También podría gustarte
- Funciones y anatomía del cerebeloDocumento10 páginasFunciones y anatomía del cerebeloAnthony RíosAún no hay calificaciones
- Funciones Cerebelosas.Documento12 páginasFunciones Cerebelosas.MariGuedezGuAún no hay calificaciones
- CerebeloDocumento21 páginasCerebelowilkermanAún no hay calificaciones
- CEREBELODocumento9 páginasCEREBELOsuquillopAún no hay calificaciones
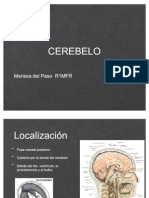
- CEREBELODocumento52 páginasCEREBELOMarissa del Paso100% (1)
- Funciones Del CerebeloDocumento5 páginasFunciones Del CerebeloNIÑAS MEDICINA100% (3)
- Morfofisiologia Humana 2 Orientadora 6 ValeriaDocumento3 páginasMorfofisiologia Humana 2 Orientadora 6 ValeriaValeria Reyes100% (2)
- APUNTE Clase 10Documento12 páginasAPUNTE Clase 10Felipe GuzmanAún no hay calificaciones
- Ce RebeloDocumento10 páginasCe RebeloÁngeles Inostroza SilvaAún no hay calificaciones
- Motor Cerebelo.Documento16 páginasMotor Cerebelo.Marcelita VelardeAún no hay calificaciones
- Resumen Capitulo 3 Fisologia Del Control MotorDocumento8 páginasResumen Capitulo 3 Fisologia Del Control MotorCatalina Mahecha CubillosAún no hay calificaciones
- 8 - MOTOR II - 3.3 - CerebeloDocumento8 páginas8 - MOTOR II - 3.3 - CerebeloFacundo RodriguezAún no hay calificaciones
- Presentacion CerebeloDocumento31 páginasPresentacion CerebeloLuisa BucioAún no hay calificaciones
- #4 CerebeloDocumento8 páginas#4 CerebeloMonserrat Olivo TalaveraAún no hay calificaciones
- CEREBELODocumento7 páginasCEREBELOSebastián GaiborAún no hay calificaciones
- Funcion Del CerebeloDocumento2 páginasFuncion Del CerebeloCristobal CantilloAún no hay calificaciones
- Circuitos CerebelososDocumento7 páginasCircuitos CerebelososVane BustamanteAún no hay calificaciones
- CEREBELODocumento11 páginasCEREBELOBritez Paredes Alejandra SilvanaAún no hay calificaciones
- Neur - Vestíbulo CerebeloDocumento3 páginasNeur - Vestíbulo CerebeloKen MoyaAún no hay calificaciones
- Morfología y funciones del cerebelo y tronco encefálicoDocumento19 páginasMorfología y funciones del cerebelo y tronco encefálicoJE King100% (1)
- Sistema Motor Somatico (Reydel Gonzalez)Documento6 páginasSistema Motor Somatico (Reydel Gonzalez)Reydel Gonzalez RodriguezAún no hay calificaciones
- Cerebelo y Ganglios Basales PDFDocumento14 páginasCerebelo y Ganglios Basales PDFCandela MercadoAún no hay calificaciones
- Contribuciones Del Cerebelo y Los Ganglios Basales Al Control Motor GlobalDocumento5 páginasContribuciones Del Cerebelo y Los Ganglios Basales Al Control Motor Globaleladio recalde100% (2)
- ELCEREBELODocumento5 páginasELCEREBELOIgnacio Andres Verdugo MontecinosAún no hay calificaciones
- Sindrome CerebelosoDocumento13 páginasSindrome CerebelosoorianamarinaAún no hay calificaciones
- El Cerebro y El CerebeloDocumento8 páginasEl Cerebro y El CerebeloWilmer Alberto Gonzalez BlancoAún no hay calificaciones
- Modulación Del Movimiento Por El CerebeloDocumento23 páginasModulación Del Movimiento Por El Cerebeloclarisa.rivera.2000Aún no hay calificaciones
- Tema 3. Psicobio II.Documento4 páginasTema 3. Psicobio II.irechmtnzAún no hay calificaciones
- Cerebelo: Estructura, Funciones y AlteracionesDocumento43 páginasCerebelo: Estructura, Funciones y AlteracionesDaniela Garrido Tapia100% (1)
- Cerebelo AtaxiaDocumento45 páginasCerebelo AtaxianekotefiAún no hay calificaciones
- CEREBELO - ResumenDocumento10 páginasCEREBELO - ResumensalvayopsAún no hay calificaciones
- Fibras Eferentes CerebelosasDocumento10 páginasFibras Eferentes CerebelosasCdcgs PWinAún no hay calificaciones
- Semana N°07Documento69 páginasSemana N°07DJ EDWAún no hay calificaciones
- Control de Funciones Motoras Por La Corteza y El Tronco Del EncéfaloDocumento73 páginasControl de Funciones Motoras Por La Corteza y El Tronco Del EncéfaloPamela CabrejaAún no hay calificaciones
- Taller Sistema VestibularDocumento5 páginasTaller Sistema VestibularJEISSON ARLEY HERNANDEZ CASTILLO100% (1)
- Clase 4 Cerebelo y GangliosDocumento67 páginasClase 4 Cerebelo y GangliosAlejandro AlejandroAún no hay calificaciones
- Tarea de La Semana 5. Cerebelo MFHIIDocumento4 páginasTarea de La Semana 5. Cerebelo MFHIIEuquely LesmoAún no hay calificaciones
- Bynf Vi-MovimientoDocumento23 páginasBynf Vi-MovimientoPaula PajónAún no hay calificaciones
- El CerebeloDocumento7 páginasEl CerebeloArleen SalinasAún no hay calificaciones
- Resumen Clase de CerebeloDocumento15 páginasResumen Clase de CerebeloSamyack Apcha100% (1)
- Fisio Sistemas Tema 3Documento4 páginasFisio Sistemas Tema 3GARY CARHUAMACAAún no hay calificaciones
- Vesiculas Cerebrales SecundariasDocumento8 páginasVesiculas Cerebrales SecundariasMaria Virginia Sosa RodriguezAún no hay calificaciones
- Cerebelo 2Documento2 páginasCerebelo 2claravega120618Aún no hay calificaciones
- Función motora corteza y tronco encéfaloDocumento7 páginasFunción motora corteza y tronco encéfaloMirian GutierrezAún no hay calificaciones
- CEREBELODocumento38 páginasCEREBELOYam AriasAún no hay calificaciones
- 1 CerebeloDocumento25 páginas1 CerebeloINES VASQUEZAún no hay calificaciones
- Capítulo 56 - Cerebelo y Ganglios BasalesDocumento8 páginasCapítulo 56 - Cerebelo y Ganglios BasalesJesica AndanAún no hay calificaciones
- Sindrome CerebelosoDocumento59 páginasSindrome Cerebelosohemer666100% (14)
- Cap 57, Cerebelo y Ganglios Basales Función MotoraDocumento19 páginasCap 57, Cerebelo y Ganglios Basales Función Motoraedin100% (1)
- CEREBELODocumento15 páginasCEREBELOMaia CucchianiAún no hay calificaciones
- Vías somatomotoras: control del movimientoDocumento27 páginasVías somatomotoras: control del movimientoMartha HerreraAún no hay calificaciones
- El Cerebelo y Sus ConexionesDocumento4 páginasEl Cerebelo y Sus ConexionesBarbara InsulzaAún no hay calificaciones
- Informe FisiologiaDocumento7 páginasInforme FisiologiaDaniela RodriguesAún no hay calificaciones
- Sistema visual y organización jerárquica del sistema motorDocumento38 páginasSistema visual y organización jerárquica del sistema motorFelipe Barrios BravoAún no hay calificaciones
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- Anatomia y Fisiología del Sistema Nervioso IIDe EverandAnatomia y Fisiología del Sistema Nervioso IICalificación: 4 de 5 estrellas4/5 (28)
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2De EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Aún no hay calificaciones
- El cannabis en patologías del sistema nervioso centralDe EverandEl cannabis en patologías del sistema nervioso centralAún no hay calificaciones
- Dolor anterior de rodilla: Síndrome de dolor femoropatelarDocumento12 páginasDolor anterior de rodilla: Síndrome de dolor femoropatelarDiegoAún no hay calificaciones
- Test Milani Comparetti 445412 Downloable 2401484Documento2 páginasTest Milani Comparetti 445412 Downloable 2401484DiegoAún no hay calificaciones
- Hombro: Hombro DolorosoDocumento13 páginasHombro: Hombro DolorosoDiegoAún no hay calificaciones
- IRRADIACIONDocumento7 páginasIRRADIACIONDiegoAún no hay calificaciones
- Sindrome Del Canal de GuyonDocumento1 páginaSindrome Del Canal de GuyonDiegoAún no hay calificaciones
- Falsos Amigos - HeterosemânticasDocumento1 páginaFalsos Amigos - HeterosemânticasDiegoAún no hay calificaciones
- Induccion SucesivaDocumento3 páginasInduccion SucesivaDiegoAún no hay calificaciones
- Dolor en CaderaDocumento10 páginasDolor en CaderaDiegoAún no hay calificaciones
- Parkinson y Discinesias 2012Documento15 páginasParkinson y Discinesias 2012DiegoAún no hay calificaciones
- Laser terapia baja potenciaDocumento2 páginasLaser terapia baja potenciaDiegoAún no hay calificaciones
- Vancouver Citar y ReferenciarDocumento20 páginasVancouver Citar y ReferenciarDiegoAún no hay calificaciones
- Scribd ATAXIADocumento2 páginasScribd ATAXIADiegoAún no hay calificaciones
- COXINHADocumento9 páginasCOXINHADiegoAún no hay calificaciones
- Ajustes Posturales AnticipatoriosDocumento2 páginasAjustes Posturales AnticipatoriosDiegoAún no hay calificaciones
- Metodología de La Investigación en Ciencias Médicas y SaludDocumento449 páginasMetodología de La Investigación en Ciencias Médicas y SaludLuisHerreraLópez100% (3)
- CerebellumDocumento8 páginasCerebellumDiegoAún no hay calificaciones
- RODILLADocumento11 páginasRODILLADiegoAún no hay calificaciones
- Scribd ENFERMEDAD DE PARKINSONDocumento2 páginasScribd ENFERMEDAD DE PARKINSONDiegoAún no hay calificaciones
- Scribd HemiplejiaDocumento2 páginasScribd HemiplejiaDiegoAún no hay calificaciones
- Corrientes Electricas Según Su FrecuenciaDocumento7 páginasCorrientes Electricas Según Su FrecuenciaDiegoAún no hay calificaciones
- Clinica Topografica de La HemiplejiaDocumento2 páginasClinica Topografica de La HemiplejiaDiegoAún no hay calificaciones
- C02 AnatomiaHumana 2aedDocumento15 páginasC02 AnatomiaHumana 2aedDiegoAún no hay calificaciones
- Scribd Lesion MedularDocumento1 páginaScribd Lesion MedularDiegoAún no hay calificaciones
- Agenda 2030 Los Objetivos de Desarrollo Sostenible y El Envejecimiento GlobalDocumento16 páginasAgenda 2030 Los Objetivos de Desarrollo Sostenible y El Envejecimiento Globalsandra estepanAún no hay calificaciones
- Parkinson y Discinesias 2012Documento15 páginasParkinson y Discinesias 2012DiegoAún no hay calificaciones
- Indicaciones: Comparación Pre y Post Tratamiento deDocumento2 páginasIndicaciones: Comparación Pre y Post Tratamiento deDiegoAún no hay calificaciones
- Tratamiento Post QuirúrgicoDocumento2 páginasTratamiento Post QuirúrgicoDiegoAún no hay calificaciones
- Facilitación en Pelota para NiñosDocumento4 páginasFacilitación en Pelota para NiñosDiego100% (1)
- Parkinson y Discinesias 2012Documento15 páginasParkinson y Discinesias 2012DiegoAún no hay calificaciones
- C Lculo Diferencial IF FIA UAQ 2021 II ADocumento355 páginasC Lculo Diferencial IF FIA UAQ 2021 II AIGNACIO ISMAEL FLORES LANDAVERDEAún no hay calificaciones
- 1786 PDFDocumento464 páginas1786 PDFWilmer Galvis100% (1)
- Espectroscopia AtomicaDocumento6 páginasEspectroscopia AtomicaValentina Franco MartinezAún no hay calificaciones
- 2 Area Central-Area Monumental-Centro HistoricoDocumento14 páginas2 Area Central-Area Monumental-Centro HistoricoLourdes BordaAún no hay calificaciones
- Pentacloro saume fungicida de semillaDocumento3 páginasPentacloro saume fungicida de semillaChristian A. Navarro BazalarAún no hay calificaciones
- El Viaje de La CometaDocumento2 páginasEl Viaje de La CometaMOnse ToapantaAún no hay calificaciones
- Marjorie Ramos Semana5Documento5 páginasMarjorie Ramos Semana5Marjorie RamosAún no hay calificaciones
- Informes Caminadores 2015Documento9 páginasInformes Caminadores 2015Johanna SáenzAún no hay calificaciones
- Pruebas Saber de Undc3a9cimo Series Sucesiones y Lc3admites1Documento2 páginasPruebas Saber de Undc3a9cimo Series Sucesiones y Lc3admites1Flor Yadira Moreno GarzonAún no hay calificaciones
- Qué Aire NecesitoDocumento2 páginasQué Aire NecesitoMalka Sarmiento100% (1)
- Presentacion Esquematica de La Cadena de AbastecimientoDocumento29 páginasPresentacion Esquematica de La Cadena de AbastecimientoDeivid Leonardo Gallo PascagazaAún no hay calificaciones
- Ensayo Argumentativo N°1 FinalisimoDocumento4 páginasEnsayo Argumentativo N°1 FinalisimoKelyZulemaTacoraDuranAún no hay calificaciones
- Plan Educativo Aprendamos Juntos en Casa - Compressed PDFDocumento96 páginasPlan Educativo Aprendamos Juntos en Casa - Compressed PDFEdison Cordova88% (8)
- Adquisición yeso estadio UrubambaDocumento4 páginasAdquisición yeso estadio UrubambaSharon LeivaAún no hay calificaciones
- Practica 1 BiocaDocumento7 páginasPractica 1 BiocaVAZQUEZ GOMEZ DEL VILLAR MICHELLEAún no hay calificaciones
- Copia de Analisis Del Impacto Del NegocioDocumento1 páginaCopia de Analisis Del Impacto Del NegocioElver ToboAún no hay calificaciones
- Análisis de puestos, descripción y diseñoDocumento5 páginasAnálisis de puestos, descripción y diseñoMiguel AngelAún no hay calificaciones
- El Discurso de Graduación.Documento6 páginasEl Discurso de Graduación.Milagro Gracia0% (1)
- 124 - Guia Emprendimiento 11 2 Periodo 2020Documento3 páginas124 - Guia Emprendimiento 11 2 Periodo 2020loren niñoAún no hay calificaciones
- Planificacion 5to. Perito Admon Conta FinancieraDocumento5 páginasPlanificacion 5to. Perito Admon Conta FinancieraLuis RodriguezAún no hay calificaciones
- RESUMEN (1) (1) PopperDocumento2 páginasRESUMEN (1) (1) Popperapi-375628582% (33)
- Fundamentos de Inyeccion de PlasticosDocumento17 páginasFundamentos de Inyeccion de PlasticosJuan Ross67% (3)
- 6to de Primaria Corazon (Cartilla)Documento16 páginas6to de Primaria Corazon (Cartilla)doris morandoAún no hay calificaciones
- PENSAMIENTO Económico InicioDocumento13 páginasPENSAMIENTO Económico InicioUriel Yue ValdiviaAún no hay calificaciones
- Manual de Administracion de ContratosDocumento41 páginasManual de Administracion de Contratoscarmen espinozaAún no hay calificaciones
- Actividad Entregable 2 FisicaDocumento6 páginasActividad Entregable 2 FisicaEL BRANDON100% (1)
- Analisis Foda: Fortalezas AmenazasDocumento2 páginasAnalisis Foda: Fortalezas AmenazasJacquelin Andrea Corado BautistaAún no hay calificaciones
- Cuaderno de Campo-Gestalt y ConstelacionesDocumento56 páginasCuaderno de Campo-Gestalt y ConstelacionesveronicacerAún no hay calificaciones
- Datos Del Estudiante: Facultad de Ciencias EmpresarialesDocumento32 páginasDatos Del Estudiante: Facultad de Ciencias Empresarialesmario paradaAún no hay calificaciones