También podría gustarte
- Fisiología renal y metabolismo hidrosalino: Segunda ediciónDe EverandFisiología renal y metabolismo hidrosalino: Segunda ediciónAún no hay calificaciones
- Exposicion BioquimicaDocumento23 páginasExposicion Bioquimicamarco antonioAún no hay calificaciones
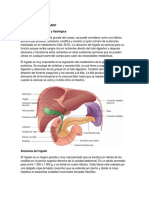
- PancreasDocumento10 páginasPancreasgiuliana fernandez campodonicoAún no hay calificaciones
- 2da charla cirugiaDocumento4 páginas2da charla cirugiaEstela QuirosAún no hay calificaciones
- Resumen ExposicionDocumento7 páginasResumen ExposicionStephanie RamirezAún no hay calificaciones
- Secreción PancreáticaDocumento7 páginasSecreción Pancreáticarociofigueroa.m130Aún no hay calificaciones
- Tema 4 GuíaDocumento5 páginasTema 4 GuíakeimaraularAún no hay calificaciones
- Digestion en Diferentes EspeciesDocumento11 páginasDigestion en Diferentes EspeciesHello CASHAún no hay calificaciones
- Guía metabolismo nitrógenoDocumento13 páginasGuía metabolismo nitrógenoMicaela del milagro DiazAún no hay calificaciones
- Fisiología Del PáncreasDocumento3 páginasFisiología Del PáncreasLucia TorresAún no hay calificaciones
- Resumen-Fisiología de La Secreción PancreáticaDocumento3 páginasResumen-Fisiología de La Secreción PancreáticaJose Alfredo HernandezAún no hay calificaciones
- Pregunta BioquimicaDocumento2 páginasPregunta BioquimicaJose SantanaAún no hay calificaciones
- Metabolismo de Los Cuerpo CetónicoDocumento7 páginasMetabolismo de Los Cuerpo CetónicoJordy Anthony Abanto RuizAún no hay calificaciones
- Información de Cuerpos CetónicosDocumento4 páginasInformación de Cuerpos CetónicosJUAN PABLO ESPINOSA ROSASAún no hay calificaciones
- BilisDocumento7 páginasBilisStalin QuijijeAún no hay calificaciones
- Jugos GastricosDocumento8 páginasJugos GastricosBryan Giovanny ArmentaAún no hay calificaciones
- SEMINARIO 6 GASTROINTESTINAL Sistema DigestivoDocumento9 páginasSEMINARIO 6 GASTROINTESTINAL Sistema DigestivoSamuel Millar FigueroaAún no hay calificaciones
- Digestivo 2 FisioDocumento6 páginasDigestivo 2 FisioAzazel RiseAún no hay calificaciones
- La digestión: proceso y regulaciónDocumento4 páginasLa digestión: proceso y regulaciónJose Luis Llamoza Romero50% (2)
- Secrecion GastricaDocumento6 páginasSecrecion Gastricaandres877Aún no hay calificaciones
- Funciones secretoras del tubo digestivoDocumento5 páginasFunciones secretoras del tubo digestivoKarla DuarteAún no hay calificaciones
- Transformacion de La Leche.Documento12 páginasTransformacion de La Leche.Katty CalderónAún no hay calificaciones
- Guia de Fisiologia de La Secrecion GastricaDocumento9 páginasGuia de Fisiologia de La Secrecion GastricaIsaAún no hay calificaciones
- Integración y Regulación Metabólica - Resumen Qca IIDocumento15 páginasIntegración y Regulación Metabólica - Resumen Qca IIVEROAún no hay calificaciones
- El Páncreas Fisiológicamente Esta Conformado Por Dos Tipos de Tejidos Que Determinan Su FunciónDocumento5 páginasEl Páncreas Fisiológicamente Esta Conformado Por Dos Tipos de Tejidos Que Determinan Su Funciónerika cabreraAún no hay calificaciones
- Biosíntesis Del ColesterolDocumento19 páginasBiosíntesis Del ColesterolFrank RodríguezAún no hay calificaciones
- Secreciones Del Páncreas ExocrinoDocumento6 páginasSecreciones Del Páncreas ExocrinoMila Apaza PilcoAún no hay calificaciones
- Cuestionario Digestivo 3Documento4 páginasCuestionario Digestivo 3KarlaAún no hay calificaciones
- Anatomía fisiología del páncreasDocumento4 páginasAnatomía fisiología del páncreasurpiAún no hay calificaciones
- Script FinalDocumento9 páginasScript FinalJouZyAún no hay calificaciones
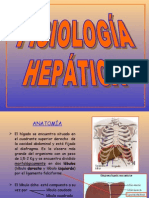
- Fisiologia HepaticaDocumento8 páginasFisiologia HepaticaYuli OrtizAún no hay calificaciones
- Metabolismo en el ayuno prolongado: procesos metabólicosDocumento9 páginasMetabolismo en el ayuno prolongado: procesos metabólicoslissette gabriela solis monarAún no hay calificaciones
- w4 Clase Bioca Cetogenesis Cetolisis Xenob 20052020Documento32 páginasw4 Clase Bioca Cetogenesis Cetolisis Xenob 20052020Darreem Steve Villarreal PutnamAún no hay calificaciones
- La gluconeogénesis, clave para la homeostasis glucémicaDocumento5 páginasLa gluconeogénesis, clave para la homeostasis glucémicaEl club de la LecturaAún no hay calificaciones
- Metabolismo de Las Proteínas Tarea BioquimicaDocumento47 páginasMetabolismo de Las Proteínas Tarea Bioquimicacybermar2005Aún no hay calificaciones
- Fisiologia HepáticaDocumento7 páginasFisiologia HepáticaKokabiel TamielAún no hay calificaciones
- Casos Clinicos y PreguntasDocumento4 páginasCasos Clinicos y PreguntasAngheles FrontanillaAún no hay calificaciones
- Secreción Gástrica y PancreáticaDocumento9 páginasSecreción Gástrica y PancreáticaBelen BerruttiAún no hay calificaciones
- CARBOHIDRATOS UNIFICADOoDocumento11 páginasCARBOHIDRATOS UNIFICADOoJesus Smaikel Teran MuenalaAún no hay calificaciones
- Tema 25 Clinicamedica ESTE SIDocumento4 páginasTema 25 Clinicamedica ESTE SIoasis fotocopiadosAún no hay calificaciones
- Anatomía y Funciones Principales Del HígadoDocumento4 páginasAnatomía y Funciones Principales Del Hígadonathaliecastillo758Aún no hay calificaciones
- Fisiologia HepaticaDocumento21 páginasFisiologia HepaticaLalo Valdivia100% (1)
- FORMACIÓN Y SECRECIÓN DE LA BILISDocumento5 páginasFORMACIÓN Y SECRECIÓN DE LA BILISAlejandroAún no hay calificaciones
- Via de La GluconeogenesisDocumento38 páginasVia de La Gluconeogenesisfmh200967% (6)
- Segundo Caso Clinico-Histologia - Aparato DigestivoDocumento5 páginasSegundo Caso Clinico-Histologia - Aparato DigestivoRod CuevaAún no hay calificaciones
- Efectos metabólicos insulina glucagón tejidosDocumento13 páginasEfectos metabólicos insulina glucagón tejidoskaty gradosAún no hay calificaciones
- Colecistoquinina y SecretinaDocumento2 páginasColecistoquinina y SecretinaGeranni RondonAún no hay calificaciones
- Lipoproteínas y metabolismo de lípidosDocumento5 páginasLipoproteínas y metabolismo de lípidosDeisy LópezAún no hay calificaciones
- Hormonas Digestivas-Sandra Delcarpio AzañeroDocumento15 páginasHormonas Digestivas-Sandra Delcarpio AzañeroSandra Adela Del Carpio AzañeroAún no hay calificaciones
- Hormonas gastrointestinales y secreción gástricaDocumento23 páginasHormonas gastrointestinales y secreción gástricapaulaAún no hay calificaciones
- Pepsina y PepsinogenoDocumento5 páginasPepsina y PepsinogenoAlvaro Aquije100% (1)
- Apuntes Insulina-Glucosa Dra v. MujicaDocumento15 páginasApuntes Insulina-Glucosa Dra v. MujicaNicol Ibarra ArellanoAún no hay calificaciones
- Cetogénesis: producción y regulación de cuerpos cetónicosDocumento5 páginasCetogénesis: producción y regulación de cuerpos cetónicosvianyse100% (1)
- S10 - Proteínas IDocumento11 páginasS10 - Proteínas IFabiola Chuquillanqui veliAún no hay calificaciones
- Digestivo Colorado II PREGUNTASDocumento9 páginasDigestivo Colorado II PREGUNTASDANIEL ALEJANDROAún no hay calificaciones
- Archivo Exposicion Secrecion PancreaticaDocumento14 páginasArchivo Exposicion Secrecion PancreaticaVanessa De AguasAún no hay calificaciones
- Bioquimica de Aparato DigestivoDocumento8 páginasBioquimica de Aparato DigestivoDesiret Sanchez50% (2)
- Resumen UP 1Documento37 páginasResumen UP 1danilonaimespindolaAún no hay calificaciones
- Metabolismos Del CerebroDocumento5 páginasMetabolismos Del CerebroLilibeth RangelAún no hay calificaciones
- Stryer BIBLIOGRAFIA CLASE 26Documento7 páginasStryer BIBLIOGRAFIA CLASE 26Andrea MongeAún no hay calificaciones
- Herrera BIBLIOGRAFIA CLASE 26Documento9 páginasHerrera BIBLIOGRAFIA CLASE 26Andrea MongeAún no hay calificaciones
- Marks BIBLIOGRAFIA CLASE 23Documento7 páginasMarks BIBLIOGRAFIA CLASE 23Andrea MongeAún no hay calificaciones
- Harper BIBLIOGRAFIA CLASE 23Documento3 páginasHarper BIBLIOGRAFIA CLASE 23Andrea MongeAún no hay calificaciones
- Harper BIBLIOGRAFIA CLASE 24Documento2 páginasHarper BIBLIOGRAFIA CLASE 24Andrea MongeAún no hay calificaciones
- LBaynes BIBLIOGRAFIA CLASE 24Documento5 páginasLBaynes BIBLIOGRAFIA CLASE 24Andrea MongeAún no hay calificaciones
- Marks BIBLIOGRAFIA CLASE 24Documento8 páginasMarks BIBLIOGRAFIA CLASE 24Andrea MongeAún no hay calificaciones
- Laboratorio N°4 - Citocromo OxidasaDocumento5 páginasLaboratorio N°4 - Citocromo OxidasaAndrea MongeAún no hay calificaciones
- Laboratorio 4 MicroDocumento32 páginasLaboratorio 4 MicroAndrea MongeAún no hay calificaciones
- Laboratorio 5 BiocaDocumento6 páginasLaboratorio 5 BiocaAndrea MongeAún no hay calificaciones
- Laboratorio 5 MacroDocumento32 páginasLaboratorio 5 MacroAndrea MongeAún no hay calificaciones
- Cal HidratadaDocumento2 páginasCal HidratadaFerney Mauricio Calderón RincónAún no hay calificaciones
- Seminario Paleo Mafil - PPT Vergara-Contreras PDFDocumento53 páginasSeminario Paleo Mafil - PPT Vergara-Contreras PDFSan PedroAún no hay calificaciones
- El Ñanduti o Encaje Del ParaguayDocumento6 páginasEl Ñanduti o Encaje Del ParaguayklaudiaAún no hay calificaciones
- Circular 001 - 2005Documento9 páginasCircular 001 - 2005Angel Rubén Ruiz LópezAún no hay calificaciones
- Caso Demanda de AlimentosDocumento2 páginasCaso Demanda de AlimentosActivoz promotora inmobiliariaAún no hay calificaciones
- Fundamentos EconómicosDocumento18 páginasFundamentos EconómicosNhora GelvezAún no hay calificaciones
- Aracne y La Diosa AteneaDocumento1 páginaAracne y La Diosa AteneaIara RossoAún no hay calificaciones
- Globalizado 1er BimestreDocumento3 páginasGlobalizado 1er BimestreJimmy Manrique EnriquezAún no hay calificaciones
- Formulario de Examen Xunta de Galicia 2019Documento20 páginasFormulario de Examen Xunta de Galicia 2019alumno120Aún no hay calificaciones
- Acta DistribuciondividendoDocumento5 páginasActa DistribuciondividendoClau Pa100% (1)
- Max Weber: El origen del espíritu capitalistaDocumento56 páginasMax Weber: El origen del espíritu capitalistaAna BonlluvanniAún no hay calificaciones
- 2 Origen de Los OceanosDocumento15 páginas2 Origen de Los Oceanosemerson leyva leónAún no hay calificaciones
- Metodos Alternativos Solución ConflictosDocumento6 páginasMetodos Alternativos Solución Conflictostomacho21Aún no hay calificaciones
- Infertilidad PDFDocumento2 páginasInfertilidad PDFG Jacqueline DíazAún no hay calificaciones
- Asesoria de Impacto HogaramaDocumento7 páginasAsesoria de Impacto HogaramaJose GuzmanAún no hay calificaciones
- Guía completa para dominar Excel: Formato, cálculos, gráficos y másDocumento3 páginasGuía completa para dominar Excel: Formato, cálculos, gráficos y másJhonny RodasAún no hay calificaciones
- Proyecto de Aprendizaje P.A.Documento47 páginasProyecto de Aprendizaje P.A.mnieto2009100% (12)
- Unxx Toxicocinetica y Toxicodinamia Del Monoxido de CarbonoDocumento13 páginasUnxx Toxicocinetica y Toxicodinamia Del Monoxido de CarbonoSantiago leon de pradaAún no hay calificaciones
- GM apuesta por el Chevy VoltDocumento3 páginasGM apuesta por el Chevy VoltJosé Miguel Gonzalez PihuaveAún no hay calificaciones
- KING Versión de Montaje 1Documento36 páginasKING Versión de Montaje 1Daniel de La OAún no hay calificaciones
- Fisica Problemas Resueltos Part 1Documento3 páginasFisica Problemas Resueltos Part 1James ArredondoAún no hay calificaciones
- Prueba General para A.A y Proteinas-with-cover-page-V2Documento11 páginasPrueba General para A.A y Proteinas-with-cover-page-V2Nelson CruzAún no hay calificaciones
- Informe psicológico niño 8 añosDocumento2 páginasInforme psicológico niño 8 añossol arcosAún no hay calificaciones
- La MetamorfosisDocumento44 páginasLa MetamorfosisKarol Yanahi Garnica BizarroAún no hay calificaciones
- Modelo Sistemico FinalDocumento16 páginasModelo Sistemico FinalJulia Godínez100% (1)
- 1504 Historia de Mexico 2Documento20 páginas1504 Historia de Mexico 2Maria Reynolds-HamiltonAún no hay calificaciones
- Arquitectura Islamica Ventura Santos Alex GabrielDocumento11 páginasArquitectura Islamica Ventura Santos Alex Gabrielalex venturaAún no hay calificaciones
- Abrir MENU - BARRAHERMOSA - JUN2023Documento6 páginasAbrir MENU - BARRAHERMOSA - JUN2023Santiago Casanova RejonAún no hay calificaciones
- Definicion y Alacance de Los Deberes, Derechos y Garantia de Los VenezolanosDocumento11 páginasDefinicion y Alacance de Los Deberes, Derechos y Garantia de Los VenezolanosRhenan Bolivar0% (1)
- Especialidad de Nudos y AmarrasDocumento18 páginasEspecialidad de Nudos y AmarrasJhandira MamajaiAún no hay calificaciones