También podría gustarte
- OMS - Manual de Bioseguridad en El LaboratorioDocumento223 páginasOMS - Manual de Bioseguridad en El LaboratorioCarlos de Paz90% (30)
- CASOS A RESOLVER MalariaDocumento4 páginasCASOS A RESOLVER MalariaFERNANDO GOMEZ ORDOÑEZAún no hay calificaciones
- Estructura y funciones de las inmunoglobulinasDocumento11 páginasEstructura y funciones de las inmunoglobulinasGloria Mardeni100% (1)
- Línea Del Tiempo GenéticaDocumento3 páginasLínea Del Tiempo GenéticaLeslie LouAún no hay calificaciones
- CAPÍTULO 11 - Desarrollo de La PlacentaDocumento11 páginasCAPÍTULO 11 - Desarrollo de La PlacentaAnahi Ortiz100% (3)
- Curva de disociación de la hemoglobina: factores que desplazan la curva y su importancia fisiológicaDocumento36 páginasCurva de disociación de la hemoglobina: factores que desplazan la curva y su importancia fisiológicaLuis Medina100% (1)
- Ensayo CarbohidratosDocumento9 páginasEnsayo CarbohidratosPabloLopez0% (1)
- A Enzimas en Los Alimentos 14mexDocumento14 páginasA Enzimas en Los Alimentos 14mexRALPH STEIN RIVERA BOTONARESAún no hay calificaciones
- Estructura y Función de La HemoglobinaDocumento15 páginasEstructura y Función de La HemoglobinaIsabella CatañoAún no hay calificaciones
- Trabajo HemoglobinaDocumento3 páginasTrabajo HemoglobinaAlejandroToqueAún no hay calificaciones
- Expo - Taller 9 MOBILUMCUSDocumento12 páginasExpo - Taller 9 MOBILUMCUSIsabel CRAún no hay calificaciones
- Metabolismo Del HierroDocumento3 páginasMetabolismo Del HierroDeyvis Escalante100% (1)
- Clase de SarconidosDocumento14 páginasClase de SarconidosTania PardoAún no hay calificaciones
- Creatinina en Suero Metodo ColorimetricoDocumento2 páginasCreatinina en Suero Metodo ColorimetricoElenaL100% (10)
- Hist. Parasitologia Siglo XXDocumento3 páginasHist. Parasitologia Siglo XXDianelis DuarteAún no hay calificaciones
- Disoluciones AmortiguadorasDocumento31 páginasDisoluciones Amortiguadoraspaola sofia cordova huamani0% (1)
- Antropologia Bio Estadistica Y Demografia Medica: Universidad de Aquino Bolivia UdabolDocumento119 páginasAntropologia Bio Estadistica Y Demografia Medica: Universidad de Aquino Bolivia UdabolMarcy de los ÁngelesAún no hay calificaciones
- Pelvis EspinozaDocumento131 páginasPelvis EspinozaRenzo RiveraAún no hay calificaciones
- Metabolismo de Quilomicrones y VLDLDocumento3 páginasMetabolismo de Quilomicrones y VLDLCRISTINA DIAZ LOPEZAún no hay calificaciones
- Capitulo 14 GuytonDocumento29 páginasCapitulo 14 GuytonMaridiose Ruiz Barrera100% (2)
- COMPILADO CHOICE FISIOLOGÍA UA2 R1 (Parte2)Documento16 páginasCOMPILADO CHOICE FISIOLOGÍA UA2 R1 (Parte2)Deyanira AdaAún no hay calificaciones
- Mol, Osmolaridad, Molalildad y MolaridadDocumento5 páginasMol, Osmolaridad, Molalildad y MolaridadSamsaraCT PokémonGo100% (1)
- Técnica de Fructosamina en SueroDocumento2 páginasTécnica de Fructosamina en SueroQUIMICO CLINICO WILLIANS SANCHEZ100% (1)
- Niridazol, Niclosamida y LevamisolDocumento6 páginasNiridazol, Niclosamida y LevamisolCarlos EspinozaAún no hay calificaciones
- Drenaje VenosoDocumento20 páginasDrenaje VenosoAracellyHerreraAún no hay calificaciones
- Investigación métodos 40cDocumento807 páginasInvestigación métodos 40cCesar ChoqueAún no hay calificaciones
- Parasito Mansonelosis InformeDocumento22 páginasParasito Mansonelosis InformeLaura Aguilar PerezAún no hay calificaciones
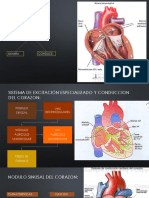
- Excitacion Ritmica Del CorazonDocumento9 páginasExcitacion Ritmica Del CorazonJose Velasquez TeranAún no hay calificaciones
- Vasoconstricción pulmonar hipoxiaDocumento19 páginasVasoconstricción pulmonar hipoxiaCapige CapiAún no hay calificaciones
- Senescencia Lesiones de AlmacenamientoDocumento6 páginasSenescencia Lesiones de Almacenamientocatalina AlcarazAún no hay calificaciones
- Sindrome de Lesh NyhanDocumento10 páginasSindrome de Lesh NyhanBirzabit Dominguez LopezAún no hay calificaciones
- Exitacion Ritmica Del CorazonDocumento26 páginasExitacion Ritmica Del CorazonKristell Pérez100% (2)
- Práctica N°7: Separación de Proteínas en La Sangre PDFDocumento8 páginasPráctica N°7: Separación de Proteínas en La Sangre PDFSarahAún no hay calificaciones
- Fenomeno de ArthusDocumento3 páginasFenomeno de ArthusRuben100% (1)
- HEMOGLOBINADocumento18 páginasHEMOGLOBINAkatlin gonzalezAún no hay calificaciones
- Marco de Inmuno3Documento3 páginasMarco de Inmuno3Ani NicolasAún no hay calificaciones
- Vena Toracica Interna o Vena Mamaria InternaDocumento3 páginasVena Toracica Interna o Vena Mamaria InternaAngelica Paredes CubaAún no hay calificaciones
- GLUCÓGENOSIS TIPO IIIDocumento10 páginasGLUCÓGENOSIS TIPO IIIEmmanuel PaniaguaAún no hay calificaciones
- Coccidioidomicosis: hongo dimórfico agente causalDocumento5 páginasCoccidioidomicosis: hongo dimórfico agente causaljuanAún no hay calificaciones
- Anguiano Gonzalez Mariana FACTOR DEPRESOR DEL MIOCARDIODocumento17 páginasAnguiano Gonzalez Mariana FACTOR DEPRESOR DEL MIOCARDIOMariana Anguiano GonzalezAún no hay calificaciones
- Kato MiuraDocumento17 páginasKato MiuraCeci Mendez0% (1)
- Metabolismo Del Agua y ElectrolitosDocumento15 páginasMetabolismo Del Agua y ElectrolitosVanessa Tapia QuirozAún no hay calificaciones
- Guia Fisiologia IDocumento151 páginasGuia Fisiologia Ijosmiguel_bv100% (1)
- Gonadotropina Coriónica HumanaDocumento6 páginasGonadotropina Coriónica HumanariacidmiAún no hay calificaciones
- Atlas de ParasitologiaDocumento118 páginasAtlas de ParasitologiaKaren Alavez0% (1)
- ERITROCITOSDocumento41 páginasERITROCITOSAylén AguirreAún no hay calificaciones
- Regulación osmolaridad orinaDocumento12 páginasRegulación osmolaridad orinaZilene Córdova EstradaAún no hay calificaciones
- Practica 8Documento6 páginasPractica 8Ysmael Barrientos100% (1)
- Amortiguadores de PH FlujogramaDocumento5 páginasAmortiguadores de PH FlujogramaEvelyn RodriguezAún no hay calificaciones
- Medicina RomanaDocumento26 páginasMedicina RomanaItala RoncoAún no hay calificaciones
- Preguntas Resueltas Del Libro Thompson y Thompson de GenéticaDocumento5 páginasPreguntas Resueltas Del Libro Thompson y Thompson de GenéticaSihomara Alejandra A CruzAún no hay calificaciones
- Histologia Sistema CirculatorioDocumento11 páginasHistologia Sistema Circulatoriokelvin Tapulllima PashanasiAún no hay calificaciones
- Proteínas: Composición y estructuraDocumento36 páginasProteínas: Composición y estructurajuan sebastian parra boadaAún no hay calificaciones
- Caso Clínico Semana 12.2020.2Documento33 páginasCaso Clínico Semana 12.2020.2LuisAún no hay calificaciones
- Quimica SanguineaDocumento15 páginasQuimica SanguineaGustavo100% (1)
- Bacterias 1Documento51 páginasBacterias 1Manuel MuñozAún no hay calificaciones
- Fisiología del sistema endocrinoDocumento16 páginasFisiología del sistema endocrinoSHEILA CESPEDESAún no hay calificaciones
- Triángulo de Calot: disección de elementos biliares y vascularesDocumento6 páginasTriángulo de Calot: disección de elementos biliares y vascularesDaisuke0% (1)
- Circulacion UteroplacentariaDocumento4 páginasCirculacion UteroplacentariaAle CachoAún no hay calificaciones
- HemoglobinaDocumento6 páginasHemoglobinamairaAún no hay calificaciones
- HEMOGLOBINADocumento57 páginasHEMOGLOBINArebeca moreliAún no hay calificaciones
- Unidad 4 - ProteinaDocumento16 páginasUnidad 4 - ProteinaprimeliaAún no hay calificaciones
- 01 Motilidadjorge 141121102219 Conversion Gate02 PDFDocumento66 páginas01 Motilidadjorge 141121102219 Conversion Gate02 PDFprimeliaAún no hay calificaciones
- SALMONELLADocumento16 páginasSALMONELLAprimeliaAún no hay calificaciones
- Unidad 25 - Balance Hidromineral Equilibrio Ácido-BaseDocumento19 páginasUnidad 25 - Balance Hidromineral Equilibrio Ácido-BaseprimeliaAún no hay calificaciones
- Unidad 3-Acidos NucleicosDocumento9 páginasUnidad 3-Acidos NucleicosprimeliaAún no hay calificaciones
- Taxonomia BasicaDocumento8 páginasTaxonomia BasicaAnonymous WpLhklaAún no hay calificaciones
- Agar SsDocumento2 páginasAgar SsAnonymous 1d7b0eUhyAún no hay calificaciones
- Bacterias Gastrointestinales PDFDocumento23 páginasBacterias Gastrointestinales PDFprimeliaAún no hay calificaciones
- ShigellaDocumento24 páginasShigellamarce battiAún no hay calificaciones
- 03 LípidosDocumento7 páginas03 LípidosbeabergamoAún no hay calificaciones
- XLD PDFDocumento4 páginasXLD PDFPaolo Gambini ValderramaAún no hay calificaciones
- Manual - Laboratorio de Microbiologia de Suceptibilidad A Los Nos y de BacteriasdDocumento410 páginasManual - Laboratorio de Microbiologia de Suceptibilidad A Los Nos y de Bacteriasdquimico clinico 2795% (39)
- Músculo diafragma: función y descripción enDocumento1 páginaMúsculo diafragma: función y descripción enprimeliaAún no hay calificaciones
- CampylobacterDocumento23 páginasCampylobacterprimeliaAún no hay calificaciones
- Digestivo. GeneralidadesDocumento32 páginasDigestivo. GeneralidadesprimeliaAún no hay calificaciones
- Lipidos - FosfolipidosDocumento27 páginasLipidos - FosfolipidosprimeliaAún no hay calificaciones
- Bacterias Gastrointestinales PDFDocumento23 páginasBacterias Gastrointestinales PDFprimeliaAún no hay calificaciones
- Identificación bacteriana: métodos fenotípicosDocumento17 páginasIdentificación bacteriana: métodos fenotípicosprimeliaAún no hay calificaciones
- Fisiologia Muscular en La Biomecanica.Documento17 páginasFisiologia Muscular en La Biomecanica.Omar GonzalezAún no hay calificaciones
- DeltoidesDocumento1 páginaDeltoidesprimeliaAún no hay calificaciones
- Manual Instrumental Yami 150807170847 Lva1 App6892Documento42 páginasManual Instrumental Yami 150807170847 Lva1 App6892TamaraAún no hay calificaciones
- Manual de Instrumental QuirúrgicoDocumento462 páginasManual de Instrumental QuirúrgicoprimeliaAún no hay calificaciones
- Troncoenceflico 120809132414 Phpapp01Documento89 páginasTroncoenceflico 120809132414 Phpapp01primeliaAún no hay calificaciones
- RIUNNE - Trabajo Copado de Contaminantes PDFDocumento101 páginasRIUNNE - Trabajo Copado de Contaminantes PDFprimeliaAún no hay calificaciones
- Técnicas de SiembraDocumento8 páginasTécnicas de SiembraprimeliaAún no hay calificaciones
- Elcerebelo 100827233243 Phpapp02Documento14 páginasElcerebelo 100827233243 Phpapp02Edwin Chávez HuamánAún no hay calificaciones
- Medios de CultivoDocumento26 páginasMedios de CultivoprimeliaAún no hay calificaciones
- BacteriologíaDocumento25 páginasBacteriologíaprimeliaAún no hay calificaciones
- Reacciones Cualitativas Sobre Pigmentos de PlantasDocumento9 páginasReacciones Cualitativas Sobre Pigmentos de Plantasyamily888Aún no hay calificaciones
- Informe 4 Derivados de Alcaloides TropánicosDocumento9 páginasInforme 4 Derivados de Alcaloides TropánicosHedén Vega100% (1)
- Informe #1. Caracterización Aire Interior ColmenaDocumento21 páginasInforme #1. Caracterización Aire Interior ColmenaSergio B.Aún no hay calificaciones
- 1.3.4 1,3 PropanodiolDocumento9 páginas1.3.4 1,3 PropanodiolLaura Calvo DevesaAún no hay calificaciones
- Cs Naturales 5° básicoDocumento4 páginasCs Naturales 5° básicoMarcia Igor100% (2)
- Ejercicios Acido Base CinetDocumento3 páginasEjercicios Acido Base CinetfakesasoAún no hay calificaciones
- Ejercicios Balance Reacción QuímicaDocumento6 páginasEjercicios Balance Reacción QuímicaJorge Mario Arrieta RodríguezAún no hay calificaciones
- CueroDocumento2 páginasCueroTeFany Csc MousesitaAún no hay calificaciones
- Clorhexidina y CetrimidaDocumento29 páginasClorhexidina y Cetrimidarobertogarci50% (2)
- Parcial 2 Analisis InstrumentalDocumento6 páginasParcial 2 Analisis InstrumentalJONATHAN SEBASTIAN MENDIETA RAMIREZAún no hay calificaciones
- Presentación Nitacion de BencenoDocumento12 páginasPresentación Nitacion de BencenoChemiita LfcAún no hay calificaciones
- Los Hidrocarburos Liquidos Gaseosos y SolidosDocumento11 páginasLos Hidrocarburos Liquidos Gaseosos y Solidosenrique33% (3)
- Procesos industriales y química ambientalDocumento151 páginasProcesos industriales y química ambientalNatalia Leyton ValenzuelaAún no hay calificaciones
- Evidencia Integradora ADocumento13 páginasEvidencia Integradora ABere HuertaAún no hay calificaciones
- Programa Competencias OI 2021Documento9 páginasPrograma Competencias OI 2021ERICKA ODILY FLORES VALENCIAAún no hay calificaciones
- Práctica N°3 Elaboración de Pomada AntirreumáticaDocumento6 páginasPráctica N°3 Elaboración de Pomada AntirreumáticaZayda Juchatuma100% (1)
- Marchas AnalíticasDocumento11 páginasMarchas AnalíticasJesus Martin Briones Inciso100% (1)
- Plan para Remediales 2018-2019Documento4 páginasPlan para Remediales 2018-2019Byron PuetateAún no hay calificaciones
- Síntesis de ProteínasDocumento13 páginasSíntesis de Proteínasabril daffaraAún no hay calificaciones
- Alcaloides PDFDocumento3 páginasAlcaloides PDFtere_figueroaAún no hay calificaciones
- Rizósfera y absorción por simbiosis: efecto en la nutrición vegetalDocumento11 páginasRizósfera y absorción por simbiosis: efecto en la nutrición vegetalluis kunAún no hay calificaciones
- Medios de CultivoDocumento31 páginasMedios de CultivoMelanie AshleyAún no hay calificaciones
- La Membrana MagicaDocumento2 páginasLa Membrana MagicaRuben MP100% (1)
- TesisDocumento175 páginasTesisSuggeis ChAún no hay calificaciones
- Célula eucariota: organelos y funcionesDocumento15 páginasCélula eucariota: organelos y funcionesgabyh12100% (2)
- Tinto Pretratamiento F Proteicas LanaDocumento46 páginasTinto Pretratamiento F Proteicas Lananicole talavera zeladaAún no hay calificaciones
- Reportaje CeresitaDocumento1 páginaReportaje CeresitadirecminAún no hay calificaciones
- Reacciones de Los AlquinoDocumento9 páginasReacciones de Los AlquinoIrvin AdunaAún no hay calificaciones
- Acción de La Lipasa Pancreática - Lab BioquimicaDocumento10 páginasAcción de La Lipasa Pancreática - Lab BioquimicaLilibeth Juliana Rojas Carrillo100% (2)