repaso BiologiaDiego VillegasAún no hay calificaciones
- CitosolDocumento15 páginasCitosolAlba Macipe FeitoAún no hay calificaciones
- TEMA 11 Transcripción y TraducciónDocumento8 páginasTEMA 11 Transcripción y TraducciónLAURA LARAAún no hay calificaciones
- Biosíntesis de ProteínasDocumento15 páginasBiosíntesis de Proteínasgian diaz olazoAún no hay calificaciones
- Bioquímica Traduccion GeneticaDocumento4 páginasBioquímica Traduccion GeneticaRafael ColmenaresAún no hay calificaciones
- TraduccionDocumento1 páginaTraduccionLiz CastellanosAún no hay calificaciones
- Taller 3 HECHO PDFDocumento7 páginasTaller 3 HECHO PDFPascual FerrerAún no hay calificaciones
- PROTEINADocumento11 páginasPROTEINAAlexandra PazAún no hay calificaciones
- REPLICACIÓNDocumento7 páginasREPLICACIÓNAlfonzo CabreraAún no hay calificaciones
- 25) Código GenéticoDocumento1 página25) Código Genéticoandrea mejia leonAún no hay calificaciones
- TraducciónDocumento9 páginasTraducciónPaula Estefania Urrego VillalobosAún no hay calificaciones
- Sesion 3 Sintesis de ProteinasDocumento26 páginasSesion 3 Sintesis de ProteinasMathiasAronMartinezTellesAún no hay calificaciones
- C7. Proteinas y Traducción 2Documento2 páginasC7. Proteinas y Traducción 2ELISA TABOADAAún no hay calificaciones
- Genetica IIIDocumento7 páginasGenetica IIIClaudia Lopez VicenteAún no hay calificaciones
- Mecanismo de Traduccion de ProteinasDocumento1 páginaMecanismo de Traduccion de ProteinasNocturnal PhotosyntesisAún no hay calificaciones
- Wuolah-Premium-Tema 6 - La Traducción PDFDocumento24 páginasWuolah-Premium-Tema 6 - La Traducción PDFISALEAún no hay calificaciones
- TEMA 7 Biología FarmaciaDocumento6 páginasTEMA 7 Biología FarmaciasaraAún no hay calificaciones
- Ácidos Nucleicos 2 21042022Documento31 páginasÁcidos Nucleicos 2 21042022Vicky Olivero MengoAún no hay calificaciones
- Cuestionario Traducción, MutagénesisDocumento3 páginasCuestionario Traducción, MutagénesisZuri ManriqueAún no hay calificaciones
- Taller N°9 Transcripcion y Traduccion (RESPONDIDO)Documento4 páginasTaller N°9 Transcripcion y Traduccion (RESPONDIDO)Inés Berlín OssandónAún no hay calificaciones
- Actividad Traducción y Transcripcion AdnDocumento5 páginasActividad Traducción y Transcripcion AdnMalena CrowAún no hay calificaciones
- Sintesis Proteinas - B3 - 2022Documento6 páginasSintesis Proteinas - B3 - 2022Vale RyAún no hay calificaciones
- Clase S13 Transcripcion-TraduccionDocumento26 páginasClase S13 Transcripcion-Traduccionmanuel ramirezAún no hay calificaciones
- REGULACIÓNDocumento5 páginasREGULACIÓNCaesarAún no hay calificaciones
- Talle TranscripsionDocumento2 páginasTalle TranscripsionyuriAún no hay calificaciones
- Guía Molecular. 3er ParcialDocumento18 páginasGuía Molecular. 3er ParcialMichelle RodríguezAún no hay calificaciones
- Traduccion 2021Documento34 páginasTraduccion 2021Alexia Ramos CaetanoAún no hay calificaciones
- Transcripción y TraducciónDocumento12 páginasTranscripción y TraducciónJuana RamirezAún no hay calificaciones
- Parcial 4 de MicrobiologiaDocumento4 páginasParcial 4 de MicrobiologiaSantiago AgredaAún no hay calificaciones
- Rna InfografíaDocumento1 páginaRna InfografíaJennifer NaranjoAún no hay calificaciones
- TranscripcionDocumento54 páginasTranscripcionjosemaortizAún no hay calificaciones
- SEM - 7 SESION 25 y 26 - SINTESIS DE PROTEINAS TRANSAMINASASDocumento44 páginasSEM - 7 SESION 25 y 26 - SINTESIS DE PROTEINAS TRANSAMINASASJose Elver GalargaAún no hay calificaciones
- Cushicondor Alexis PAE8 AI 19 07 2022Documento3 páginasCushicondor Alexis PAE8 AI 19 07 2022Alexis CushicondorAún no hay calificaciones
- Química-Síntesis de Proteínas - 22-23Documento28 páginasQuímica-Síntesis de Proteínas - 22-23paula carmen alonso gonzalezAún no hay calificaciones
- Expresion Genica y Sintesis de ProteinasDocumento60 páginasExpresion Genica y Sintesis de ProteinasDiego Andres Riquelme VeraAún no hay calificaciones
- Monografia Biosintesis de ProteinasDocumento23 páginasMonografia Biosintesis de ProteinasSoberon BernabeAún no hay calificaciones
- Tema 29 TraducciónDocumento6 páginasTema 29 TraducciónHugo NavalónAún no hay calificaciones
- Actividades Transcr Traducc MutacionesDocumento2 páginasActividades Transcr Traducc MutacionesZoe V. SalernoAún no hay calificaciones
- Resumen de Destinación de ProteinasDocumento21 páginasResumen de Destinación de ProteinasMonserrat Cortez ArévaloAún no hay calificaciones
- Actividades Guía 3 Genética Molecular-ArnDocumento5 páginasActividades Guía 3 Genética Molecular-ArnOSCAR DAVID MARTINEZ SANABRIAAún no hay calificaciones
- P P TraduccionDocumento50 páginasP P TraduccionElsa MoranAún no hay calificaciones
- GTP T3.genética y Evolución 3 Parte Traducción 2016-17Documento60 páginasGTP T3.genética y Evolución 3 Parte Traducción 2016-17Fernanda SanchezAún no hay calificaciones
- Cap 8Documento52 páginasCap 8Estefi SolangeAún no hay calificaciones
- 4 Transcripcion y Regulación EucariotaDocumento32 páginas4 Transcripcion y Regulación EucariotaMarcela Ivette Fernández VelázquezAún no hay calificaciones
- Guia Sintesis de ProteinasDocumento7 páginasGuia Sintesis de Proteinaskaty alvarezAún no hay calificaciones
- Genetica I - Texto WordDocumento10 páginasGenetica I - Texto WordMichelle PalaciosAún no hay calificaciones
- Síntesis de ProteínasDocumento4 páginasSíntesis de ProteínasAriana VelardeAún no hay calificaciones
- Teorico 10 TraducciónDocumento11 páginasTeorico 10 TraducciónJulieta PedrouzoAún no hay calificaciones
- Síntesis de ProteínasDocumento4 páginasSíntesis de ProteínasRafaelDíazGuerreroAún no hay calificaciones
- GUIÓN DE LECTURA SOBRE LA SÍNTESIS DE PROTEÍNAS - SuperHiperMegaUltra ResumenDocumento13 páginasGUIÓN DE LECTURA SOBRE LA SÍNTESIS DE PROTEÍNAS - SuperHiperMegaUltra ResumenDaniel GuzmanAún no hay calificaciones
- Biol. MolecularDocumento75 páginasBiol. MolecularChristian AvendañoAún no hay calificaciones
- Ficha Informativa: Biosíntesis de Las ProteínasDocumento5 páginasFicha Informativa: Biosíntesis de Las ProteínasRuth ApazaAún no hay calificaciones
- Tema15 TranscripcionDocumento11 páginasTema15 TranscripcionIrene PG100% (1)
- Los Ribosomas y La Síntesis de ProteínasDocumento3 páginasLos Ribosomas y La Síntesis de ProteínasJuan Carlos Vázquez50% (4)
- 17 TraducciónDocumento84 páginas17 TraducciónJOSE ADRIAN ISAI GONZALEZ SANCHEZAún no hay calificaciones
- TRADUCCIÓN DEL mRNADocumento8 páginasTRADUCCIÓN DEL mRNAARUQUIPA AJLLAHUANCA GISELAAún no hay calificaciones
- Traduccion, Codigo Genetico BiologiaDocumento17 páginasTraduccion, Codigo Genetico BiologiaLaura SanchezAún no hay calificaciones
- Clase 9 Re Direccionamiento ProteicoDocumento16 páginasClase 9 Re Direccionamiento ProteicoAguss FernandezAún no hay calificaciones
- Clase 7 Metabolismo Del Grupo HemoDocumento17 páginasClase 7 Metabolismo Del Grupo HemoAguss FernandezAún no hay calificaciones
- Clase 6 Metabolismo de Ácidos NucleicosDocumento18 páginasClase 6 Metabolismo de Ácidos NucleicosAguss FernandezAún no hay calificaciones
- Clase 5 Metabolismo de AminoácidosDocumento21 páginasClase 5 Metabolismo de AminoácidosAguss FernandezAún no hay calificaciones
- Clase 4 Digestión ProteicaDocumento17 páginasClase 4 Digestión ProteicaAguss FernandezAún no hay calificaciones
- Clase 3 Estrés OxidativoDocumento17 páginasClase 3 Estrés OxidativoAguss FernandezAún no hay calificaciones
- Examen Pce Uned Biologia 2020Documento7 páginasExamen Pce Uned Biologia 2020Yamila YoAún no hay calificaciones
- Biologia - 3 - SolucionarioDocumento13 páginasBiologia - 3 - SolucionarioMiguel SalcedoAún no hay calificaciones
- Hipótesis Sobre El Origen de LaDocumento8 páginasHipótesis Sobre El Origen de Laysbecarmen100% (1)
- APUNTES BACHILLERATO BIOLOGIA ARN Mensaje Genetico PDFDocumento2 páginasAPUNTES BACHILLERATO BIOLOGIA ARN Mensaje Genetico PDFmerchinetsAún no hay calificaciones
- CRUCIBIOQDocumento2 páginasCRUCIBIOQCarolina LoaAún no hay calificaciones
- Realizar El Significado de 50 Palabras de La Rama de Ciencias NaturalesDocumento28 páginasRealizar El Significado de 50 Palabras de La Rama de Ciencias NaturalesJosue Villacreses VeraAún no hay calificaciones
- Glosario de GeneticaDocumento44 páginasGlosario de GeneticaSILVIA MOGROVEJOAún no hay calificaciones
- Ejercicios de Genetica Molecular PDFDocumento4 páginasEjercicios de Genetica Molecular PDFAle OrtegaAún no hay calificaciones
- Sesion 3 Sintesis de ProteinasDocumento41 páginasSesion 3 Sintesis de ProteinasAnonymous me3HTbZbh100% (2)
- RESUMEN GENETICA LiviaDocumento10 páginasRESUMEN GENETICA LiviaAivil MarquezAún no hay calificaciones
- NatDocumento9 páginasNatLitzy VerdugoAún no hay calificaciones
- 9 GeneticaMDocumento3 páginas9 GeneticaMmauricio fernandezAún no hay calificaciones
- Examen de BiologíaDocumento4 páginasExamen de BiologíaArancha Noemi MenchenAún no hay calificaciones
- TraducciónDocumento6 páginasTraducciónvaleriasantoyo289Aún no hay calificaciones
- Telegram - Biologia - Nicole Diaz - 5ADocumento2 páginasTelegram - Biologia - Nicole Diaz - 5ANicole DiazAún no hay calificaciones
- Transcripcion y TraduccionDocumento64 páginasTranscripcion y TraduccionFABIANA ANDREA DAZA ROCHAAún no hay calificaciones
- Mapa Mental CETEDocumento1 páginaMapa Mental CETEvelkis carreraAún no hay calificaciones
- Proceso de Replicación +transcripción+traducciónDocumento3 páginasProceso de Replicación +transcripción+traducciónjm rcAún no hay calificaciones
- Almaraz Ana Karen Act 1 Parcial 3Documento3 páginasAlmaraz Ana Karen Act 1 Parcial 3Ana Karen Almaraz RodríguezAún no hay calificaciones
- Ácidos NucleicosDocumento11 páginasÁcidos NucleicosJeisson Valencia PamplonaAún no hay calificaciones
- Sintesis de ProteinasDocumento24 páginasSintesis de ProteinasSofi CuevasAún no hay calificaciones
- Capitulo 47Documento32 páginasCapitulo 47apuntesmedicos2100% (1)
- Ut.5. Ácidos Nucleicos Y Enzimas AsociadasDocumento7 páginasUt.5. Ácidos Nucleicos Y Enzimas AsociadasTeresa AgrazAún no hay calificaciones
- Taller de BiologíaDocumento11 páginasTaller de BiologíaSara ValenciaAún no hay calificaciones
- Sintesis de ProteinasDocumento19 páginasSintesis de ProteinasSofia Lopez Aleman100% (1)
- Pronap 2019-1 CompletoDocumento140 páginasPronap 2019-1 Completoana86% (7)
- Caracteristicas Del ARNDocumento3 páginasCaracteristicas Del ARNdiegoAún no hay calificaciones
- Introduccion A La Genetica ClinicaDocumento28 páginasIntroduccion A La Genetica ClinicaGaby MoralesAún no hay calificaciones
- TALLER SINTESIS DE PROTEINAS ADocumento14 páginasTALLER SINTESIS DE PROTEINAS AKamilo Valera Moreno100% (1)
- TraducciónDocumento8 páginasTraducciónMaria FernandaAún no hay calificaciones