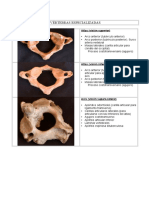
Documentos de Académico
Documentos de Profesional
Documentos de Cultura
Apuntes Completos de La Asignatura
Cargado por
ALBERTO GALLEGODerechos de autor
Formatos disponibles
Compartir este documento
Compartir o incrustar documentos
¿Le pareció útil este documento?
¿Este contenido es inapropiado?
Denunciar este documentoCopyright:
Formatos disponibles
Apuntes Completos de La Asignatura
Cargado por
ALBERTO GALLEGOCopyright:
Formatos disponibles
lOMoARcPSD|4316946
Apuntes completos de la asignatura
Organografía Microscópica Humana (Universidad Complutense Madrid)
StuDocu no está patrocinado ni avalado por ningún colegio o universidad.
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Curso 2015-2016
Fátima Carrasco
Marta Lloret
Ainhoa Lorenzo
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
ÍNDICE
APARATO RESPIRATORIO………...…………………………………………………3
APARATO CARDIOVASCULAR ………………………………………………...…31
SISTEMA INMUNE O LINFOIDE……………………………………………………57
SISTEMA TEGUMENTARIO………………………………………………………...81
MAMA………………………………………………………………………………..110
APARATO DIGESTIVO…………………………………………………………….122
ANEJOS AL DIGESTIVO…………………………………………………………...160
APARATO URINARIO……………………………………………………………...187
APARATO GENITAL MASCULINO………………………………………………210
APARATO GENITAL FEMENINO………………………………………………...245
PLACENTA…………………………………………………………………………..279
OÍDO…………………………………………………………………………….……289
OJO…………………………………………………………………………………....313
SISTEMA NERVIOSO……………………………………………………………….351
GLÁNDULA PITUITARIA O HIPÓFISIS………………………………………….405
GLÁNDULA PINEAL O EPÍFISIS…………………………………………………419
TIROIDES…………………………………………………………………………….425
PARATIROIDES……………………………………………………………………..431
GLÁNDULAS SUPRARRENALES…………………………………………………435
PÁNCREAS ENDOCRINO…………………………………………………………..445
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
APARATO RESPIRATORIO
x Permite el intercambio de gases
x Percepción de olores
x Fonación
Partes
x Porción conductora: Nariz, faringe, laringe, tráquea y árbol bronquial; (boca
espiración forzada)
x Porción respiratoria: bronquios respiratorios, alveolos
x Un mecanismo de ventilación, controlado por los movimientos inspiratorio y
espiratorio de la caja torácica, la cual está movida por el diafragma.
PORCIÓN CONDUCTORA
Funciones
x Conducción del aire a los pulmones
x Acondicionamiento
o Limpieza
o Atemperamiento
o Humectación
x Quimiorrecepción
x Defensa del organismo
x Fonación
x Vocalización
x Separación de las vías digestiva y respiratorio
Nariz
Piel con pelos finos y glándulas sebáceas grandes, tejido conjuntivo y músculo.
Está dividida en dos por el tabique nasal óseo-cartilaginoso. Posee unas entradas
anteriores, narinas, y otras posteriores, coanas.
Cada cavidad nasal, separada de la otra por el tabique, consta de vestíbulo,
porción respiratoria y área olfatoria. El aire penetra a través de las narinas, cuya
superficie externa está revestida de epitelio escamoso queratinizado. En el vestíbulo, el
epitelio se vuelve no queratinizado.
Sus paredes son irregulares (cornetes), que aumentan la superficie y crea
turbulencias en el flujo aéreo para favorecer el contacto con la mucosa (epitelio +
lámina propia) y el moco. Se produce una precipitación turbulenta que aumenta la
eficacia en la filtración del aire.
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Parte frontal: epitelio plano estratificado queratinizado con pelos rígidos grandes
(vibrisas), abundantes glándulas sebáceas (independientes) y sudoríparas.
Continuación de la epidermis.
x Parte posterior: epitelio respiratorio, pseudoestratificado ciliado con
caliciformes. Posee moco, que tiene una capa periciliar y una capa de gel
mucoso. Está en la porción posterior del vestíbulo, y no es regiones expuestas
(cuerdas vocales, cornetes...).
Los cilios desplazan la capa de gel (1cm/min).
La hidratación determina el grado de viscosidad y las propiedades elásticas,
esenciales para la eliminación
o El tabaquismo va a alterar el patrón de batido sincrónico y reduce la
frecuencia de batido ciliar. Se altera la eliminación del moco y se
produce tos crónica. El número de células ciliadas disminuye. Hay
metaplasia (se origina epitelio plano estratificado).
o La enfermedad de Kartagener los cilios son inmóviles. Hay sinusitis
crónica, bronquitis crónica, bronquiectasias (dilatación permanente de
uno o varios bronquios). En varones provoca esterilidad pues se ven
afectado los cilios de todas las células.
El moco es segregado por células caliciformes, células de Clara y glándulas. Se
compone de 97% de agua y 3% de mucinas, proteínas antimicrobianas, sales,
lípidos y restos de células. Las mucinas más importantes son las MUC5AC
(caliciformes, Clara), MUC5B (caliciformes, Clara y glándula). La síntesis es
continua.
Las vibrisas se encargan de filtrar las partículas y el moco atrapa el polvo e
impide la deshidratación. A continuación el aire se transporta hasta la faringe. Reflejo
del estornudo provocado por nervios en esta zona.
Epitelio respiratorio
Hay moco formado por células caliciformes y glándulas de la lámina propia, por
tanto, esta mucosa es húmeda. En pacientes con traquotomía el aire pasa directamente y
no se humidifica, formándose cortes.
Bajo el epitelio hay una lámina basal engrosada. En la lámina propia hay células
libres y fibras elásticas, con presencia de melanocitos. Hay glándulas serosas y
mucosas, con gran vascularización. Apoya sobre periostio/pericondiro. También se
encuentran leucocitos, mastocitos y plasmáticas. En la rinitis alérgica los mastocitos de
los capilares permites el paso del plasma al conjuntivo edematoso.
Este epitelio se localiza en el suelo y las paredes laterales de la mucosa que
recubre los cornetes medio e interior, que se denomina pituitaria roja por su rica
vascularización:
x Capilares superficiales en hilera perpendiculares al flujo de aire. El flujo a
contracorriente permite atemperar el aire.
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Plexo venoso superficial, tejido eréctil (parecido al cuerpo cavernoso).
x Abundante anastomosis arteriovenosas de trayecto enrollado.
x Las paredes venosas tienen haces musculares gruesos en espiral (esfínter).
Además, hay estancamiento en las sinusoides cavernosas y apertura y cierre de
anastomosis. Esto produce que el flujo de sangre sea variable. Cuanto mayor sea
la mucosa menor será el flujo de aire.
x La inervación simpática (vasoconstrictora) y parasimpática (vasodilatación y
secreción), de modo que van a influir en el tamaño de la mucosa).
Obstrucción nasal Æ agentes adrenérgicos.
En la nariz se produce el atemperamiento del aire y disolución y absorción de
vapores hidrosolubles.
Epitelio olfativo
En el techo y las paredes laterales superiores está la mucosa que recubre el
cornete superior, la pituitaria amarilla, por la lipofucsina de células epiteliales y
glandulares. El epitelio es olfatorio y la vascularización se reduce.
El epitelio olfatorio es psudoestratificado (50 micras) y ciliado, sin caliciformes,
pero con células de sostén, basales y olfativas.
x Células de sostén: Sostén mecánico y metabólico de células olfativas (glía). Son
prismáticas y tienen microvellosidades. Su núcleo es oval central, hacia la zona
apical. Abundantes REL, RER y lipofucsina. Tiene uniones estrechas y
adhesivas entre ellas y adhesivas con células olfativas.
x Células basales: Células madres que aseguran la renovación de las células de
sostén y olfativas. No alcanzan la luz, con núcleo heterocromático, pertenecen a
la zona basal del epitelio. Están en contacto con el axón de las células olfativas.
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Células olfativas: 107 neuronas bipolares, con soma central ensanchado y
prismático. Están entre las células de sostén y se renuevan rápidamente. Con la
edad disminuye el número y se pierde agudeza olfativa.
o Las dendritas están en la zona apical y alcanza la superficie y forma una
vesícula olfativa (maza). En esa maza se observan 10-20 quinocilios (200
micras) con disposición radial dentro de la capa de moco.
o La otra prolongación es el axón, basal y mucho más delgado. Atraviesa
la lámina basal y se une a otros. Forma el par craneal I y se dirige a la
lámina cribosa del etmoides hasta el bulbo olfatorio.
La lámina propia está formado por tejido conjuntivo laxo con vasos sanguíneos
y glándulas de Bowman. Las glándulas de Bowman son túbuloalveolares, que se abre a
la superficie y con secreción serosa. Su secreción mantiene húmeda la mucosa
permitiendo que las sustancias olorosas se disuelvan y asegura el flujo constante que
lave la mucosa para percibir nuevos olores.
El líquido secretado contiene la proteína ligadora de moléculas de olor (OBP),
con una elevada afinidad para unirse con un gran número de moléculas olorosas. La
OBP transporta las sustancias olorosas a los receptores presentes en la superficie de los
cilios modificados y las retira una vez percibidas. Además, el producto de secreción de
las glándulas de Bowman contiene sustancias protectoras, como la lisozima y la
inmunoglobulina A (IgA), secretadas por las células plasmáticas.
Mecanismo olfativo
Unos 1000 genes codifican para receptores olfativos (1% del total de genes).
Cada células olfativa solo expresa uno de esos genes y expresa un receptor es la
membrana de los quiliocilios.
La OBP se une a las sustancias odoríferas y juntas se dirigen al receptor. El
receptor está acoplado a proteína G. La OBP se retira y se recicla, se produce una
despolarización de membrana, produciéndose un impulso nervioso.
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Senos paranasales
Son oquedades que se encuentran excavadas en diferentes huesos (maxilar o
antro de Highmore (es el mayor), etmoidal, frontal, esfenoidal…). Los senos se revisten
de un epitelio cilíndrico pseudoestratificado ciliado. Además, contienen aire dentro de
los huesos del cráneo. Aparecen en número par, y su tamaño es variable.
Los senos paranasales están tapizados por mucosa epitelial respiratoria, con
epitelio menos grueso y con menos células caliciformes que la cavidad nasal. Existen
menos glándulas que en el resto del epitelio respiratorio y no se observa tejido eréctil
como en la pituitaria roja.
El epitelio es más fino y, al haber pocas glándulas, el moco que lo cubre procede
de glándulas de fuera de los senos.
Los cilios llevan el moco a la cavidad nasal. Frecuentemente los senos presentan
un problema clínico debido a la mucosa; provocando inflamación e infección
(SINUSITIS). Por ello es importante el batido de los cilios, para conseguir movilidad
continua de la capa de moco hacia la cavidad nasal.
Los senos se comunican con la cavidad nasal a través de orificios revestidos de
un epitelio parecido al presente en la cavidad nasal. Los senos etmoidales desembocan
por debajo del cornete superior y el seno maxilar lo hace por debajo del cornete medio.
Faringe
Es el punto de cruce de las vías del aparato digestivo y el respiratorio. Actúa
como cámara de resonancia para la fonación (amplificando los sonidos). Se divide en:
x Epifaringe o rinofaringe
x Mesofaringe u orofaringe
x Hipofaringe o laringofaringe
Nasofaringe
Presenta un epitelio continuo igual al de la cavidad nasal. El epitelio se modifica
a plano estratificado no queratinizado cuando nos acercamos a la mesofaringe.
Lateralmente a la nasofaringe se abren las trompas de Eustaquio.
Luego bajo el epitelio hay una lámina propia en la que destaca MALT (tejido
linfoide), donde se localiza el anillo de Waldeyer y la amígdala faríngea, impar y media
y cuya inflamación causa vegetaciones al obstruirse el paso del aire, lo que obliga a
respirar por la boca.
Laringe
Es la parte del aparato respiratorio que une la faringe con la tráquea. Presenta
una longitud de 4-5 cm. Y está cubierta por una pieza móvil: la epiglotis.
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
La entrada está cubierta por la epiglotis (zona con movilidad, cubierta que se
abre y cierra). Se divide en 3 tramos:
x La supraglotis se inicia en la epiglotis, el primer par pliegues vocales y los
ventrículos laríngeos o de Morgagni.
x La glotis la forman los segundos pliegues vocales (cuerdas), y las comisuras
anterior y posterior.
x La subglotis es la zona de la laringe que se extiende bajo los segundos
pliegues vocales, hasta el cartílago cricoides.
Mucosa laríngea
El epitelio respiratorio de la laringe presenta una capa de moco que es batida
hacia la faringe. Debajo encontramos una lámina propia laxa, con abundantes glándulas
seromucosas y folículos linfoides. En esta lámina hay una gran cantidad de linfocitos
que pueden formar nódulos.
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Como se ha mencionado, la lámina propia no es toda igual, es más laxa en la
zona superficial. La lámina propia TC tiende a hacerse más densa conforme bajamos,
tiene muchas células cebadas y esto es idóneo para reacciones de defensa
(hipersensibilidad causando crups y falso crup que puede llegar a provocar asfixia en
niños pequeños). Por ello no es raro encontrar de manera recurrente mastocitos.
CRUP es el proceso inflamatorio que estrecha las vías respiratorias, hace un
sonido muy característico: estridor inspiratorio. En niños es fácil encontrarlo.
Cartílagos
En la faringe encontramos cartílagos rodeados por abundante pericondrio
de conjuntivo denso que une además las placas cartilaginosas. Algunos de los
cartílagos son pares (aritenoides, corniculados y cuneiformes) y otros impares
(tiroides, cricoides, epiglotis). Son cartílagos hialinos, excepto la epiglotis, que es
cartílago elástico.
La epiglotis actúa de tapa de esta laringe, tiene dos caras claramente
diferenciadas: la superior esta tapizada por un epitelio plano estratificado no
queratinizado y la inferior sí tiene epitelio respiratorio y glándulas.
Todos estos cartílagos calcifican con la edad.
Músculos esqueléticos
Los músculos de la laringe pueden ser intrínsecos o extrínsecos y están entre los
cartílagos.
Los músculos intrínsecos permiten la apertura y cierre de la glotis. Se
relacionan con las placas cartilaginosas. Repercuten en el diámetro de la glotis debido a
su contracción. Además, cuando se contraen se tensan las cuerdas vocales y se modula
la fonación.
Los músculos extrínsecos no actúan entre las placas cartilaginosas y por tanto
en la glotis sino que unen la laringe con estructuras próximas y su función es la
elevación o descenso de esta laringe (lo que es importante pues estamos muy cerca del
cruce de las vías respiratorias y digestiva) para que el aire y el bolo alimenticio pasen
por el sitio correcto. Luego repercute en la longitud de la laringe.
Reflejo de la tos
Hay una inervación aferente que produce el reflejo de la tos. Esto es por una
inspiración forzada acompañada por un cierre brusco de la glotis, seguida por una
contracción de la musculatura abdominal e intercostal y siendo el aire expulsado a una
gran velocidad (180km/h). Se produce una apertura brusca de la epiglotis y de las
cuerdas vocales, saliendo el aire de manera brusca y súper fuerte.
10
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Pliegues de la laringe
El primer par de pliegues vocales son las cuerdas vocales falsas o
ventriculares. Están cubiertas de epitelio respiratorio, si bien es frecuente
encontrar parches de epitelio plano estratificado no queratinizado.
La lámina propia es laxa y presenta glándulas seromucosas que pueden
introducirse en la epiglotis y lubrican los pliegues. No presenta musculatura
intrínseca. No está implicado en la fonación y su función es eminentemente
protectora.
Entre estos pliegues y los siguientes, encontramos una zona hendida
conocida como ventrículo laríngeo o de Morgagni, que se prolonga en un sáculo (zona
más profunda y estrecha). Está tapizado por epitelio respiratorio, con una lámina propia
laxa con glándulas seromucosas. Participa en la resonancia, intensificando el sonido en
las cuerdas vocales verdaderas. Si hay musculatura: músculo ventricular.
La secreción va a cubrir el primer par de pliegues y actúa como lubricante en
esta primera sección. Tiene función de protección y no colabora en la fonación.
Cuerdas vocales verdaderas
Ocupada por cuerdas vocales verdaderas que no tienen epitelio respiratorio sino
epitelio estratificado no queratinizado que ofrece mejor protección a esta zona frente a
la abrasión que supone el paso del aire (y el fuerte estrés mecánico), en él encontramos
melanocitos y bajo el cual tenemos una lámina propia (TC) con tres zonas:
x Superficial: también llamado espacio de Reinke.
x Media.
x Profunda.
x Músculo vocal o tiroartenoideo (musculatura esquelética).
El espacio de Reinke es TC muy laxo donde no hay glándulas ni vasos linfáticos
y que está ocupado por una matriz (Sust. Fundamental) en la que hay fibras elásticas. En
cambio, el ligamento vocal (capa media y profunda) está ocupado por TC rico en fibras
elásticas llegando a encontrar haces bastante densos de estas. Esta zona también es
propicia a los falsos crup, porque encontramos muchos mastocitos, produciéndose
edemas y obstrucción faríngea.
Funcionalmente las cuerdas vocales tienen 2 zonas diferenciadas:
x Cubierta: epitelio + espacio de Reinke
x Centro o Núcleo: Lig vocal + musculatura vocal.
Son distintas porque la cubierta es enormemente flexible mientras que el centro
es rígido. Además, la cubierta está completamente recubierta de secreción glandular (y
11
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
como no hay glándulas aquí, esta secreción viene de las cuerdas vocales falsas, es decir,
de las glándulas que están en la epiglotis).
El ligamento vocal está formado por elementos elásticos, abundantes y formando
haces paralelos. Hay muchos mastocitos. Si hay demasiados con frecuencia, reacciones
de hipersensibilidad. El núcleo dentro del cual se encuentra la musculatura vocal y el
ligamento vocal, da rigidez.
Fonación
Las cuerdas vocales verdaderas son responsables de la fonación sufriendo
primero una abducción y como consecuencia empiezan a vibrar y se acaban acercando
dejando un espacio mínimo, estrecho y lineal. Este ciclo se repite una y otra vez y está
provocado por el aire que sale que choca contra las cuerdas vocales provocando su
fonación. Lo que vibra es la cubierta que es la que tiene flexibilidad.
En definitiva, tenemos una estructura par de tal manera que cuando el aire choca
se produce un movimiento horizontal y una onda vertical (onda de la cubierta). Este
ciclo debe estar acompañado por la contracción de la musculatura intrínseca que hace
que el centro de las cuerdas vocales sea aún más rígido.
Hay diferentes sonidos dependiendo de la flexibilidad de la cubierta y la rigidez
del centro. Cuanto más rígido sea el centro, más rápidamente viaja la onda y hace que se
aumente el tono. Los tonos se diferencian por el diámetro de la glotis y el grado de
distensión de la cuerda vocal. A más relajación de las cuerdas, más profundo es el tono.
Tras la pubertad la laringe se alarga más en el varón y es por eso por los que los
tonos siempre van a ser más profundos que los de la mujer.
Vocalización
Intervienen el paladar blando, la lengua, los labios y los dientes, permiten formar
diferentes sonidos.
Apunte clínico
x Laringitis: inflamación de cuerdas vocales que impiden correcta vibración
x Disfonía: voz ronca o susurrante
x Paraqueratosis en cuerdas vocales verdaderas por exposición a irritantes
(parches de queratina en el epitelio)
x Pólipos, nódulos y edemas crónicos.
x Cáncer laríngeo
12
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Tráquea
Tubo de 10 x 2 cm, rígido y flexible (puede cambiar su longitud y su diámetro)
con unos 15 o 20 anillos cartilaginosos (incompletos e hialinos) en forma de C cuyos
extremos están unidos por conjuntivo y miocitos. Entre los anillos hay ligamentos
formado por conjuntivo denso ricos en fibras elásticas puestas longitudinalmente
(ligamentos anulares), luego los extremos libres van a estar unidos por tejido conjuntivo
denso y musculatura lisa. Esto permite que el tubo varíe el diámetro y longitud
dependiendo de la musculatura y el TC.
Estrucutra histológica
Histológicamente, tiene un epitelio respiratorio apoyado sobre una Lámina
Basal más gruesa de lo normal y cubierto por moco. Debajo de esta tenemos una lámina
propia laxa (TC) con unidades linfoides (linfocitos y BALT) y finalmente, en una zona
más o menos profunda aparece una banda de fibras elásticas conocida como membrana
elástica que marcaría el límite de la lámina propia con la submucosa (Al microscopio no
vemos esta membrana así que vemos la lámina propia y la submucosa como un
conjunto). La submucosa es un tejido conjuntivo laxo que da paso a placas
cartilaginosas o a una banda muscular (según nos encontremos o no en una zona
de anillo de cartílago). Las placas cartilaginosas están unidas por músculo liso y a
continuación se dispone una túnica adventicia que es la capa más externa.
13
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Epitelio respiratorio y lámina basal
Es un epitelio ciliado (células ciliadas) con células caliciformes, endocrinas
(dispersas) y basales (células madre). La lámina basal es gruesa.
Hay muchos agentes, como el tabaco, que actúan como irritantes del aparato
respiratorio. En los fumadores crónicos se produce una tos crónica y en el caso
de la tráquea además se ve disminución en el número de células ciliadas,
aumento de las caliciformes e hipersecreción glandular (que es de diferente
composición a la habitual). Además, la lámina basal (llamada también lamina
reticular) que ya era de por sí más gruesa, tiende a ser aún más gruesa en el caso
de estos fumadores.
Los cilios del epitelio baten el moco a una frecuencia de 15 veces/segundo hacia
la faringe. El moco de desplaza a 1 cm/min.
x Lámina propia
La lámina propia es bastante laxa, altamente rica en células (del SI, fibroblastos,
linfocitos…), con abundantes fibras elásticas que en la parte inferior convergen
para formar la membrana elástica y con abundantes unidades linfoides (BALT).
x Submucosa
La submucosa, debajo de la membrana elástica (que separa la lámina propia de
la submucosa), es un TC laxo con grandes glándulas seromucosas, muchas fibras
elásticas y por ella discurren los grandes vasos así como toda una red de vasos
linfáticos.
x Cartílago o banda muscular
Los anillos de cartílago son importantes para evitar que este tubo se colapse
cuando sale el aire (espiración), gracias a las placas. Como el cartílago hialino
tiende a calcificarse con los años la tráquea pierde flexibilidad. Por otro lado, en
las zonas sin cartílago encontramos músculo liso.
x Capa adventicia
Está asociada a este tubo en las estructuras próximas y donde encontramos los
vasos y nervios de mayor calibre. Se continúa con el tejido conjuntivo del cuello
y del mediastino, fijando la tráquea a estas estructuras.
14
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Árbol bronquial
La tráquea termina abriéndose con ramificaciones dicotómicas en un árbol
bronquial que se forma en el periodo embrionario pudiendo observarlo en el cuarto mes
de vida. Así, partiendo de la tráquea tenemos dos bronquios primarios extrapulmonares
que entran en el pulmón por la raíz pulmonar que tiene mucho TC y grandes vasos. Una
vez dentro estos bronquios se ramifican y tenemos cinco bronquios secundarios o
lobares (3 en el pulmón derecho y 2 en el izquierdo) que se siguen ramificando y dan
lugar a los bronquios terciarios o segmentarios de los cuales tenemos 8 en el pulmón
izquierdo y 10 en el derecho.
En la ramificación nº 12 tenemos el bronquiolo que se ramificará hasta 4 veces
más y en la ramificación nº 16 tenemos el bronquiolo terminal de los cuales hay unos
65000. Hasta aquí llega la porción conductora.
Durante todo este proceso de ramificación tanto la pared como el diámetro del
tubo ha disminuido.
Cuando estudiamos el árbol bronquial hablamos de segmentos
broncopulmonares que sería cada bronquio segmentario junto con todo el parénquima
asociado a él, con su propia inervación y vascularización y estroma, de modo que la
extirpación (en caso de tumores por ejemplo) se realiza a nivel de estos segmentos.
Bronquios
La tráquea se divide en dos ramificaciones que forman los bronquios principales
o primarios. Desde el punto de vista anatómico, estas divisiones se conocen
simplemente como bronquios fuente izquierdo y derecho, siendo el derecho mucho más
amplio y corto que el izquierdo.
Los bronquios disponen de varias capas, que de dentro hacia afuera son: mucosa,
muscular lisa, submucosa (con placas cartilaginosas) y adventicia. Están muy
vascularizados.
Mucosa
En la mucosa hallamos un epitelio respiratorio cuya altura disminuye a medida
que las ramificaciones bronquiales avanzan, con varios tipos celulares:
15
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Linfocitos
x Mastocitos
x Células ciliadas: similares a las ya vistas
x Células caliciformes: en un número muy abundante.
x Células madre: situadas cerca de los nódulos
x Células endocrinas, de gránulos pequeños o de Kulchitsky: se trata de los
equivalentes respiratorios de las células enteroendocrinas del aparato digestivo.
Como estas últimas, se trata de células pertenecientes al sistema APV difuso. En
su citoplasma hay gránulos de centro denso limitados por membrana, que
contienen hormonas y neurotransmisores. Aunque normalmente aparecen
individualizadas, dispersas por el epitelio bronquial, también las podemos hallar
en grupos o cúmulos denominados cuerpos neuroepiteliales.
Los cuerpos neuroepiteliales se hallan normalmente en los puntos de
ramificación bronquial, y tienen una gran importancia clínica debido a su
relación con carcinomas y carcinoides. Estas estructuras son quimiorreceptores
sensibles a los cambios en la presión parcial de oxígeno, y por ello sinaptan con
fibras aferentes, lo cual permite regular la presión pulmonar y la apertura de las
vías respiratorias.
La lámina propia bronquial es un tejido conjuntivo laxo, con abundantes fibras
elásticas. Es muy celular, y contiene linfocitos abundantes, que se infiltran con
frecuencia en el epitelio constituyendo tejido difuso o se agrupan en forma de nódulos
(valus), y tienen gran importancia debido a su participación en reacciones inflamatorias
y alérgicas. También encontramos células M. La lámina propia está altamente
vascularizada, conteniendo en muchos casos VEA, y tiene también un alto contenido en
glándulas (que, al igual que la altura del epitelio, van disminuyendo conforme las
ramificaciones bronquiales avanzan). Si estas glándulas son seromucosas, a su alrededor
encontraremos células mioepiteliales para ayudar a expulsar la secreción que contienen.
Muscular
Es una capa continua de músculo liso en los bronquios mayores. Aun así, su
disposición es helicoidal, en espiral, de manera que en un corte histológico transversal
puede parecer que esta continuidad no es tal, pero solo se debe a su organización. La
contracción de este músculo mantiene el diámetro adecuado de la vía aérea, y a ella se
acopla la mucosa, que se pliega cuando el músculo se contrae.
Está regulada por el sistema nervioso autónomo: la división simpática aumenta
el diámetro bronquial e inhibe la secreción glandular, mientras que la parasimpática
disminuye el diámetro bronquial y aumenta la secreción mucosa. Llegados a este punto,
se ha de destacar la alta concentración de VIP, un neurotransmisor cuyo origen a este
nivel concreto no está claro, pero que se encarga de provocar broncodilatación.
A medida que vamos avanzando en las ramificaciones, la capa muscular va
disminuyendo, pero en menor proporción que otras, de manera que proporcionalmente
el componente muscular va aumentando.
16
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Submucosa
Permanece como un tejido conjuntivo bastante laxo, que en los bronquios
mayores contiene glándulas y tejido adiposo. Podemos hallar rodeándola placas
cartilaginosas, cada vez menores.
Adventicia
Similar a la traqueal.
En resumen, a medida que avanzan las ramificaciones:
x Disminuyen: altura del epitelio, contenido en glándulas de la submucosa y la
lámina propia, contenido en placas cartilaginosas
x Aumenta: proporción de músculo liso bronquial
Bronquiolos
Hablamos de bronquiolos a partir de la duodécima división (D12) del árbol
bronquial. Los bronquiolos son vías de conducción aérea cuyo diámetro es < 1 mm.
Mucosa
Ya no hallamos un epitelio respiratorio, sino un epitelio prismático simple ciliado.
En cuanto a los tipos celulares:
x Células ciliadas: en un principio son muy numerosas, pero su número y altura
van disminuyendo a medida que aumentan las ramificaciones.
x Células caliciformes: son escasas, ante la existencia de otros tipos celulares
encargados de la secreción, y van desapareciendo conforme se ramifican los
bronquiolos. Sin embargo, en presencia de agentes tóxicos o nocivos, como es el
caso de los fumadores, las células caliciformes siguen existiendo a este nivel,
con lo que hallamos secreciones que no deberían de estar.
x Células de Kulchitsky y cuerpos neuroepiteliales
x Células de Clara: son células cúbicas no ciliadas que exhiben una prominencia
característica en forma de seta o cúpula en su extremo superior. Sus orgánulos
están muy desarrollados: poseen un RER basal, REL apical y un aparato de
Golgi lateral muy desarrollado. Estas células, que constituyen entre un 80 y un
85 % de las células del epitelio bronquiolar, poseen gránulos de secreción, ya
que son las encargadas de secretar un agente tensioactivo de naturaleza
lipoproteica que impide la adhesión luminal si la pared de la vía aérea se
colapsa, en particular durante la espiración. Además, se encargan de sintetizar
las mucinas MUC5AC y MUC5B.
Clínica: fibrosis quística
Las células de Clara están vinculadas con el transporte de iones cloruro, lo cual
se halla íntimamente relacionado con la fibrosis quística. Esta enfermedad
pulmonar crónica sigue un patrón obstructivo, y afecta principalmente a los
17
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
niños y a los adultos jóvenes. Es un
trastorno autosómico recesivo que
afecta a la viscosidad de la
secreción de las células de Clara.
La causa es un defecto genético en
la proteína canal para el ion cloruro
que conduce a un transporte
anormal a través del epitelio.
Además, disminuye la secreción de
ion bicarbonato y aumenta la de
ion sodio, lo cual crea secreciones
demasiado espesas ricas en sodio
(sudor salado es un signo) y pobres
en cloro, cuyas mucinas están
enmarañadas, lo cual hace muy
difícil mover el moco. Además de
ser poco móvil y obstruir las vías
aéreas, el moco atrae a
microorganismos patógenos.
También se pueden observar
alteraciones pancreáticas.
La lámina propia del epitelio bronquiolar es continua con la de los bronquios: se
trata de un tejido conjuntivo altamente vascularizado y rico en fibras elásticas. Sin
embargo, existe una gran diferencia entre ambas láminas: la bronquiolar ya no dispone
de glándulas.
Muscular
Se trata de una capa de músculo liso denominado músculo de Reissesen,
relativamente gruesa. No es continuo, pero posee abundantes desmosomas.
Los miocitos lisos del músculo de Reissesen son sensibles a cambios en la
presión parcial de oxígeno: cuando la presión parcial de oxígeno es baja el músculo se
dilata y cuando es alta se contrae. Ello hace que el aire inspirado se distribuya de forma
uniforme en los alveolos: si uno de los bronquiolos está peor ventilado, su presión
parcial de oxígeno será más baja, y ello contribuirá a la relajación del músculo liso y por
tanto a la broncodilatación.
Al igual que la contracción de los bronquios, la del músculo liso bronquiolar está
regulada por el sistema nervioso autónomo: la división simpática aumenta el diámetro
bronquial e inhibe la secreción glandular, mientras que la parasimpática disminuye el
diámetro bronquial y aumenta la secreción mucosa. También aquí actúa el VIP, un
neurotransmisor cuyo origen a este nivel concreto no está claro, pero que se encarga de
18
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
provocar broncodilatación. La broncoconstricción sostenida a este nivel resulta mucho
más peligrosa, pudiendo causar la asfixia.
La submucosa en los bronquiolos, al igual que las placas cartilaginosas, es
INEXISTENTE, aunque sí que podemos hablar de adventicia.
Clínica: enfermedad pulmonar obstructiva crónica (EPOC)
La EPOC es un cuadro clínico consistente a grandes rasgos en una combinación
de enfisema pulmonar y asma. El enfisema se caracteriza por un agrandamiento
permanente de los espacios aéreos distales con respecto al bronquiolo terminal, ya que
el pulmón no recupera la distensión a la que se ve sometido en inspiración. Se asocia a
una desaparición irreversible de las fibras elásticas pulmonares debido a un aumento de
la actividad elastasa derivada de macrófagos y neutrófilos (por ejemplo, en fumadores
crónicos) y a una disminución de la actividad de la alfa-1-antitripsina, enzima encargada
de controlar la elastasa. Tenemos dos tipos de enfisema:
1. Centroacinar: comienza en los lobulillos, con lo que unos se ven afectados y
otros no.
2. Paraacinar: empieza por los bronquiolos.
El asma es una inflamación crónica de las vías respiratorias. Esta enfermedad
conlleva un aumento del número de células libres y la secreción de interleucinas (entre
ellas, destaca la 5). Asimismo, se produce una hipersecreción glandular (la secreción es
espesa y difícilmente eliminable) y aumenta la permeabilidad de los vasos sanguíneos,
lo cual da lugar a la aparición de edema. Como principal signo hallamos
bronconstricción exagerada y persistente, aunque reversible. En la luz de los
bronquiolos de asmáticos podemos encontrar cristales de Charcot-Leyden, fácilmente
distinguibles debido a su alta eosinofilia.
Bronquiolos terminales
Su pared es continua todavía.
x Epitelio: en él sólo encontramos células de Clara, ya no tiene ni siquiera
caliciformes. Su lámina propia es escasa y posee fibras elásticas.
x Muscular: es espiral, y solo tiene inervación parasimpática
Anexo: conceptos
x Lobulillo pulmonar: 1 bronquiolo terminal + región pulmonar asociada
(bronquiolos respiratorios, sacos alveolares y conductos alveolares, alveolos)
x Ácino: bronquiolo respiratorio + región pulmonar asociada (sacos alveolares y
conductos alveolares, alveolos)
19
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
PORCIÓN RESPIRATORIA
Bronquiolos respiratorios
Se ramifican hasta 3 veces. Sigue tratándose de un epitelio prismático simple, y
el número de cilios sigue disminuyendo, pero sus paredes están muy interrumpidas, de
hecho, lo que denominamos paredes son en realidad los bordes libres de los tabiques
interalveolares, ya que los bronquiolos respiratorios se abren al saco o atrio alveolar, al
que también se abren una gran cantidad de alveolos.
Alveolos
La extensión del espacio disponible para el intercambio gaseoso está acrecentada
por los alveolos, que son los espacios aéreos terminales del aparato respiratorio en los
que tiene lugar el intercambio gaseoso entre el aire y la sangre. Cada alveolo es una
cavidad que confluye en un saco alveolar:
x Conductos alveolares: vías aéreas alargadas que casi no tienen paredes, sino sólo
alveolos como sus límites periféricos.
x Sacos alveolares: espacios rodeados por cúmulos de alveolos que se abren a
ellos.
Los sacos alveolares suelen estar al final de un conducto alveolar, pero pueden
aparecer en toda su longitud. Los alveolos están separados unos de otros por una fina
capa de tejido conjuntivo que contiene capilares, denominada tabique alveolar o pared
septal, y perforada por los llamados poros de Kohn, que tienen entre 7 y 9 micras de
diámetro. Los poros permiten la circulación colateral alveolar, lo cual es bueno, porque
asegura una distribución uniforme del aire inspirado, pero puede resultar un
inconveniente debido a que facilita la diseminación de patógenos.
Los alveolos se forman entre el cuarto y el sexto mes de VIU, y su morfología en
el periodo fetal es esférica, hallándose llenos de líquido amniótico. En el momento del
nacimiento, se llenan de aire y se tornan angulosos (hay unos 20 millones), y a los 8
años de vida ya disponemos de unos 300 millones de alveolos, mucho más grandes.
20
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
21
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
En los alveolos tiene lugar la secreción del surfactante, secreción grasa que evita
el colapso pulmonar (agente tensioavctivo). Es un epitelio plano simple formado por
neumocitos tipo I (90-95% superficie) y neumocitos tipo II (5-10%). Debajo se
encuentran la lámina basal y el intersticio.
Los neumocitos tipo I son claros, células planas con escasos orgánulos, unidos a
los tipo II y entre ellas; no se dividen.
Los neumocitos tipo II son redondeados, sobresalen y se encuentran en los
ángulos creados por tabiques alveolares; además poseen microvellosidades apicales
cortas y RER, REL y mitocondrias abundantes. Su núcleo es heterocromátimico.
22
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
En su citoplasma hay vesículas densas y cuerpos laminares densos, responsables
de la síntesis y secreción del surfactante. El surfactante se libera mediante exocitosis y
se dispersa por encima de una delgada capa de líquido que normalmente reviste la
superficie alveolar. Por este mecanismo, el surfactante pulmonar reduce la tensión
superficial en la superficie de contacto entre el aire y el líquido, reduciendo de este
modo la tendencia al colapso del alveolo al final de la espiración. Las células de Clara,
localizadas en los bronquiolos terminales, también secretan surfactante pulmonar.
Composición del surfactante:
x Colesterol – 50%
x Fosfolípidos – 40%
o Dipalmitoil fosfatidilcolina (DPpC)
o Fosfatidilcolina, fosfatidilglicerol, fosfatidilinositol, fosfatidilserina,
fosfatidiletanolamina, esfingomielina.
x Glucolípidos
x Proteínas (10%): SP-A, SP-B, SP-C, SP-D
Las proteínas específicas del surfactante (SP) corresponden a una glucoproteína
hidrófila (SP-A) y dos proteínas hidrófobas (SP-B y SP-C).
Se encargan del metabolismo y función del surfactante y sirven de defensa
inmunológica.
o El SP-A ayuda a formar la mielina tubular y media la respuesta
inmunitaria frente a virus, bacterias y hongos.
En menor medida promueve la fagocitosis y opsoniza y regula la
producción de citosinas.
o El SP-B forma una monocapa, adsorbe lípidos y extiende el surfactante
en la superficie alveolar. También interviene en la lisis.
o La SP-C estabiliza la monocapa.
o La SP-D participa en reacciones alérgicas y respuestas inmunológicas.
También estabiliza el fosfatidilinositol y es un opsonizador
En definitiva, las SP-B y SP-C interactúan fuera de la célula con SP-A y con
Ca2+ para formar una monocapa.
23
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
El agente tensoactivo reduce el riesgo de colapso en la espiración facilita la
reexpansión alveolar en la inspiración. Su ciclo de vida consiste en:
1. Inicialmente, se forma mielina tubular en la hipofase entre la película y la
superficie celular. Sirve de reserva de los componentes que se van a incorporar a
la monocapa según la necesidad (bidireccional). La monocapa está formada por
grandes agregados.
2. Con el tiempo, los componentes serán convertidos por proteasas y convertasas
en pequeños agregados, menos funcionales. Se produce reabsorción por
neumocitos II o fagocitosis por macrófagos.
En el pulmón enfermo actúan más enzimas como la elastasa con lo que hay
mayor porción de pequeños agregados.
24
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Los neumocitos tipo II son activos antes del nacimiento, sobre la semana
35. El desarrollo está controlado por el sistema endocrino (insulina, cortisol, prolactina,
tiroxina). El síndrome de dificultad respiratoria o enfermedad de las membranas hialinas
se produce por falta de producción de surfactante, por lo que el exudado es rico en
proteínas y fibrina, resultando mortal. Para combatirla se administra surfactante
exógeno en neonatos prematuros. A madres con amenaza de partos prematuros se les
administra cortisol.
Otra función de las células alveolares de tipo II es mantener y reparar el epitelio
alveolar cuando sufre alguna lesión. Cuando las células alveolares de tipo I se dañan, se
produce un aumento del número de las células de tipo II, que se diferencian a células
parecidas a las alveolares de tipo I.
Los neumocitos tipo II se dividen y se encargan de mantener y reparar el epitelio
alveolar en caso de lesión, por lo que su hiperplasia indica lesión alveolar. Las células
de Clara también proliferan y migran para reparar el epitelio alveolar dañado
(broncolización alveolar).
Intersticio
En el intersticio encontramos fibroblastos, leucocitos, mastocitos y fibras
colágenas y reticulares, que contribuyen a dar sostén. Las fibras elásticas permiten la
expansión de la pared alveolar (inspiración), la expulsión del aire por retracción
pulmonar (espiración) y evita el colapso bronquiolar y alveolar en la espiración. Entre
las células libres hay macrófagos alveolares, procedentes de monocitos, libres o
residentes en el intersticio, desde donde pueden migrar a la superficie pulmonar.
Los macrófagos alveolares ascienden hasta la tráquea, movimiento facilitado por
los cilios que arrastran el moco hasta la faringe. Se eliminan por la tos o la deglución
Igual que salen, pueden retornar al intersticio, aparecen en la pleura y paredes de los
bronquios, cargados de carbón. Son grandes, redondeadas y con alta tasa de recambio,
25
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
contienen CD64 y CD74. Su núcleo es irregular y abundan aparato de Golgi,
mitocondrias, ribosomas y glucógeno, con lisosomas muy heterogéneos.
Su función fundamental es de defensa frente a restos celulares (intersticio) y
polen, tóxicos, polvo y microorganismos (aire). En caso de reconocer algún Ag o toxina
segregan factores quimiotácticos que estimulan migración de leucocitos hasta epitelio
alveolar.
En los macrófagos puede haber lisosomas muy heterógeneeos, que pueden
corresponderse con distintas patologías:
x Pacientes cardiacos con congestión pulmonar, gránulos de hemosiderina de
eritrocitos fagocitados.
x El exceso de sílice provoca la liberación de agentes que producen alveolitis y
silicosis.
x Fumadores, muchas inclusiones y figuras de mielina.
x Fagocitan Mycobacterium tuberculosis pero no lo digieren. Un daño posterior
puede liberar las bacterias (tuberculosis recidivante)
Los macrófagos también contribuyen a la eliminación del surfactante degradado.
Los contaminantes atmosféricos oxidan FL del surfactante, degradándolo. Finalmente se
produce la eliminación del surfactante degradado.
En el intersticio no hay linfáticos, pero la vascularización es extensa, formando
una red capilar. En estos capilares tiene lugar:
x Metabolizar/transformar sustancias circulantes
x Transformación de angiotensina I en II (ECA)
x Destrucción de serotonina
x Inactivación de bradicina
x Intercambio de gases por difusión pasiva, donde la anhidrasa carbónica acelera
la expulsión de CO2.
o Se forma una barrera de difusión (aire-sangre) delgada, de 0,2 micras.
Está formada por el surfactante, el epitelio alveolar, el endotelio y las
láminas basales fusionadas. Aquí es donde tiene lugar la mayor parte del
intercambio de gases.
o También hay una barrera de difusión gruesa, con surfactante, epitelio
alveolar, lámina basal, intersticio con células y fibras, lámina basal
endotelial y endotelio. En estas zonas se puede acumular líquido.
El líquido acumulado drena por linfáticos de los bronquiolos terminales.
Por tanto, la pared alveolar tiene 2 porciones diferenciadas histológica y
funcionalmente. La acumulación de líquido no interfiere con el
intercambio gaseoso.
26
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Síndrome de dificultad respiratoria aguda (SDRA)
Si la barrera que impide la fuga de líquido desde los capilares se rompe se
producen dos mecanismos:
x Aumento de la pH en capilares alveolares. Suele deberse a insuficiencia del
ventrículo izquierdo, estenosis de la válvula mitral (edema cardiogénico,
insuficiencia cardiaca izquierda aguda).
x Lesiones en el epitelio alveolar o endotelio. Defectos en la permeabilidad, con
edema no cardiogénico:
o 1ª fase exudativa; edema intersticial y alveolar, infiltración, hemorragia y
depósitos de fibrina.
o 2ª fase, proliferativa: células alveolares proliferan y se diferencian para
regenerar el revestimiento epitelial alveolar, consiguiendo en la mayor
parte de los casos normalizar el intercambio de gases. En otros casos, en
o el intersticio se seguirán observando células inflamatorias y fibroblastos.
o 3ª fase, fibrosis crónica: oclusión de los vasos.
En la red de capilares también hay pérdida de agua, hasta 800 m al día. Una
ventilación inadecuada lleva a cambios en la presión parcial de CO 2, equilibrio ácido-
base y pH. Si hay hiperventilación la presión CO2 baja y el pH sube; si hay
hipoventilación ocurre lo contrario.
27
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
PLEURA
Superficie de intercambio de la caja torácica y superficie externa de los
pulmones están revestidas por serosas, pleura parietal y pleura visceral. Ambas se unen
en el hilio. Entre ambas hay líquido lubricantes que permite el deslizamiento al respirar.
Pleura parietal
Se asienta sobre:
x Tejido adiposo
x Tejido conjuntivo fibroso rico en vasos y nervios
x Periostio (costillas)
x Perimisio (músculos intercostales)
Pleura visceral
Mesotelio con microvellosidades apicales cortas. Descanse sobre un conjuntivo
laxo que contiene una lámina elástica externa. El conjuntivo inferior es rico en vasos y
nervios y tiene una lámina elástica interna que se continúa con las fibras elásticas de los
intersticios alveolares.
Sella la superficie pulmonar impidiendo la fuga de aire hasta la cavidad torácica.
Se puede producir neumotórax, hidrotórax, hemotórax, quilotórax o mesotelioma
(cáncer por amianto).
Ventilación
1. Inhalación
a. Requiere energía, con contracción del diafragma, los intercostales, escalenos y
accesorios.
b. Se amplía el volumen de la caja torácica.
c. Disminuye la presión interna
d. La diferencia de presión lleva al aire hasta los pulmones.
e. Los pulmones se amplían
f. Las pleuras se acercan
g. Se reduce el volumen y aumenta la presión
2. Espiración o exhalación
a. No requiere gasto energético, hay relajación pulmonar.
b. Disminuye el volumen de la cavidad pleural
c. Disminuye el volumen de la cavidad pleural
d. Aumenta la presión interna
e. Las fibras elásticas recuperan longitud inicial
f. Salida del aire
28
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Cuadros clínicos:
x Poliomielitis: la musculatura respiratoria se debilita y los músculos accesorios se
hipertrofian.
x Miastemia grave y síndrome de Guillaume-Barré: debilidad de músculos
respiratorios y accesorios, dificultad inspiratorio que lleva a la muerte.
VASCULARIZACIÓN
Linfática
x Profunda: No existe drenaje linfático ni en sacos alveolares ni en tabiques
interalveolares
Los linfáticos en bronquiolos respiratorios se unen y van creciendo
El drenaje tiene lugar en los ganglios peribronquiales.
x Superficial (pleura): En la pleura visceral y tabiques fibrosos que dividen el
parénquima pulmonar. Drena al espacio pleural.
Sanguínea
Puede ser a partir de arterias y venas pulmonares o bronquiales.
29
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Circulación pulmonar
Las arterias pulmonares se originan en el ventrículo derecho, traen la sangre
pobre el oxígeno y su presión es muy baja (25 mm Hg). Son arterias elásticas.
Desde la unión broncobronquiolar son musculares, disminuyen su tamaño y son
supernumerarias. La arteriola pulmonar da lugar a la red capilar en el intersticio de
tabiques alveolares, donde se oxigena.
Vuelve al corazón por vénulas, venas medias, venas mayores (sin elásticas
definidas y sin válvulas) hasta la aurícula izquierda.
Circulación mayor
Las arterias bronquiales son ramas laterales de la aorta torácica. Llevan sangre
rica en oxígeno, con presión sistémica (110-135 mmHg). Siguen el curso del árbol
bronquial hasta bronquiolos respiratorios, irrigando todo el tejido pulmonar excepto los
tabiques alveolares.
Se anastomosas con arterias pulmonares a la altura de la transición entre la
porción conductora y la respiratoria.
Cambios con la edad:
x Antes del nacimiento la vascularización pulmonar apenas está perfundida.
x En las primeras semanas se perfunde y aparecen arterias supernumerarias.
x En los adultos hay miocitos lisos en la íntima de las aterias bronquiales
x En la vejez la íntima de arterias y venas se engrosan.
30
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
APARATO CARDIOVASCULAR
INTRODUCCIÓN
El aparato cardiovascular es un sistema continuo y completamente cerrado de
tubos endoteliales. El objetivo general de este sistema es la perfusión de los lechos
capilares que permean todos los órganos con sangre fresca dentro de un estrecho rango
de presiones hidrostáticas. Se pueden diferenciar dos circulaciones:
x Menor o pulmonar: VD – pulmones – AI
x Mayor o sistémica: VI – resto del cuerpo – AD
ESTRUCTURA DE LOS VASOS SANGUÍNEOS
x Túnica íntima: endotelio y tejido conjuntivo subendotelial.
x Túnica media: músculo liso y tejido conjuntivo.
Músculo liso rodeado de un número variable de fibras de colágeno, matriz
extracelular y vainas elásticas con hendiduras irregulares (membranas elásticas
fenestradas).
x Túnica adventicia: tejido conjuntivo laxo.
La adventicia de los grandes vasos (arterias y venas) contiene pequeños vasos
(vasa vasorum) que penetran en la porción externa de la túnica media para
aportar oxígeno y nutrientes.
La túnica íntima está separada de la media mediante una capa de elastina
conocida como lámina elástica interna, mientras que la túnica media se separa de la
adventicia a través de la lámina elástica externa.
31
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
ARTERIAS
Elásticas
Tienen un diámetro mayor (>10 mm) y se encargan de la conducción. La aorta y
sus grandes ramas (las arterias braquiocefálicas, la carótida común, la subclavia y la
ilíaca común) son elásticas.
Las arterias elásticas de gran calibre tienen dos características fundamentales:
x Reciben sangre del corazón a una presión elevada.
x Mantienen la sangre circulando de forma continua aunque el corazón bombee de
forma intermitente. Como se distienden durante la sístole y se recuperan durante
la diástole, las arterias elásticas pueden mantener un flujo de sangre continuo a
pesar de que la acción de bombeo cardíaco sea intermitente.
Túnica íntima
x Endotelio:
o Células planas y alargadas paralelos al eje mayor del vaso.
o Zonulae occludens y nexos: aislamiento y comunicación.
o Vesículas de transcitosis (y/o receptores).
o Polaridad.
o Cuerpos de Weibel-Palade: Inclusiones bastoniformes que contienen
factor von Willebrand (VIII de coagulación).
Los anticuerpos contra el factor von Willebrand suelen usarse como
marcadores histoquímicos para la identificación de tumores derivados
del endotelio.
La activación endotelial consiste en cambios en las propiedades de las células
endoteliales en respuesta a diferentes estímulos, como bacterias, virus,
citotoxinas, productos lipídicos, componentes del complemento, hipoxia… Son
parte de la fisiología normal y se producen moléculas de adhesión y nuevos
productos (citosinas, moléculas vasoconstrictoras/vasodilatadoras…).
Funciones de las células endoteliades:
o Integridad estructural de la pared vascular: Gracias a sus células y a sus
conexiones.
o Barrera de permeabilidad selectiva, atendiendo a los siguientes criterios
Trasporte de moléculas según tamaño y carga
Vesículas de pinocitosis o transcitosis (muy abundantes): Entran
por la membrana apical y salen por la membrana basal.
Endocitosis mediada por receptor
o Homeostasis de la sangre
Cuerpos de Weibel-Palade: protrombógeno.
Anticoagulantes (trombomodulina, prostaciclina, activador de
plasminógeno).
32
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Mantenimiento de una barrera no trombógena entre las plaquetas
y el tejido subendotelial.
En caso de lesión el endotelio libera agentes protrombógenos
(factor von Willebrand, inhibidor del activador de
pllasminógeno) para estimular la agregación plaquetaria. Se
liberan factores que causan la formación de trombos.
Si la el tejido endotelial no estuviera bien diferenciado de las
plaquetas se formarían trombos peligrosos para el organismo.
o Regulación del flujo sanguíneo y la resistencia vascular: Se produce a
través de la contracción y la relajación de los miocitos lisos.
Vasoconstrictores: endotelina, E convertidora de angiotensina.
Vasodilatadores: NO y prostaciclina.
o Reacción inflamatoria y homing (linfocitos)
Homing: control de la interacción de linfocitos y de la superficie
endotelial.
Para llevar a cabo esta función las células endoteliales expresan distintas
moléculas de adhesión, así como diversas interleuquinas.
o Modificación de lipoproteínas por oxidación: Las células endoteliades
liberan radicales libres que oxidan LDL.
Importancia clínica: Los macrófagos fagocitan LDL modificados,
transformándose en células espumosas que dan lugar a placas
ateroscleróticas. La ateroesclerosis es una enfermedad inflamatoria
crónica caracterizada por el engrosamiento y endurecimiento de las
paredes arteriales. Las placas se forman en la túnica íntima de arterias
sometidas a una presión elevada, perdiendo sus propiedades.
1. LDL se infiltra y se acumula en tejido subendotelial.
2. Expresión de VCAM1, que permite el anclaje de monocitos que
atraviesan el endotelio y se diferencian en macrófagos, que
expresan RC SR-A que se une a LDL oxidadas.
3. La acumulación de macrófagos se transforman en células
espumosas cargadas de colesterol.
4. El núcleo de ateroma crece y atrae a los miocitos vasculares que
llegan de la media, formando una cubierta fibrosa.
5. Reacción inflamatoria: llegada de linfocitos T.
6. La secreción de mataloproteinasas y citosinas llevan a la rotura
de la placa, que predispone la trombosis.
o Síntesis de factores de crecimiento y hormonas
o Metabolización
Conversión de angiotensina I en II (pulmón).
Inactivación/conversión NA, trombina, 5-HT prostraglandinas,
bradicina.
o Angiogénesis
33
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Conjuntivo subendotelial: Se trata de tejido conjuntivo laxo con fibras elásticas
y colágenas y células miointimales, que son células musculares lisas con
capacidad contráctil encargadas de la síntesis de la MEC y también de las fibras
colágenas y elásticas. Se pueden observar también algunos fagocitos
(macrófagos).
x Lámina elástica interna: La lámina elástica interna no se distingue con claridad
debido a la gran cantidad de láminas elásticas existentes en la pared del vaso, es
la más interna de todas.
Túnica media
La túnica media de las arterias elásticas es la más gruesa, y consiste básicamente
en capas múltiples de células musculares lisas separadas por láminas elásticas:
x Láminas elásticas: hasta 50 concéntricas. La más externa es la lámina elástica
externa. Estas láminas elásticas se hallan fenestradas, de manera que se facilita
la difusión de sustancias dentro de la pared arterial. El número y espesor de las
34
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
láminas depende de factores como la tensión arterial y la edad, así, la aorta al
nacimiento no tiene casi ninguna lámina elástica, mientras que en el adulto hay
entre 40 y 70. En las personas hipertensas, aumenta tanto la cantidad como el
grosor de las láminas.
x Miocitos lisos: Se hallan unidos a las láminas, y se encargan de sintetizar la
MEC y las fibras colágenas y elásticas. Se trata de células muy activas, que
gracias a su capacidad sintética participan en procesos de reparación, y son
capaces de proliferar, migrando mayoritariamente hacia la túnica adventicia. Es
importante saber que en la túnica media NO hay fibroblastos.
x Fibras colágenas y sustancia amorfa (proteoglicanos): como ya se ha dicho,
sintetizadas por los miocitos lisos.
Correlación clínica de la túnica media:
Aneurisma
Cuando se produce una dilatación de un vaso permanente se forma un
aneurisma. Puede ser sifilítico o abdominal (consecuencia de la ateroesclerosis). En el
vaso afectado se produce:
x Pérdida de láminas elásticas y miocitos en la túnica media.
x Aumento del grosor de la pared por incremento del número de fibras de
colágeno, que es inelástico y no contráctil.
La pared por tanto es más gruesa pero resulta débil, no es resistente a la presión
elevada y tiende a romperse.
Síndrome de Marfan
El síndrome de Marfan es una enfermedad autosómica dominante causada por
mutaciones en el gen de la fibrilina 1, que provocan que esta proteína se vea alterada.
Ello puede provocar alteraciones esqueléticas, oculares (sobre todo en el cristalino) y
cardiacas. Suele asociarse a aneurismas disecantes de la aorta.
Túnica adventicia
x Red fibrilar laxa: Se trata de una red, no de láminas (está menos organizada que
la túnica media), de fibras colágenas y elásticas. Las fibras colágenas
contribuyen a impedir la distensión de la pared arterial más allá de los límites
fisiológicos durante la sístole.
x Fibroblastos y macrófagos.
x Vasa vasorum: Arterias y venas similares a las del sistema vascular general que
se encargan de proveer de nutrientes a la porción externa de la pared vascular y
eliminar productos de desecho, respectivamente. Sus ramas pueden penetrar
parcialmente en la túnica media, pero la túnica íntima se nutre por difusión
desde la luz.
x Nervi vascularis
35
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Musculares
Su número es mayor y están especializadas en la distribución. Tienen entre 10 y
0’1 mm de grosor. Por lo general, en la región de transición entre las arterias elásticas y
las musculares disminuye el componente elástico y aumenta el número de células
musculares lisas en la túnica media. Su diámetro disminuye al ramificarse, siendo su
pared relativamente gruesa.
Túnica íntima
Se diferencia de la de las arterias elásticas en que es más delgada y contiene una
lámina elástica interna prominente. Consiste en un revestimiento endotelial con lámina
basal, una capa de conjuntivo subendotelial y la ya mencionada lámina elástica interna
(aunque a veces el conjuntivo subendotelial es tan escaso que parece que están en
contacto la lámina basal endotelial y la lámina elástica interna).
El espesor de la túnica íntima aumenta con la edad, y en los adultos mayores esta
puede hallarse expandida por depósitos de lípidos, a menudo en forma de estrías
lipídicas irregulares.
Túnica media
Está formada hasta por 30 capas de miocitos lisos entre fibras colágenas y muy
escasas fibras elásticas (prácticamente consideramos desaparecido el componente
elástico).
Los miocitos son finos, cortos y están muy juntos, acoplados mediante nexos. Su
contracción contribuye a mantener la tensión arterial o tono: actúan como diana de
sustancias vasoactivas endoteliales (NO, prostaciclina, endotelina 1) y se hallan
36
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
inervados por el sistema autónomo (inervación simpática – vasoconstricción,
vasaparasimpática – vasodilatación).
Túnica adventicia
Es más gruesa que la de las arterias elásticas, del mismo espesor que la túnica
media, y suele estar separada de ella por una lámina elástica externa reconocible.
Contiene fibroblastos y fibras colágenas gruesas y elásticas. También en ella hallamos
vasa vasorum y nervi vascularis.
Arterias pequeñas
x Tienen entre 3-8 capas de miocitos y sin lámina elástica externa.
x No poseen tejido conjuntivo subendotelial.
SISTEMA MICROVASCULAR
Comprende las arteriolas, capilares y vénulas postcapilares. Las paredes de estos
vasos son parcialmente permeables, por lo que permiten el intercambio.
37
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Arteriolas
Tienen un diámetro de entre 20 y 120 micrómetros.
Se distinguen de las arterias pequeñas por la cantidad de miocitos lisos de la
túnica media, dado que las arterias pequeñas tienen entre 3 y 8 capas y las arteriolas
solo 1 ó 2. Los miocitos de las arteriolas tienen una marcada inervación simpática, lo
que les permite regular el flujo capilar por vasoconstricción. La contracción del músculo
liso de la arteriola aumenta la resistencia vascular y de este modo permite reducir o
cortar la entrada de sangre en los capilares. En el origen del lecho capilar que está en
contacto con la arteriola hay un engrosamiento en el músculo liso en relación con esta
finalidad, llamado esfínter precapilar.
La mayor parte de las arteriolas puede dilatarse entre el 60% y el 100% (vasos
de resistencia), y son capaces de mantener una contracción sostenida de hasta el 40%.
Distribuyen la sangre corporal, determinan la presión diastólica y controlan el tono
arterial y venoso.
Túnica íntima
Su endotelio es similar al de otras arterias, y se pueden apreciar prolongaciones
en las células endoteliales que llegan a los miocitos de la túnica media, con los que
establecen uniones en nexo. La lámina elástica interna solo existe en el extremo
proximal de la arteriola.
Túnica media - 1-2 capas de miocitos
Túnica adventicia - Mínima.
Metaarteriolas
Poseen sólo una capa discontinua de miocitos. No son vasos nutricionales, sino
que aseguran la continuidad del flujo. También presentan esfínteres precapilares.
ArteriolaÆ CapilaresÆ Vénula postcapilar
Metaarteriola
Capilares
Los capilares forman redes vasculares sanguíneas que permiten que líquidos con
gases, metabolitos y productos de desecho atraviesen sus paredes. En total, ocupan
80000 km de superficie. La cantidad de capilares depende del individuo, su peso, y su
actividad, y su construcción atendiendo a las distintas necesidades hace que la red
capilar tridimensional de un tejido, un órgano o un individuo sea específica, y no sea
por tanto intercambiable (por ejemplo, entre distintos órganos).
Su diámetro es de 5-10 micras, por lo que los eritrocitos y células sanguíneas
que discurren por ellos están en fila o plegados. Como consecuencia de este diámetro
38
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
tan pequeño, el flujo se ralentiza y se permite el intercambio bidireccional. La
permeabilidad de las sustancias que se intercambian es variable según las necesidades.
La presión alcanza 35 mmHg en el extremo arterial, siendo de 10 mmHg en el
extremo venoso.
En los capilares solo existe túnica íntima, ocupando el endotelio un 85% de la
superficie endotelial total. Es decir, los capilares solo disponen de endotelio y lámina
basal, cuyas características son similares a los de las arterias elásticas (ver en endotelio
arterias elásticas).
Distinguimos dos canales de flujo capilar:
x Preferencial: se trata de una vía por la cual la sangre fluye directamente de
arteriola a vénula postcapilar. Por ella la sangre siempre fluye. En ella participan
las metaarteriolas.
x Verdadero: los capilares se anastomosan formando un dominio capilar. El flujo
es intermitente, gracias en parte a los esfínteres precapilares.
Tipos de capilares
x Continuos: los encontramos en el tejido
conjuntivo, músculo, SNC, pulmón, piel, timo,
hueso. Su endotelio y su lámina basal son
continuos (el primero tiene un grosor de 0’2
micras). Posee:
o Acuaporinas y vesículas de pinocitosis y transcitosis (70 nm). En el SNC
no vamos a encontrar este tipo de vesículas.
o Pericitos o células de Rouget: alargados, estrellados y contráctiles
(regulan perfusión). Actúan como fagocitos y poseen nexos. Se hallan
entre el endotelio y la lámina basal.
x Fenestrados: se hallan en las glándulas endocrinas y en los sitios de absorción de
líquidos y metabolitos, como la vesícula biliar, los riñones y el tubo digestivo.
Su membrana basal también es continua, pero su endotelio (de 0’1 micras) posee
poros de unos 70 (80-100 según Ross) nm, que en ocasiones tienen un fino
diafragma no membranoso compuesto de un material fibrilar radial. La carga de
este diafragma, al igual que la de la membrana basal endotelial, es negativa.
Hallamos escasas vesículas de transcitosis en estos capilares.
39
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Discontinuos: tienen un diámetro mucho
más grande que el capilar convencional, de
unas 30 o 40 micras. Su recorrido es
irregular y sinuoso, puesto que se adaptan a
los espacios intercelulares, de ahí que
también se les conozca con el nombre de
sinusoides. Son típicos del hígado, el bazo y
la médula ósea. Su lámina basal no es
continua y su endotelio es incompleto (las células endoteliales no están unidas
por occludens y no hayamos diafragma cubriendo los poros).
40
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Mecanismo de transporte
x Difusión: CO2 y O2.
x Transcitosis
x Transcitosis mediada por receptor
x Filtración y reabsorción: a ellas se debe la formación del líquido intersticial.
o La P hidrostática en los capilares obliga al agua y solutos pequeños a
salir de ellos. Esta P es máxima en el extremo arterial.
o La P coloidosmótica atrae a líquido y solutos al interior del capilar.
Es constante.
Equilibrio de Starling: Mto=K(Pc+πi)-(Pi+πc)
La hipótesis de Starling nos dice que el movimiento de las moléculas a través de
la pared capilar depende de una constante de permeabilidad, específica para cada
capilar. La hidrostática en el capilar y la oncótica en el intersticial, que tienden a la
filtración, se ponen como positivas, y la hidrostática del intersticial y la oncótica del
líquido son negativas, pues se produce absorción. Luego si nos da positivo habrá
filtración y si no absorción.
Mto=K(Pc-πc)
Podemos pasar de todo lo demás porque es muy pequeño, a no ser que haya
patología. Cuando la hidrostática en el capilar sea superior a la oncótica, habrá
filtración, cuando sea menor tendremos absorción.
Se ha de tener en cuenta que un 10% del líquido filtrado no se reabsorbe, sino
que es devuelto mediante vías linfáticas.
Vénulas postcapilares
Su luz es mayor que la de los capilares (10-25 micras). Una vénula postcapilar es
similar al capilar continuo, pero con más pericitos. Al igual que los capilares, está
dotada solo de endotelio (cuyas células están unidas por occludens laxas) y membrana
basal.
Estas vénulas son responsables de drenar la sangre capilar. Su endotelio es el sitio
diana principal de agentes vasoactivos como la histamina o la serotonina: la respuesta a
estos agentes produce la extravasación de líquido y la migración de los leucocitos desde
el vaso durante la inflamación y las reacciones alérgicas (diapédesis). De ahí que en la
pared de las vénulas postcapilares haya numerosos receptores para los linfocitos.
Las vénulas postcapilares de los órganos linfoides secundarios se conocen
también como vénulas del endotelio alto (VEA o HEV).
41
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
SISTEMA VENOSO
Vénulas colectoras
Drenan en las vénulas musculares.
Vénulas musculares
Tienen un diámetro de 50-100 micras. Su luz está colapsada y su contorno es
irregular. Su túnica media es más gruesa que la de las vénulas anteriores (que no
poseían) y está constituida por 1 ó 2 capas de miocitos. Su delgada adventicia posee
fibras colágenas. No tiene láminas elásticas ni interna ni externa, ni pericitos.
Venas
El sistema venoso como tal es
un sistema de diámetro ascendente y
presión descendente que lleva la
sangre al corazón. En comparación
con las arterias, las venas poseen una
luz más grande y una pared más
delgada y distensible. Tienen una gran
capacitancia, almacenando hasta un
70% del volumen sanguíneo.
El flujo venoso depende de
fuerzas externas (musculatura
esquelética circundante) y de válvulas.
Las venas tienen 3 túnicas con límites poco definidos:
Túnica íntima
No tiene lámina elástica interna.
En ella hallamos válvulas. Las válvulas son
proyecciones pares revestidas de endotelio y
reforzadas por fibras elásticas y colágenas. Su borde
libre se orienta al corazón y en su zona de inserción
la pared venosa es más fina, y sus miocitos son
longitudinales.
La contracción de la musculatura esquelética
asegura el flujo unidireccional de la sangre, que
avanza produciendo la distensión en zonas de
ensanchamiento llamadas senos. Para evitar el reflujo
desde los senos venosos, las válvulas se cierran.
42
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Correlación clínica: venas varicosas
Las venas varicosas son más comunes en las piernas, aunque también hay
varices esofágicas, hemorroides y varicoceles (cordón espermático), de modo que se
pueden producir en otras partes del organismo (aun así, la más frecuente y susceptible
es la vena safena). Se producen ante un mal funcionamiento de las válvulas que las
obliga a trabajar en exceso, hipertrofiándolas y volviéndolas más tortuosas a la par que
haciendo más sensible la pared vascular. Se pierde el tono muscular, lo cual influye en
el retorno venoso también.
Túnica media
Más delgada que en las arterias. Tiene fibras elásticas y colágenas y miocitos de
distribución irregular, aunque normalmente concéntrica. A diferencia de las arterias, en
la túnica media existen menos haces de células musculares lisas asociadas a fibras
reticulares y elásticas.
Tipos de venas
x Pequeñas
x Medianas
o Íntima con endotelio y conjuntivo subendotelial
o Media con 3-4 capas de miocitos lisos concéntricos
o Adventicia grande de conjuntivo laxo
o Válvulas
x Grandes
o Íntima con endotelio, conjuntivo subendotelias y algunos miocitos.
En personas mayores la íntima está engrosada con miocitos
longitudinales y aparece lámina elástica externa.
o Media muy delgada, con pocas capas de miocitos
o Adventicia muy gruesa con conjuntivo y capas de miocitos lisos
longitudinales (adventicia muscular).
o Vasa vasorum.
43
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
VASOS PORTA
En el riñón, el hígado y la hipófisis hay vasos interpuestos entre dos dominios
capilares. Estos vasos pueden contener tanto sangre arterial como venosa.
Riñón: arteriola eferente
Transporta sangre arterial desde capilares del ovillo glomerular (situado en el
glomérulo renal) hasta capilares peritubulares (vasos rectos). En los capilares
glomerulares no hay intercambio de gases ni de nutrientes, sino que se trata de un
ultrafiltrado de plasma.
Hígado: vena porta
Transporta sangre venosa rica en nutrientes desde redes capilares del digestivo,
páncreas, vesícula biliar y bazo hasta las sinusoides del hígado. La sangre drena por los
vasos colectores.
Sistema porta hipofisiario
Las vénulas conectan el plexo primario (eminencia media) con el plexo
secundario (adenohipófisis). Se trata de sangre venosa, rica en factores estimuladores e
inhibidores segregados por el hipotálamo.
44
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
ANASTOMOSIS ARTERIOVENOSAS
Son comunicaciones directas entre arteriolas y vénulas. Se evita el dominio
capilar (flujo no nutricional). Se encuentra en lechos unguenales de manos y pies,
pabellones auriculares, nariz, labios, pene y clítoris.
Tipos de anastomosis arteriovenosas:
x Rectas o en puente
x Retorcidas o en glomo
Están fuertemente encapsuladas y contienen una gran pared muscular. El
extremo arterial es muy grueso y se denomina canal de Sucquet, y está rodeada de
miocitos. Después viene una región central con miocitos longitudinales entre el
endotelio y la media., sin lámina elástica interna. Finalmente termina en el extremo
venoso, más delgado.
El extremo arterial está muy inervado por el sistema simpático y el
parasimpático. El simpático se encarga de cerrar la anastomosis, de modo que la sangre
pasa a los capilares. Cuando se produce la relajación muscular la sangre es capaz de
pasar a la anastomosis y evitar el capilar.
Regulan la perfusión periférica junto con los esfínteres precapilares.
Un ejemplo de ello es la piel, donde contribuyen a la termorregulación en la
superficie corporal. Cuando la anastomosis se cierra el flujo de sangre entra en los
capilares dérmicos, produciendo la pérdida de calor. Pero cuando está abierta, el lecho
capilar se cierra y el calor se conserva.
En el pene y el clítoris participan en el inicio de la erección. Al cerrarse la
anastomosis, el flujo sanguíneo se dirige a los cuerpos cavernosos.
SISTEMA DE LAS VÍAS LINFÁTICAS
Funciones
El sistema de las vías linfáticas tiene tres funciones principales:
1. Es un sistema de drenaje: retira el exceso de líquido tisular, que se está
generando de forma constante debido a que el sistema microvascular se produce
una reabsorción del 90% del agua y los solutos previamente filtrados, pero un
10% permanece como líquido tisular en los tejidos. Este exceso de líquido es
drenado por las vías linfáticas, que se encargan de transportarlo al sistema
venoso. Pero este sistema no es independiente, completo, no forma una
circulación, simplemente devuelven a la sangre lo que en un momento dado se
había filtrado: agua, solutos y proteínas. Estas últimas están en una cantidad
45
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
apreciable, dependiendo de la proteína que midamos podemos estar hablando de
en torno a un 50% de la proteína circulante, que se devuelve a la sangre cada día.
2. Participa en el sistema inmunológico, puesto que conduce la linfa y las células
inmunitarias a los ganglios linfáticos
3. Interviene en el transporte de los quilomicrones
Este sistema de las vías linfáticas es un sistema muy generalizado en el
organismo, pero no es un sistema universal. Determinados tejidos, como el epitelial,
cartilaginoso y óseo, placenta, SNC, médula ósea y oído interno no lo presentan.
Flujo linfático
La linfa es un flujo unidireccional que circula a muy baja presión y no presenta
una bomba central (como el corazón). Entonces, ¿Cómo es posible que la linfa fluya?
Por dos mecanismos:
1. Contracción intrínseca: reside en la capacidad del vaso de expandirse y
contraerse. La musculatura lisa del vaso permite que este se contraiga, y cuando
esto ocurre la linfa se empuja hacia delante. No todo el vaso se expande y se
contrae a la vez, esto ocurre de trecho en trecho: cada segmento entre dos
válvulas consecutivas se llama linfangión, y estos linfangiones actúan como
mini-bombas. Un linfangión se expande, tras ello se contrae e impulsa la linfa
hacia delante, con lo que la válvula que está delante se abre y posteriormente se
cierra. Por tanto, la linfa fluye lentamente, segmento a segmento, hacia delante.
2. Contracción extrínseca: la musculatura esquelética circundante comprime los
linfáticos cuando ejercitamos y hace con ello que la linfa fluya hacia delante.
Se calcula que cada día se devuelven a la sangre entre 2 y 3 L de linfa. Podemos
definir linfa como un ultrafiltrado muy claro de plasma sanguíneo, con un contenido
bajo en proteínas (2-5%) y alto en linfocitos. Hay una excepción: la linfa intestinal,
particularmente tras la ingesta, es muy blanquecina, de aspecto lechoso y altamente
enriquecida en quilomicrones (colesterol y ácidos grasos). Esta linfa intestinal se
absorbe en unos vasos que se denominan quilíferos.
Clínica
x Si el drenaje linfático no funciona bien, el líquido tisular se acumula en los
tejidos, dando lugar a lo que conocemos como edema.
x En quemaduras se produce la salida de plasma (se rompen los vasos sanguíneos)
y proteíans plasmáticas. El aumento de la presión oncótica en el tejido debido a
las proteínas que han salido lleva a una mayor pérdida de plasma fuera del lecho
capilar.
46
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Constitución histológica general
Los vasos linfáticos recuerdan a las venas, pero su luz es mayor. Hay una
tendencia a hablar de tres capas o túnicas en ellos, aunque su diferenciación resulta más
compleja que la de las túnicas de arterias y venas:
1. Íntima: constituida por endotelio y conjuntivo subendotelial. Forma las válvulas,
muy numerosas.
2. Media: con unas cuantas capas de miocitos dispuestos circularmente y fibras
colágenas.
3. Adventicia: en ella hallamos conjuntiva rica en fibras colágenas y elásticas.
Capilares linfáticos
Se trata de vasos sumamente finos, de hasta 100 micras de diámetro. Sólo tienen
túnica íntima. Sus extremos son ciegos, son pequeños tubos cerrados por un extremo
que son capaces de formar plexos, de ramificarse. Es frecuente encontrarlos en el
conjuntivo laxo y en las mucosas, así como en proximidad a los capilares sanguíneos.
Los plexos capilares linfáticos tienen alta permeabilidad,
por lo que van a ser vías preferentes para la entrada de grandes
moléculas (lípidos y proteínas grandes). Ello se debe a que en
ellos el endotelio es más fino y hay menos complejos de unión
célula-célula, y además la lámina basal es discontinua.
Es importante señalar la presencia de filamentos de
anclaje entre la lámina basal y el conjuntivo perivascular, que
son clave cuando la presión se eleva en situaciones de
inflamación, ya que evitan el posible colapso, asegurando que el
líquido tisular siga entrando al capilar linfático. Ya hemos dicho
que los capilares linfáticos se unen en plexos, y van a acabar
formando vasos de pared más gruesa denominados vasos
colectores.
Vasos colectores
Su pared es algo más gruesa, similar a la de una vena pequeña. Suelen rodear a
las venas y también pueden anastomosarse. Presentan un aspecto en collar de perlas
porque la linfa está separada en linfangiones, delimitados por las válvulas. En ellos,
además de la túnica íntima, encontramos una túnica media formada por 1 ó 2 capas de
miocitos lisos capaces de contraerse, iniciando ondas peristálticas que contribuyen al
avance lento y paulatino de la linfa.
Los vasos colectores se unen formando vasos linfáticos terminales, que se
pueden observar en las proximidades de un ganglio linfático, si bien antes de llegar el
ganglio se dividen dando lugar a los vasos linfáticos aferentes, que perforan la cápsula
del ganglio e introducen en él la linfa.
47
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Senos linfáticos
En el interior de los ganglios, la linfa se encuentra en unos espacios
denominados senos linfáticos. Como los senos también están delimitados por células
endoteliales, se los puede definir como vías linfáticas intraganglionares. En ellos la linfa
circula despacio, y los Ag pueden activar los linfocitos de los ganglios desencadenando
una respuesta inmunológica. La linfa sale del ganglio por los linfáticos eferentes.
Clínica
Todos los capilares linfáticos de un área drenan en los ganglios linfáticos
regionales. Ello es muy importante en la diseminación del cáncer, ya que las células
cancerosas acceden a las vías linfáticas. Si se quedan en un ganglio son capaces de
replicarse y producir tumores secundarios. Por ello los ganglios regionales se han de
extirpar, y así se evita la metástasis y la diseminación.
Vías linfáticas de mayor calibre
Finalmente toda esta linfa puede volver al sistema venoso por dos vías que
drenan en la encrucijada yugulo(int.)-sublavia:
1. Conducto torácico: en el lado izquierdo. Tiene unos 5 mm de diámetro, y
presenta una íntima muy desarrollada con una lámina elástica interna marcada,
una media gruesa con miocitos tanto concéntricos como longitudinales y una
adventicia mal delimitada que también tiene miocitos.
2. Conducto linfático principal derecho: en el lado derecho.
En ambos casos vamos a encontrar una válvula que impide el reflujo, evitando
que la sangre se introduzca en el vaso linfático.
Clínica
x Elefantiasis: infección parasitaria de los vasos linfáticos (por Wucheria bancrofti
o por Brugia Malayi, como curiosidad). Causa linfedema crónico en los
miembros inferiores y en genitales.
x Ascitis quilosa y quilotórax: se deben a la acumulación de líquido rico en grasas
en el abdomen o en el tórax por traumatismo que destruya las vías linfáticas, por
su obstrucción o por un desarrollo anómalo de las mismas.
48
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
EL CORAZÓN
El corazón es un órgano compuesto por cuatro cavidades: dos atrios (derecho e
izquierdo) y dos ventrículos (derecho e izquierdo), siendo el responsable del flujo de la
sangre mediante contracciones rítmicas y se le conoce como una bomba muscular de
presión aspiración.
Es el principal factor determinante de la presión arterial sistémica. Y como se ha
mencionado anteriormente tiene 4 cámaras, tabiques interauriculares e interventriculares
y válvulas que impiden el reflujo. Pesa 300 gramos y sus dimensiones son 12x9x6 cm.
La sangre proveniente de todo el organismo retorna al atrio derecho desde donde
pasa al ventrículo derecho. Desde el ventrículo derecho la sangre se bombea hacia los
pulmones para que se oxigene y luego retorna al atrio izquierdo. El atrio izquierdo envía
la sangre al ventrículo izquierdo desde donde se bombea hacia el resto del organismo o
sea al circuito sistémico.
El corazón, que se diferencia a partir de un tubo vascular recto en el embrión,
tiene la misma estructura básica de tres capas en su pared que los vasos sanguíneos de
orden superior a los capilares y vénulas poscapilares. En los vasos sanguíneos las tres
capas se denominan túnica íntima, media y adventicia, mientras que en el corazón se
llaman endocardio, miocardio y epicardio, respectivamente. Además tiene un sistema
cardionector, esqueleto fibroso…
El corazón tiene dos tabiques:
1. El tabique interauricular, el cual es más fino y delgado y en el que se distingue
un eje de miocitos cardiacos y un revestimiento endotelial en ambas caras.
2. El tabique interventricular que es más grueso y además de tener el eje de
miocitos, el revestimiento endotelial, tiene también un elemento que forma parte
del elemento fibroso: la pars membranosa que no tiene eje muscular.
Endocardio
Está en contacto con la sangre y en continuidad con la íntima. Es de membrana
lisa y brillante. Hay que recalcar que NO es íntima, sino que está en su continuidad.
El auricular es más grueso y blanquecino, mientras que el ventricular es más
translúcido. Las lesiones de eyección son áreas localizadas de endotelio engrosado y son
consecuencia de un flujo turbulento. En este tipo de patologías el endotelio engrosado
se puede comparar con la forma de los parches. Suele aparecer en las aurículas.
El endotelio a su vez se divide en tres capas: interna, media y externa.
x La capa interna es como la íntima y corresponde al endotelio y al conjuntivo
subendotelial.
x La capa media va a ser a más gruesa, siendo una zona de tejido conjuntivo, con
fibras elásticas organizadas en haces paralelos y también miocitos.
x La capa externa se encuentra bajo el miocardio, se denomina capa
subendocárdica y es la más externa del endocardio. Hay tejido conjuntivo, vasos
y fibras nerviosas. En esta capa es donde se encuentra el sistema cardionector.
Hay algunos puntos en los que no se encuentra, como en los músculos papilares
49
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
y las cuerdas tendinosas. Esta capa, como se ha explicado, está pegada a
continuación del miocardio y ofrece fijación del endocardio al miocardio. El
endocardio sufre su estrés y el de la capa con la que se pega, es para eso la
fijación, para evitar que se produzca una rotura.
Miocardio
Corresponde al tejido muscular esquelético cardiaco. En él hay miocitos
cardiacos, junto con un tejido conjuntivo (endomisio). En las aurículas son menores los
miocitos y tienen un mayor desarrollo del RER y del AG, con rica red fibrilar elástica.
En el ventrículo, es todo al contrario, con grandes miocitos y pobre cantidad de
orgánulos, aunque está más desarrollado
El miocardio está más desarrollado en los ventrículos, con un desarrollo máximo
en el lado izquierdo. La capa más externa es más lisa, pero el miocardio que está en
contacto con la subendocárdica es tremendamente irregular y a esa zona se le denomina
trabéculas carnosas.
Hay dos sincitios, uno para las aurículas y otro para los ventrículos. Los sincitios
son unidades histológicas y funcionales y están separados por elementos conjuntivos
principalmente fibrosos. Son responsables de la contracción del corazón las cuales
consumen muchísima energía, es por ello por lo que el corazón tiene que estar muy
vascularizado.
Si una persona tiene una coronariopatía, tiene una alta probabilidad de sufrir
isquemia, si no se trata puede llevar a tener una angina y al final puede tener hasta un
infarto de miocardio. Este tipo de sucesos ocurre fundamentalmente en la zona antes
mencionada, siendo lo normal en el ventrículo izquierdo.
Además de miocitos, hay células mioendocrinas que aparecen fundamentalmente
en las aurículas y sobre todo en la aurícula derecha. Son capaces de segregar hormonas
a la sangre, en concreto sintetizan un péptido precursor del Péptido Natriurético
Auricular.
En las células mioendocrinas se localizan cerca del núcleo los orgánulos, siendo
su localización variable, pues tienen una variación circadiana dependiendo de nuestras
ingestas.
El PNA, mencionada anteriormente, tiene un efecto con el aparato urinario.
Aumenta la cantidad de excreción de agua y sales, evitando así la hipervolemia.
Además también afecta al riñón inhibiendo la secreción de renina y aldosterona,
consiguiendo con ello reducir la presión arterial. Además tiene un efecto directo sobre
el propio sistema cardiovascular, pues lo relaja, reduciendo la presión con las
secreciones y relajando las capas musculares.
Se debe recalcar la presencia de músculos papilares, que son montículos con
proyección hacia la luz, y que están en el miocardio de los ventrículos. Son el lugar
donde se insertan las cuerdas tendinosas.
50
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Epicardio
Es la capa más externa y está asociado o forma parte de una estructura más
grande que es el pericardio. Este último lo forman dos capas: la visceral y la parietal.
Además tiene una cavidad pericárdica llena de lubricante. Si durante la auscultación hay
anomalías, si se oye un ruido de roce, hay daño en el pericardio.
Es un mesotelio (procede del mesénquima, epitelio plano simple) liso sobre
conjuntivo laxo rico en fibras elásticas. Tiene tejido adiposo blanco asociado y una gran
vascularización e inervación. Contiene grandes arterias y venas coronarias superficiales,
lo cual es de gran interés quirúrgico para los injertos.
ESQUELETO FIBROSO
Formado por distintos componentes. La misión principal es proveer puntos de
fijación, separando claramente los dos sincitios y actuando como aislante eléctrico al
impedir el libre flujo de impulsos eléctricos entre Aurícula y Ventrículo. Tiene tabiques
intermembranosos.
1. La pars membranosa del tabique interventricular está en la parte superior del
tabique, por debajo de la válvula aórtica. Carece de músculo cardíaco y está
formado por conjuntivo denso, con un segmento corto de haz de His.
2. El cuerpo central fibroso valvular se encuentra en el repliegue del endocardio,
siendo un elemento fibroso en la zona central. Las válvulas están formadas por
tres capas: esponjosa, fibrosa y ventricular. La primera y la tercera revestidas por
endotelio:
1. La zona esponjosa o da la aurícula lo da a una arteria. Además de tener el
endotelio, hay un conjuntivo laxo que tiene algunas fibrillas elásticas y una
matriz muy rica en proteoglucanos, importante para el funcionamiento
valvular, pues les da plasticidad a las válvulas. El conjuntivo y los
proteoglucanos amortiguan el golpe que cada uno de los cierres supone. Da a
la aurícula o a las arterias revestimiento endotelial.
2. La capa ventricular da al lado opuesto, hay endotelio y un conjuntivo muy
denso con muchas fibras elásticas en las válvulas auriculoventriculares se
continúa con las cuerdas tendinosas.
3. La zona fibrosa formada por tejido conjuntivo denso y está en continuidad
con los anillos fibrosos, estando en la base de la válvula los anillos.
*Recordar que las válvulas lo normal es que sean avasculares, y solo hay vasos en la
base de la válvula. Las valvulopatías cardíacas son muy numerosas. La causa suele ser
la angiogénesis valvular: la estructura normal se altera, las fibras fibrosas por colágenas
y la válvula se hace más rígida. Además el flujo será malo con una insuficiencia
cardiaca.
3. Los anillos fibrosos son elementos conjuntivos fibrosos, densos, que se
encuentran alrededor de los orificios de la aurícula y el ventrículo. El conjuntivo
denso está en continuidad con el eje fibroso de las válvulas. Provee puntos de
inserción para las válvulas.
51
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
4. Los trígonos fibrosos están entre los anillos fibrosos, y entre las válvulas se
encuentran unas zonas con forma de cuña conformadas por tejido conjuntivo
denso, en él puede haber una diferenciación a tejido cartilaginosos fibroso,
puede haber condrocitos.
5. Las cuerdas tendinosas son una especie de filamentos con la cara ventricular de
las válvulas atrioventriculares desde la punta de la válvula hasta la punta del
músculo papilar. Los elementos están formados por fibras de colágeno y
endocardio. Pues evitan la eversión de las hojas durante la contracción
ventricular, haciéndolas de sentido único.
SISTEMA CARDIONECTOR
El miocardio es capaz de contraerse de forma rítmica por sí mismo. Tiene
contracción involuntaria, la frecuencia cardiaca es modulada por el sistema nervioso
autónomo. La inervación simpática va a aumentar la frecuencia, mientras que la
parasimpática la va a disminuir.
Todos los miocitos cardiacos son excitables. Todos son capaces de generar,
conducir y transmitir impulsos, pero hay unos miocitos cardiacos especializados que
van a ser capaces de conducir los impulsos a una mayor velocidad y que van a iniciar el
impulso excitatorio.
El sistema cardionector genera impulsos con una frecuencia más alta, a una
velocidad 4 veces mayor que los miocitos comunes. Sus impulsos son capaces de
atravesar el anillo fibroso.
Es el encargado de coordinar la contracción de aurículas y ventrículos. Está
formado por dos nódulos sinusal y auriculoventricular, el haz auriculoventricular de
His, ramas izquierda y derecha o de Tawara y ramificaciones terminales.
Nódulo sinusal
Está en la capa subendocárdica, en la desembocadura de la vena cava superior en
la aurícula derecha. Genera impulsos para provocar las contracciones rítmicas del
músculo cardíaco.
Consiste en una malla irregular de cardiomiocitos junto con tejido conjuntivo.
Los miocitos son más cortos y delgados.
Aquí se generan los impulsos con mayor frecuencia de concentración (60-100).
Ademá, actúa como marcapasos cardiaco y produce la contracción de la aurícula
derecha.
Desde el nódulo sinusal en impulso va a una vía internodal (músculo auricular
intermodal). Tiene 3 haces: anterior, medio y posterior, indistinguibles. El impulso sufre
un retraso en esta vía intermodal y llega al nódulo interventricular.
Nódulo auriculoventricular
Está en la capa subendocárdica, en la pared medial de la aurícula derecha, justo
encima del anillo valcular tricuspídeo. y en la base del tabique interauricular.
52
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Hay una malla irregular de miocitos especializados con conjuntivo denso muy
vascularizado. Su organización es menos al azar, aunque los miocitos son muy similares
a los de nódulo sinusal.
Haz de His, ramas de Tawara y ramificaciones terminales
El haz de His es la principal vía de conducción desde las aurículas hasta los
ventrículos. Atraviesa el esqueleto fibroso del tabique interventricular y se divide en dos
ramas (ramas de Tawara), de las cuales la derecha es más fina, que transmiten el
impulso a los ventrículos. Una vez han alcanzado los ventrículos, los impulsos
nerviosos se extienden a través de las ramificaciones terminales.
Los miocitos de estas zonas están especializados (fibras de Purkinje), son
mayores y más gruesos, pero igualmente estriados. Los núcleos son redondeados y están
en grupos. Tiene escasas miofibrillas periféricas, pero poseen abundante glucógeno.
Los miocitos están alineados y en contacto por los extremos y los lados. Tienen
discos intercalados atípicos y grandes nexos que ponen las superficies en contacto. Se
encuentran separadas de los miocitos cardiacos comunes por conjuntivo, aunque en las
fibras terminales tienen nexos con los miocitos comunes.
La velocidad de transmisión del impulso puede alcanzar 2-3 m/s, Permite la
contracción de los miocitos del ventrículos. La excitación se transmite de primero hasta
los músculos papilares y, tras ello, al miocardio ventricular.
IRRIGACIÓN CORONARIA
Es muy importante, especialmente en el ventrículo izquierdo. Hay dos arterias
coronarias son ramas que nacen de la aorta: la arteria coronaria derecha y la arteria
coronaria izquierda, y son arterias elásticas. Circulan por el epicardio y emiten
abundantes ramas. Las anastomosis de arterias en el miocardio no permiten mantener la
circulación colateral efectiva, por lo que se consideran arterias terminales. Si hay
oclusión coronaria se produce un infarto.
Las venas drenan en la vena coronaria mayor y de ahí la sangre pasa al seno
coronario, que drena a la aurícula derecha. Las venas menores desembocan
directamente a la aurícula derecha.
FORMACIÓN DEL SISTEMA VASCLAR
Vasculogénesis (embrión)
Se da en la 2ª semana de vida, empieza en el sáculo vitelino y en el pedículo de
fijación. Los angioblastos son los progenitores endoteliales vasculares libres y
migratorios. Empiezan a agruparse en columnas delgadas, se anastomosas, forman una
luz y los vasos delgados se van uniendo entre sí. Se consigue una organización tubular.
Está estimulada por el VEGF (factor de crecimiento endotelial vascular),
sintetizado por células mesenquimales. Hay receptores en células endoteliales (VGEF-
53
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
1) y en angioblastos (VEGF-2): Forma una red vascular primitiva que asegura la
supervivencia del embrión.
Angiogénesis (embrión y adulto)
Formación de un vaso a partir de uno prexistente (capilar). Las células
endoteliales mantienen la capacidad proliferativa. Tiene lugar en el crecimiento de
órganos. Otros ejemplos son el ovario y el útero en el ciclo menstrual o el crecimiento
placentario durante la gestación.
54
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
La Ang1 media la maduración vascular. En ausencia de VEGF, la Ang2 bloquea
los efectos de la Ang1, lo que se traduce en la remodelación o la apoptosis endotelial.
Este proceso está asociado a:
x Respuestas inflamatorias, cicatrización de heridas, procesos reparadores.
x Degeneración macular asociada al envejecimiento.
x Angiogénesis tumoral: Células cancerosas producen péptidos angiogénicos que
estimulan la proliferación de células endoteliales para fomración de nuevos
vasos. Este mecanismo es imprescindible para la supervivencia tumoral. La
angiostatina y la endostatina son péptidos antiangiogénicos que retrasan o
detienen la angiogénesis.
x Sarcoma de Kaposi: Tumor con nódulos vasculares en piel, la mucosa y los
órganos internos y que con frecuencia afecta a pacientes con SIDA. Los nódulos
vasculares están constituidos por células neoplásicas fusiformes y espacios
vasculares muy desarrollados. Las células fusiformes expresan marcadores de
células endoteliales vasculares y linfáticas, lo que sugiere su origen endotelial.
INERVACIÓN VASCULAR Y CARDÍACA
Inervación vascular
La inervación vascular eferente hace referencia a las fibras simpáticas y
parasimpáticas que inervan los miocitos de la mayoría de las paredes vasculares.
En la mayor parte de los vasos se encuentran exclusivamente fibras simpáticas,
que inervan a los miocitos vasculares; y sólo en algunos casos es posible localizar
también parasimpáticas, causando dilatación. Gracias a las fibras simpáticas se produce
vasoconstricción.
En cuanto a la inervación aferente se puede decir que sólo se puede localizar en
algunas zonas del cuerpo, como en el seno carotídeo, en la región del cayado de la aorta,
en la arteria pulmonar o en las grandes venas que llegan al corazón, es posible encontrar
estructuras que funcionan como quimiorreceptores y barorreceptores.
Las dos más estudiadas son:
x El glomo o cuerpo carotídeo, el cual se encuentra en la bifurcación de la arteria
carótida común.
x El glomo aórtico o paraganglios, que está en el cayado aórtico.
Estos quimio o barorreceptores son de estructura alargada, presentan una cápsula
conjuntiva desde la cual se proyectan finas trabéculas hacia el interior. Se encuentran en
las paredes de esos vasos y ellos mismos tienen gran vascularización, presentando
capilares fenestrados.
Aparte de capilares fenestrados podemos encontrar fibras nerviosas aferentes
terminales que en su mayoría proceden del glosofaríngeo. Al final estas fibras nerviosas
acaban haciendo sinapsis con las células glómicas. Hay dos tipos celulares: células tipo
I o glómicas y células tipo II o de sostén.
55
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Las células glómicas son células derivadas de la cresta neural, por lo tanto
cromafines, pudiéndolas visualizar si utilizamos tinciones metálicas. Tienden a
agruparse y podemos observar nexos, en concreto uniones tipo GAP entre ellas. Son
pálidas, con un núcleo muy pálido eucromático. Pero lo que más destaca son los
gránulos electrodensos en el citoplasma. Estos gránulos contienen una gran variedad de
neurotransmisores.
Hasta hace poco se pensaba que estas células glómicas eran auténticos quimio y
barorreceptores, pero parece ser que estas lo que hacen es modular las fibras nerviosas y
por lo tanto serían estas fibras nerviosas los verdaderos quimio y barorreceptores.
Es decir, serían las fibras nerviosas las capaces de captar los cambios, las
diferencias entre las presiones parciales de O2 o CO2 y las bajadas de pH. Por lo tanto
las céulas glómicas lo que hacen es sinaptar con las fibras nerviosas, para modular la
captación y la transmisión de ese estímulo.
Así pues estas fibras nerviosas, estos glomos, están en el origen de los reflejos
circulatorios, los cuales van a regular el tono muscular de los vasos, la frecuencia del
latido cardíaco y la frecuencia de ventilación. Por ejemplo, unas bajadas en las
presiones parciales de O2 o del pH sanguíneo o una subida de la presión parcial del CO 2,
provocan un reflejo que va a conducir al aumento en la frecuencia de ventilación.
Cabe mencionar que estas estructuras sensitivas se han relacionado con la
percepción del dolor que sufren los pacientes cuando están padeciendo una angina de
pecho o una isquemia cardíaca.
Inervación cardiáca
Debemos recordar que el corazón es capaz de producir contracciones por sí
mismo pero que el Sistema Nervioso Vegetativo Autónomo es quien regula esa
frecuencia cardíaca.
En concreto el Sistema Nervioso Simpático lo que va a hacer es acelerar la
frecuencia cardíaca mientras que el Sistema Nervioso Parasimpático va a ralentizarla.
Existen fibras nerviosas simpáticas a nivel de los nódulos Sinusal y
Auriculoventricular y también a nivel del miocardio. Incluso hay fibras simpáticas
capaces de atravesar el epicardio para inervar las arterias coronarias.
También podemos encontrar fibras parasimpáticas las cuales inervan los nódulos
Sinusal y Atrioventricular y el miocardio.
Podemos localizar ganglios nerviosos tanto en los plexos que se encuentran en la
base del corazón como en las paredes auriculares.
56
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
SISTEMA INMUNE O LINFOIDE
FUNCIONES
x Inmunidad del organismo frente a moléculas extrañas
x Respuesta inmunológica defensiva
GENERALIDADES
Las células del SI son:
x Células madre linfocitarias
x Linfocitos
x Macrófagos
x Células dendríticas
x Neutrófilos
x Esosinófilos
x Mastocitos
x Basófilos
Las células del SI se encuentran en las
huecas de la malla o red creada por las
células de sostén (estroma en timo de tejido epitelial) ¿?. Además, Existen componentes
solubles, como el complemento, citosinas…
El tejido linfoide es un conjuntivo reticular en el que la célula principal es el linfocito.
Los linfocitos pueden ser circulantes (en sangre o linfa) e intraepiteliales. Diferenciamos
dos tipos de tejido linfoide:
x Difuso: linfocitos distribuidos de forma laxa en el conjuntivo.
x Folicular o nodular: linfocitos agrupados en cúmulos densos esféricos.
Los órganos linfoides implicados en el sistema de defensa del organismo pueden ser:
x Primarios: maduración de células madre linfocitarias a linfocitos no
comprometidos inmunocompertentes. Médula ósea y timo.
x Secundarios: reacciones inmunológicas. Bazo, amígdala, MALT.
Las propiedades del sistema inmune son:
1. Especificidad. Los dominios específicos de un antígeno son reconocidos por
linfocitos individuales.
2. Diversidad.
3. Memoria.
4. Autolimitación. La respuesta inmunitaria se estimula por un antígeno específico.
Cuando el antígeno se neutraliza o desaparece, la respuesta se interrumpe.
57
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
5. Tolerancia. Una respuesta inmunitaria busca la eliminación de los antígenos
ajenos, al tiempo que se sigue siendo «tolerante» frente a los antígenos propios.
TIMO
Situado en la región anterosuperior del tórax, tras el esternón. Su peso máximo
se alcanza en la infancia (50g). Tiene naturaleza linfoepitelial, con 2 lóbulos unidos por
tejidos conjuntivos. Es un órgano linfoide primario, lugar donde maduran los linfocitos
T procedentes de la médula ósea.
Una diferencia importante entre el ganglio linfático y el bazo es que el estroma del timo
corresponde a células epiteliales tímicas organizadas en una red dispersa para posibilitar
un contacto estrecho con los timocitos en desarrollo, los precursores de los linfocitos T
procedentes de la médula ósea. A diferencia de lo que sucede en el timo, el estroma del
ganglio y del bazo contiene células reticulares y fibras reticulares, pero no células
epiteliales.
La extirpación experimental del timo en ratones recién nacidos lleva a una disminución
brusca de LT y a incapacidad de llevar a cabo reacciones inmunológicas celulares.
Afecta a la producción de anticuerpos, llevando a un deterioro crónico y a la muerte.
Origen embriológico
En la 6ª semana de vida, empieza el desarrollo de los primordios pares de los epitelios
ectodérmico y endodérmico del 3er surco branquial (= arco faríngeo). La paratiroides
deriva también del 3er surco branquial. La conexión se pierde entre los primordios y se
independizan, teniendo al final pequeñas masas sólidas.
Una vez que las masas se han independizado, el mesénquima derivado de la cresta
neural forma cápsula y tabique (lobulación incompleta). Las células epiteliales se
disgregan y forman una red de células que tienen desmosomas y queratinas 5 y 8.
En la semana 14 llegan células madre hematopoyéricas desde el saco vitelino atraídas
por células epiteliales. Se produce la diferenciación a macrófagos y células
interdigitantes.
En la semana 17 hay una segunda oleada de células con linfocitos T. En el periodo fetal
el timo contiene LT derivados del hígado
Síndrome de DiGeorge: defecto congénito en desarrollo del timo. Interacción anómala
entre las células epiteliales y mesénquima. Ausencia de LT circulantes. Incapacidad
para respuestas inmunológicas mayor. Infecciones graves.
58
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Estructura histológica del timo
El timo está rodeado por una fina cápsula de tejido conjuntivo desde la cual parten
tabiques o trabéculas incompletas hacia el interior del parénquima del órgano. Las
trabéculas delimitan regiones de parénquima denominadas lobulillos tímicos, que no
59
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
son en realidad verdaderos lobulillos, sino casquetes de corteza sobre porciones de
tejido medular más profundo. Tanto la cápsula como las trabéculas contienen vasos
sanguíneos, vasos linfáticos aferentes y nervios.
Este órgano posee un estroma epitelial laxo, cuyas células epiteliales son estrelladas,
para posibilitar un contacto estrecho con los timocitos en desarrollo, que se hallan entre
los huecos que dejan las células estromales. En ello se diferencia de los ganglios y el
bazo, que poseen un estroma de células y fibras reticulares.
Un solo progenitor origina las células epiteliales tímicas corticales y medulares. El
factor de transcripción Foxn1 regula la diferenciación de las células tímicas corticales y
medulares, que comienza antes de la llegada de los linfoblastos (se verán en próximos
apartados). Esta diferenciación incluye la expresión de citoqueratinas y el
establecimiento de uniones celulares de tipo desmosoma. Una mutación del gen Foxn1
genera ratones desnudos y atímicos (dado que el gen Foxn1 regula también la
diferenciación de los queratinocitos de la piel).
El factor de transcripción AIRE permite, como veremos más adelante, la expresión de
proteínas propias específicas de tejido por las células epiteliales medulares.
Corteza
La porción externa del parénquima se denomina corteza, y es basófila (por tanto, la
vemos más oscura en las tinciones). Contiene:
x Células reticulares (epiteliorreticulares, recordemos que no son reticulares
verdaderas sino epiteliales tímicas): presentan forma estrellada (excepto las que
están junto a la cápsula, sus tabiques o los vasos sanguíneos, que son planas y
tienen lámina basal) por sus largas prolongaciones citoplásmicas, que se unen a
las de células contiguas mediante desmosomas (mácula adhaerens).
Tanto estas prolongaciones como el cuerpo celular presentan numerosos filamentos
intermedios o tonofilamentos de tipo K8.
Son pálidas y poseen un núcleo grande en el que se distinguen 1 o 2 nucléolos.
Expresan tanto MHCI como MHCII, así como timopoyetina y timosina, moléculas que
participan en la educación de los timocitos todas ellas.
x Linfocitos: tan abundantes que impiden ver el resto de componentes
x Macrófagos y dendríticas interdigitantes:
también expresan MHCI y MHCII, y son
claves en la diferenciación linfocitaria.
También tienen función fagocítica,
especialmente los macrófagos, que
eliminan los linfocitos T que han sufrido
apoptosis en la corteza.
x Fibras colágenas (nunca reticulares)
x Miocitos estriados: se cree que este tipo
celular deriva de la cresta neural, y se está
investigando su posible relación con la
60
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
miastenia gravis, frente a la cual se pueden realizar extirpaciones tímicas.
La irrigación de la corteza corre a cargo de un sistema capilar derivado de las arteriolas
corticomedulares. Llegados a este punto, cabe destacar la importancia de la barrera
hematotímica, compuesta por el endotelio de los capilares y una doble lámina basal: una
capa de esta lámina es la propia membrana basal endotelial y la otra es la lámina basal
de las células epiteliorreticulares planas que ya se han descrito alrededor de los
capilares. Los macrófagos adyacentes garantizan que los Ag que se escapan de esta
barrera no reaccionen con los linfocitos T en desarrollo.
Médula
Sus componentes son los mismos que los de la corteza, aunque hay sutiles diferencias:
x Células reticulares: presentan forma estrellada por sus largas prolongaciones
citoplásmicas, que se unen a las de células contiguas mediante desmosomas
(mácula adhaerens). Tanto estas prolongaciones como el cuerpo celular
presentan numerosos filamentos intermedios o tonofilamentos de tipo K5. Sus
características tintoriales son similares a las de las células epiteliorreticulares
corticales. Expresan una citoquina denominada linfopoyetina estromal, capaz de
estimular a las células dendríticas.
x Corpúsculos de Hassall: no tienen que aparecer necesariamente, aunque su
número aumenta con la edad. Son masas aisladas de células epiteliorreticulares
eosinófilas muy juntas, que poseen núcleos aplanados y se disponen en capas
concéntricas. En su centro pueden hallarse indicios de cornificación.
x Macrófagos: son más abundantes en la médula
x Células dendríticas: muy abundantes, especialmente en el límite corticomedular.
x Linfocitos: menos
El timo alcanza su peso máximo en la pubertad (50 g), y una máxima producción de
linfocitos. A partir de ese momento, sufre una brusca involución, apareciendo en él
tejido adiposo, y se reduce la producción de linfocitos T en el adulto. Se establecen las
estirpes de linfocitos T y la inmunidad se mantiene sin necesidad de producir linfocitos
T nuevos.
Vascularización
x Sanguínea:
o Las arterias penetran en la cápsula y discurren por las trabéculas. A
continuación, emiten ramificaciones perpendiculares en la unión
corticomedular, y estas vuelven a emitir lazos capilares hacia la corteza y
la médula. Los capilares son capilares continuos (se deduce que, si la
presencia de una barrera hematoencefálica es necesaria, los capilares no
pueden tener poros).
61
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
o El sistema venoso es opuesto, las vénulas postcapilares recogen la sangre
tanto de corteza como de médula, y las llevan a venas de la unión
corticomedular, las cuales convergen en venas perpendiculares que
penetran en las trabéculas y atraviesan la cápsula.
x Linfática: hallamos capilares en la unión corticomedular, que drenan en los
vasos linfáticos de las trabéculas.
Diferenciación de los linfocitos T
La mayor parte de este proceso de diferenciación tiene lugar en la corteza, donde se
hallan el 85% de los linfocitos T. La mayor parte de los linfocitos sufrirán apoptosis, y
solo unos pocos conseguirán sobrevivir y madurar en la médula tímica convirtiéndose
en linfocitos inmunocompetentes.
El proceso empieza cuando los linfoblastos, precursores de los linfocitos T, llegan a la
corteza a través de los vasos corticomedulares. Se trata de células dobles negativas
(CD8- y CD4-) de unas 15 micras de tamaño que van a migrar desde la unión
corticomedular hacia la zona subcapsular. Simultáneamente a esta ascensión, se produce
la proliferación y reordenación génica. Esta última está ligada a la expresión de las
enzimas Rag y Tdt, que posibilitan el proceso, consistente en:
1. Expresión de la cadena beta
2. Expresión de la cadena pre-alfa
3. Formación de un pre-TCR
4. Expresión de la cadena alfa
5. Expresión de bajos niveles de TCR
En la parte profunda de la corteza, los linfocitos T en maduración son dobles positivos,
y se hacen receptivos a los complejos MHC-péptido, a la vez que dejan de proliferar y
aumentan la expresión de TCR y lo estabilizan. El proceso de selección positiva de los
linfocitos T empieza en presencia de las células epiteliales tímicas corticales, que
expresan moléculas MHC de ambos tipos. MHCI es necesaria para el desarrollo de CD8
simples positivos, y MHCII para el desarrollo de CD4 simples positivos. Los linfocitos
T capaces de reconocer el conjunto péptido-MHC propia progresarán a la selección
negativa, y los que no serán eliminados por apoptosis.
El gen aire permite la producción de autoantígenos mediante la regulación de la
expresión de genes específicos de tejido. Las células epiteliales tímicas medulares se
encargan de expresar estos autoantígenos unidos a sus MHC, de modo que si un
linfocito reconoce el conjunto con alta afinidad va a ser eliminado. De esta forma se
promueve la autotolerancia. Sin embargo, los linfocitos autotolerantes generados no son
aún inmunocompetentes, han de permanecer en la médula una semana más
aproximadamente, donde sufren una maduración definitiva dependiente de citocinas.
Los linfocitos T funcionales entrarán en las vénulas poscapilares de la unión
corticomedular y abandonarán el timo.
62
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Clínica: APECED o APS-1
Se trata de un trastorno autosómico humano caracterizado por la destrucción
autoinmunitaria de los órganos endocrinos, la incapacidad para eliminar las infecciones
por el hongo Candida y el desarrollo de tejido ectodérmico distrófico. La producción de
Ac específics frente a tejido y una reacción inflamatoria limitada a estructuras
específicas de órganos como la retina, ovario, testículo, páncreas o estómago está
asociada a mutaciones en el gen aire.
GANGLIOS LINFÁTICOS
Generalidades
Se trata de órganos que se suelen hallar agrupados, y cuya función es la filtración de la
linfa. En ellos, los linfocitos contactan con el Ag. Se calcula que tenemos entre 500 y
700 ganglios en el cuerpo humano, aunque los grupos más destacables se hallan en la
región mediastínica, inguinal, paraaórtica y mesentérica.
Los ganglios tienen forma de habas aplanadas. Su tamaño es variable, entre 0’2 y 2 cm,
y aumenta con la edad y el número de enfermedades padecidas. En cuanto a su
estructura básica, delimitamos tres zonas:
1. Cápsula: perforada por vasos aferentes
2. Corteza: basófila
3. Médula: pálida y eosinófila
63
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Vascularización
x Arteriovenosa: las arterias que vascularizan la médula penetran por el hilio,
alcanzan los cordones medulares y forman capilares. Las arteriolas que alcanzan
la corteza lo hacen a través de las trabéculas y forman una red capilar.
Posteriormente drenan por las vénulas postcapilares a través de la paracorteza
hasta la médula. Las venas vuelven a salir por el hilio.
x VEA: se localizan en la paracorteza, la parte más profunda de la corteza, y son
zonas de fácil acceso al ganglio para los linfocitos B y T debido a sus
propiedades relacionadas con el homing (las células endoteliales cúbicas de las
VEA expresan CD34, VCAM, ICAM 1 y 2 y selectina P entre otras moléculas,
que interaccionan con distintos marcadores de los linfocitos permitiendo la firme
unión de estos y su internalización). Recordemos que las VEA se hallan también
en otros órganos linfoides secundarios, como la amígdala palatina o las placas de
Peyer.
x Linfática: los vasos linfáticos aferentes atraviesan la cápsula y conducen la linfa
hacia el seno subcapsular. Los senos paratrabeculares se originan en el
subcapsular, y entran en la corteza. Se extienden hacia la médula en forma de
senos medulares ramificados rodeados de cordones medulares. Los senos
medulares ramificados confluyen con los subcapsulares a la altura del hilio y
atraviesan la cápsula para unirse al vaso linfático eferente. En ocasiones, la linfa
que entra en el seno subcapsular puede evitar los demás y salir directamente por
el vaso linfático eferente.
Los senos son espacios de luz irregular, delimitados por células endoteliales con lámina
basal. En el lado interno de estos senos, el endotelio es más laxo y la lámina basal es
incompleta. En ellos podemos observar prolongaciones de células reticulares y
64
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
macrófagos, que contribuyen a ralentizar el flujo linfático, el cual es unidireccional
debido a tres razones:
1. Válvulas
2. Musculatura de los propios vasos linfáticos
3. Musculatura extrínseca
Estructura histológica
Un ganglio linfático está rodeado por una cápsula, y su parénquima se divide en una
corteza y una médula.
La cápsula corresponde a tejido conjuntivo denso irregular rodeado por tejido adiposo.
La cápsula del ganglio en su superficie convexa está perforada por numerosos vasos
linfáticos aferentes, lo cuales tienen válvulas para evitar el reflujo de linfa al ganglio.
Hay que mencionar que además tiene fundamentalmente fibras de colágeno,
fibroblastos y fibras elásticas.
Es un poco más gruesa en el hilio y es en él donde se encuentra la salida de las vías
eferentes. Desde la cápsula se emiten pequeños tabiques hacia el interior (donde
encontramos el estroma).
La corteza forma la porción externa del ganglio excepto a la altura del hilio. Consiste en
una masa densa del tejido linfático (malla reticular, células dendríticas, linfocitos,
plasmocitos y macrófagos) y senos linfáticos, que son conductos por los que circula la
linfa. La médula es la porción interna o profunda del ganglio linfático.
Frecuentemente asociado al ganglio, en la zona externa, hay grasa blanca. Y, por debajo
del seno subscapular, hay zona cortical externa. La cápsula emite trabéculas al interior
y a sus lados hay senos trabeculares.
Dentro de ganglio hay un estroma con células y fibras reticulares cuyo origen es NO
EPITELIAL. Para visualizar las fibras hay que recurrir a las impregnaciones de sales de
plata. Estas fibras se organizan en una malla, habiendo en sus huecos células libres:
principalmente linfocitos, pero también macrófagos, células dendríticas que pueden ser
interdigitantes o foliculares:
x Los macrófagos se encuentran tanto en corteza como en médula. Son
fundamentalmente fagocíticas, pues tienen que fagocitar muchos linfocitos que
han entrado en apoptosis. Actúan también como células presentadoras de
antígenos, que expresan MHC I, MHC II y moléculas coestimuladoras. Sin
embargo, los niveles de expresión de MHC II y moléculas coestimuladoras son
mucho menores que los de las células dendríticas, lo que los convierte en APC
menos eficientes. En cambio, tienen una capacidad enorme para la endocitosis y
la digestión de los materiales incorporados.
x Las células dendríticas interdigitantes, no son residentes en el ganglio. Sí es
cierto que llegan, pero si acuden es porque fuera del ganglio han captado un
antígeno y van a presentarlo (luego son APC). Sufren un proceso de
65
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
diferenciación, llegan a las vías linfátcas, entran en el ganglio y presentan el Ag.
Son más abundantes en la corteza interna que en la paracorteza (zona profunda
de la corteza) y presentan MHC II. Son esenciales para la interacción con
linfocitos T de memoria en paracorteza y esenciales en Ag intracelular.
x Las dendríticas foliculares están presentes en los folículos, son estrelladas y
forman una red (andamiaje) en el interior del folículo. Son distintas a las
anteriores, pues no funcionan como APC o tienen MHCII, pero tienen receptores
del complemento. Atraen hasta un folículo a los linfocitos B que se hayan
activado (lo cual ocurre en la paracorteza) y que están fuera del nódulo, siendo
las encargadas de introducirlas dentro. En su mayor parte, los linfocitos B van a
morir y son las foliculares las que salvan a los que sobreviven. Otra
característica es que tienen una larga vida y son capaces de exponer en su
superficie complejos Ag-Ab.
La corteza y la médula son difíciles de separar, pues no hay un límite nítido. La corteza
es más externa, basófila y tiene más densidad de linfocitos. En la corteza hay dos zonas,
la externa donde están los nódulos o folículos con tejido linfoide difuso y la zona
interna o paracorteza, la cual carece de nódulos. En la médula hay muchos senos
linfáticos (por eso es tan pálida).
Corteza
En la corteza encontramos dos zonas: una más externa donde encontramos los folículos
o nódulos entre los cuales hay linfocitos T (tejido linfoide interfolicular difuso) y otra
interna/paracorteza donde no hay folículos. No existe un límite claro entre una zona y
otra sino que es un paso progresivo.
Corteza externa
Los nódulos son agrupaciones más o menos esféricas donde predominan linfocitos B y
dendríticas foliculares. Se clasifican en 3 grupos: primario, secundario y terciario.
x Primarios: la masa es muy basófila y fácil de distinguir del tejido linfoide de los
lados que es mucho más difuso, en la cual no hay zonas o regiones sino que es
homogéneo. Los Linfocitos B que tenemos en su mayoría son no
comprometidos aunque también hay de memoria. Cuando ocurre una reacción
antigénica encontramos folículos secundarios en los cuales se pierde la
homogeneidad y encontramos una zona periférica (corona o mano) que rodea al
interior llamado entonces centro germinativo.
x Secundarios: SOLO aparecen tras una estimulación antigénica (reacción de
defensa) y ya no son homogéneos sino que poseen una zona externa que es la
corona o manto rodeando el centro germinativo.
El centro es mucho más pálido y encontramos linfocitos B proliferando, diferenciándose
o en apoptosis. En un centro germinativo encontramos por tanto esa malla formada por
las dendríticas foliculares, los linfocitos B y también en menor cantidad linfocitos T
66
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
helper, macrófagos y células reticulares. Hay que aclarar que no todo el centro
germinativo es homogéneo.
Dentro del centro germinativo, cuando la reacción está en marcha, distinguimos dos
zonas: una pálida donde encontramos células llamadas centrocitos y una más densa
donde encontramos linfocitos B en proliferación a los que se llama centroblastos.
Rodeando el centro germinativo tenemos la corona (tiene mayor grosor cuanto más se
aproxima a la cápsula), mucho más basófila, con linfocitos B pequeños vírgenes que no
se han activado, es decir, no han reaccionado con ningún Ag y expresan
inmunoglobulinas como IgM e IgD.
Cerca del folículo lo que vamos a encontrar es la llamada zona marginal que es rica en
B de memoria. Esta zona no forma parte del nódulo aunque está en contacto con él.
x Terciarios: Poco a poco se van degenerando y acaba desintegrándose.
Corteza profunda o paracorteza
Esta región contiene la mayor parte de los linfocitos T del ganglio linfático. A causa de
su dependencia del timo, la timectomía perinatal en los animales impide el buen
desarrollo de la paracorteza. Por este motivo la corteza profunda también se conoce
como corteza timodependiente.
En esta zona están las VEA, es la entrada masiva de linfocitos T (que se quedan cerca) y
B (migran a la corteza externa y se integran en algunos de los nódulos). Todo este
proceso es muy breve, los linfocitos están en esta zona pero luego se van, es todo un
proceso muy dinámico. Luego se quedan un tiempo y ya circulan.
Médula
Es la porción más profunda del ganglio linfático, consiste en cordones de tejido linfático
separados por senos linfáticos llamados senos medulares. Una red de células y fibras
reticulares atraviesa los cordones y los senos medulares y sirve como el armazón
(estroma) del parénquima. Además de las células reticulares, los cordones medulares
contienen linfocitos (en su mayor parte linfocitos B).
La zona interna es muy pálida debido a la menor densidad celular y a la gran presencia
de senos. Hay también células libres, linfocitos, macrófagos y plasmáticas. En algunos
ganglios hay un gran número de mastocitos. Las células plasmáticas están ahí porque se
da secreción de Anticuerpos. Van a los senos linfáticos medulares que se continúan por
los vasos que salen por el hilio.
Funciones de los ganglios
Los ganglios están intercalados en la trayectoria de un vaso linfático. La linfa va al
ganglio y fluye de manera lenta. La lentitud, facilita que si por ahí circulan agentes
patógenos, éstos se puedan localizar y fagocitarlos. No todos los microorganismos se
van a poder eliminar, pero la tasa es grande. Los ganglios no están dispersos, van uno
detrás de otro para minimizar la tasa de microorganismos que sobreviven.
67
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Cabe mencionar que las vías linfáticas son utilizadas por células cancerígenas, luego se
extirpan los ganglios para quitar la diseminación en el caso de algún tumor.
Por la linfa también van los antígenos. Todo esto va con una respuesta inmunológica,
pues se forman células específicas y van a migrar a los puntos de entrada de ese
antígeno. Parte de las células que no migran a los puntos de entrada del antígeno,
cuando lleguen a la zona, habrá células efectivas contra ese antígeno.
Luego las funciones de los ganglios son:
x Filtrar la linfa eliminando patógenos. De esta forma se pueden retener y después
ser eliminados por los macrófagos un 90% de los patógenos de ahí que la linfa
pase por varios ganglios que están en cadena, rebajando al máximo el porcentaje
de patógenos que sigan circulando.
x Generación de una respuesta inmunológica para la que los antígenos activan
linfocitos específicos contra el Ag en cuestión. Habitualmente se desencadenan
respuestas humorales y celulares y, como consecuencia de estas respuestas
defensiva, aparecen anticuerpos y células selectivas que circularan e irán al
bulbo de entrada de ese Ag y al resto de órganos linfoides (con vistas a que si
ese órgano le llega ese Ag ya haya células especificas activadas contra él).
Proceso de reacción inmunológica
Cuando un Ag llega al ganglio, normalmente es captado por APCs. Primero llegan los
linfocitos T helper. Su activación ocurre en la paracorteza. En pocas horas (48h), los
linfictos específicos se van a diferenciar en linfoblastos, más grandes y basófilos y con
una división muy activa. Estos se van dividiendo masivamente y en pocos días se da un
aumento de linfoblastos. Da dos tipos: linfocitos T de memoria y T helper.
Cuando el antígeno es intracelular, lo normal es que también se activen los linfocitos T
citotóxicos. Para la activación completa es necesaria la acción del T helper. Si se da, los
linfocitos citotóxicos sufren el proceso de activación, proliferación y formación de
nuevos citotóxicos que serán de memoria y efectores.
Al final se forman 4 poblaciones: 2 de memoria (1 linfocito T y uno citotóxico) y 2
efectores (1 linfocito T y otro citotóxico).
En muchos casos la reacción también incluye a los linfocitos B que son activados
directamente por el Ag y cuando esto ocurre decimos que se inicia la reacción de tipo
humoral. Esta activación también se da en la paracorteza (que se sitúan en los folículos
pero migran hasta allí o están entrando).
Necesitamos linfocitos B, pero al no haber APC, es necesario que los linfocitos B vayan
con los T helper. Se producen unos pequeños focos (primarios) donde hay linfoblastos
de tipo B proliferando, diferenciándose y produciendo por primera vez células
plasmáticas. Se encargan de la primera secreción de Ig (IgM e IgD) que salen de la
linfa.
68
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Cuando un Ag alcanza un ganglio en los 4 o 5 días tenemos un ganglio que está
creciendo porque en la paracorteza es posible observar pequeños focos de células T y B
en proliferación activadas específica contra ese Ag. Sobre el 5 día podemos observar en
sangre los primeros anticuerpos que se han formado a partir de células plasmáticas.
Fundamentalmente son IgM e IgG y se segregan directamente en paracorteza.
Para más eficacia, los plasmoblastos van de la paracorteza a la médula. Así se segregan
niveles elevados de Anticuerpos en el cuerpo. Los linfocitos B van a dejar la médula
una vez que hayan sido segregados muchos Anticuerpos y los linfocitos B van a la
corteza externa.
En la migración tienen un papel esencial las células dendríticas foliculares que se
encontraban en la paracorteza de la médula. Los linfocitos B llegan a la corteza externa
y se meten en algunos de los nódulos, así el nódulo primario pasa a ser secundario. Allí
encontraremos un centro germinativo. Cuando el linfocito B llega y se emite en un
nódulo se le llama centroblasto (se reduce la producción de IgD). Llegan millones.
El nódulo, que no está vacío sino lleno de linfocitos, empuja a los linfocitos a la
periferia del folículo secundario, al manto o corona. La mayor parte de linfocitos
encontrados son pequeños no activados y no produciendo reacción generalmente
presentan cantidad de IgG e IgM en su superficie
Los centroblastos se siguen multiplicando y diferenciando e interaccionan con las
células dendríticas foliculares, macrófagos y linfocitos T helper. Proceden de muy pocas
células que se activarán, por ello los centroblastos son en buena medida células
clónicas.
En el centro germinativo se distinguen dos zonas (2 polos). Los centroblastos quedan
en la zona más oscura del centro germinativo y son células que sufren hipermutaciones
somáticas. Cuando el proceso termina se les llama centrocito y se encuentra en el polo
opuesto. Gracias a eso se van a expresar receptores contra el Ag pero con diferente
grado de afinidad. EL 90% de los centrocitos originados mueren, pues se produce una
selección negativa la cual es limpiada por los macrófagos. Los que se apoptosan
muestran baja afinidad y con ello las células dendríticas foliculares determinan qué
centrocito sobrevive y cual no. Para que los centrocitos sobrevivan a esta selección
negativa no solo se necesita que los Rc de su membrana sean de gran afinidad sino que
haya contactos suficientes y adecuados con las dendríticas foliculares (que forman el
estroma y atraen a los linfoblastos y hacen que sobrevivan los centrocitos).
Un centrocito se sigue dividiendo y como consecuencia ya sí se tienen linfoctios B de
memoria y plasmáticas que segregan Anticuerpos.
Sobre las células de memoria hay que aclarar que algunas se quedan en el manto, pero
la mayoría se desplazan, salen y se encuentran en buena medida en distintos órganos,
para que en alguno momento estén para dar una reacción rápida. Las plasmáticas no se
69
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
quedan en los nódulos sino que van hasta la medula donde es el lugar idóneo para la
secreción de los Ac. (Hay una segregación masiva mucho mayor que la del principio)
Los plasmoblastos vuelven a la médula y allí segregan IgG e IgM tanto a la linfa como a
la sangre.
RECORDATORIO DE INMUNOLOGÍA: Se pueden producir variaciones de clase
(variación variable que no se modifica pero si la porción de la cadena pesada), dando
lugar a isotipos que tienen la misma especificidad.
Durante este proceso inmunológico el ganglio aumenta de tamaño, y cada folículo es
mayor, habiendo más número de ellos.
Durante este proceso también hay cambios: aumenta la tasa de proliferación de
linfocitos (los que no son específicos), aumenta su tasa de movilidad y circulación. Es
decir, la recirculación cambia (velocidad, cantidad y tiempo que permanecen en los
órganos).
Cuando cesa la estimulación del antígeno, los ganglios experimentan una disminución
del tamaño, pero no recobra el original, sino que se hace más pequeño. Además, la
mayor parte de los linfocitos mueren.
Clínica
x Linfadenitis: Es una infección de ganglios linfáticos. Aumenta el tamaño de los
ganglios y duele si se palpan, experimenta una gran cantidad de edema y una
gran cantidad de neutrófilos desde los vasos sanguíneos. Además los neutrófilos
aumentan su capacidad fagocítica.
x Linfoma: Son tumores de tejido linfoide. La mayor parte son de los linfocitos B,
en este caso en los linfomas, los ganglios aumentan de tamaño pero no duelen.
Se clasifican los linfomas en Hodgkin y No Hodgkin.
BAZO
Generalidades
El bazo es un órgano linfoide secundario del tamaño de un puño y unos 150-200g de
peso. Es intraperitoneal, y está situado en el hipocondrio izquierdo (cuadrante superior
izquierdo del abdomen). Está interpuesto en la circulación sanguínea y abundantemente
irrigado. Aunque cumple funciones de eliminación de células extrañas o dañadas y
detección de Ag circulantes, no es esencial para la vida, con lo que se puede extirpar
quirúrgicamente en caso de necesidad.
70
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
El peso y tamaño del bazo se pueden ver aumentados en exceso como asociación a
algunos cuadros clínicos, y hablaremos entonces de esplegnomegalia. En general, se
considera excesivo un peso de entre 300-350 g como mínimo.
Embriología
El bazo se origina alrededor de la quinta semana del desarrollo embrionario, por
diferenciación, migración y fusión de unas masas mesenquimatosas del mesogastrio
dorsal. Si estas masas mesenquimatosas no se fusionan el futuro individuo tendrá un
bazo accesorio de situación y tamaño variables, que podrá ser o no funcional o acarrear
problemas clínicos.
Constitución histológica del bazo
El bazo está formado por una cápsula y una pulpa, que podemos dividir en pulpa
roja y pulpa blanca.
Cápsula
Es más gruesa en la zona del hilio, al igual que la de los ganglios. Emite
trabéculas incompletas hacia el parénquima, de manera que las cámaras virtuales que
estos tabiques delimitan están comunicadas entre sí. Por las trabéculas discurren, como
veremos más adelante, arterias y venas. Esta cápsula es muy friable, esto es, tiene poca
rigidez (de la cual son responsables las mismas trabéculas), con lo que es fácil que se
rompa en caso de traumatismo.
La cápsula del bazo es de tejido conjuntivo denso, y contiene:
x Fibrocitos
x Colágeno de tipo 1
x Fibras elásticas: colaboran en la distensión del bazo, que se acomoda a los
cambios en el volumen sanguíneo. En seres humanos, el bazo no actúa como
reservorio sanguíneo, función que sí que tiene en algunas especies, para lo cual
dispone de células musculares que facilitan la contracción en caso de que se
haya de movilizar dicha reserva sanguínea. Pese a lo que se dice en algunos
textos, el bazo humano no tiene capacidad contráctil (o esto es lo que ha dicho la
profesora, Ross no opina igual).
Pulpa
La sustancia del bazo se llama pulpa esplénica, y como todos los órganos se diferencia
en parénquima y estroma. Este último es en el caso del bazo un estroma reticular, y si
disponemos de una preparación con tinciones argénticas podemos observar que las
fibras no tienen igual dirección en todas las zonas: el bazo puede dividirse desde el
punto de vista morfológico y funcional en pulpa roja y pulpa blanca, denominadas así
por su color en estado fresco, y las fibras tienen distinto patrón de distribución en estas
dos zonas.
Pulpa roja
71
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Ocupa un 75% de la superficie del bazo. Tanto en su estado fresco como en los
preparados histológicos es de color rojo, debido a su alto contenido en eritrocitos. En
esencia, la pulpa roja consiste en sinusoides esplénicos separados por cordones o placas
esplénicos o de Billroth.
x Sinusoides: recordemos que llamamos sinusoides a un tipo de capilares
irregulares de paredes finas. Las sinusoides esplénicas son especiales debido a
las características de su tejido endotelial. Las células endoteliales que lo
componen tienen forma de bastoncillos largos, con el eje mayor paralelo a la luz
del vaso, por lo que se denominan también costillas. Entre las células contiguas
hay pocos puntos de unión celular, con lo que se delimitan grandes espacios en
el endotelio que permiten a los eritrocitos y otras células sanguíneas existentes
en la luz de la sinusoide atravesarlos como si se tratase de un colador permeable.
La constitución de la lámina basal también colabora en este proceso: las
sinusoides carecen de una lámina basal continua, en su lugar hay bandas
anulares de material de la lámina basal que rodean los capilares como si fuesen
los aros de un tonel, perpendiculares a las células endoteliales, que se
entremezclan con fibras reticulares.
x Cordones de Billroth: están formados por una malla laxa de células y fibras
reticulares que contienen eritrocitos, granulocitos, linfocitos, macrófagos,
dendríticas y plasmocitos. Los macrófagos se hallan cerca del capilar, ya que
están implicados en la degradación eritrocitaria y el metabolismo del hierro.
Pulpa blanca
Constituye un 25% del bazo. Se encuentra dispersa en la pulpa roja, aunque por su color
es fácilmente identificable. En ella se introducen ramas de la arteria esplénica, que
reciben el nombre de arterias centrales, de manera que la pulpa blanca no es más que
una aglomeración de linfocitos alrededor de las arterias centrales, en su adventicia,
constituyendo lo que se denomina vaina linfática periarterial (PALS).
Su función es fundamentalmente inmunológica, y en relación con ella la pulpa blanca
presenta folículos, que suelen contener centros germinativos que, al igual que en otros
órganos linfáticos, se desarrollan conforme los linfocitos B atraviesan distintos estadios
de activación. En los seres humanos los centros germinales se desarrollan unas 24 horas
después de la exposición al Ag, y pueden adquirir un tamaño que los haga visibles a
simple vista, hasta 1 cm. Estos folículos tan grandes que aparecen de trecho en trecho se
conocen como nódulos de Malpigio.
Zona marginal
72
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Se trata de una zona ampliamente vascularizada y rica en linfocitos, que constituye la
transición entre la pulpa roja y el manto de los folículos de la pulpa blanca.
Vascularización del bazo
La arteria esplénica da una serie de ramas que se introducen en la cápsula por la zona
del hilio, y penetran en las trabéculas. Desde ahí van a ir a parar a la pulpa blanca,
donde se denominan arterias centrales (aunque ya son arteriolas) y están asociadas a
PALS. Las arterias centrales dan una serie de ramas no terminales, las arteriolas
radiales, que vascularizan los nódulos o folículos y van a parar también, directamente o
a través del interior de los nódulos como ya se ha dicho, a la zona marginal o seno
marginal.
Ya en la pulpa roja, las ramas terminales de las arterias centrales son las arterias
peniciladas, llamadas así por su configuración en forma de pincel. Estas arterias
peniciladas disponen de un endotelio cúbico y una sola capa de miocitos, y van a ir a
parar a los capilares envainados. El endotelio de estos capilares también es cúbico, y se
hallan rodeados de elementos conjuntivos como macrófagos y fibras reticulares,
denominados elipsoide. Los capilares envainados no se abren a ningún otro vaso, sino
directamente a los cordones esplénicos, con lo que ya hablamos de circulación abierta.
Desde ahí, seguirá el mismo recorrido que la parte de la sangre marginal que va a parar
a los cordones (justo debajo explicado).
En cuanto a la sangre que habíamos dejado en el seno marginal:
x Parte va a parar a la circulación abierta, esto es, a los cordones de Billroth. Allí,
los elementos sanguíneos se filtran, y los que atraviesen este filtro irán a parar a
las sinusoides (circulación cerrada).
x Parte va al seno cavernoso perimarginal e irá a los sinusoides (circulación
cerrada).
Desde las sinusoides parten vénulas que desembocan en venas, las cuales atraviesan las
trabéculas y salen finalmente por la zona del hilio.
73
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Los vasos de la zona marginal permiten la entrada y salida de células dendríticas y la
recirculación de linfocitos: los T irán a parar a PALS y los B mayoritariamente a
nódulos. De media, la recirculación supone unas 5 horas.
En un momento dado, el 90% del flujo en el bazo está representado por la circulación
cerrada, sin embargo, la mayor parte del volumen sanguíneo se almacena en la
circulación abierta. Se trata de una circulación con escasa capacidad regenerativa, razón
de más para que sean comunes las esplenectomías en caso de traumatismo.
En resumen:
x Pulpa roja
1. Estroma reticularÆ cordones de Billroth
2. Células libresÆ cordones de Billroth
3. Sinusoides
4. Arterias peniciladas
5. Capilares envainados
x Pulpa blanca
1. Arteria central
2. PALS
3. Centro germinativo
4. Corona o manto
El drenaje linfático se realiza a través de vasos linfáticos eferentes que van por la pulpa
blanca y las trabéculas y salen por el hilio.
74
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Funciones del bazo
Pulpa roja
x Hematopoyesis: el bazo es un órgano hematopoyético muy importante en el
segundo trimestre de gestación, especialmente en la eritropoyesis. De ello se
encarga la pulpa roja. Sin embargo, esta capacidad en la especie humana no se
conserva, se pierde progresivamente. Algunas patologías llevan asociada
metaplasia mieloide, que consiste en una recuperación de la capacidad
hematopoyética del bazo.
x Filtración: los macrófagos elipsoides asociados a los capilares envainados de la
pulpa roja fagocitan células anómalas y captan Ag, desempeñando la función de
filtración de la que ya se ha hablado en los cordones de Billroth.
x Destrucción de los hematíes: cuando los hematíes son defectuosos o han
superado su vida útil pierden deformabilidad, con lo que al atravesar los
sinusoides tienden a romperse. Los macrófagos los fagocitan y se encargan del
metabolismo del Fe: aunque parte se queda como inclusiones en su interior
(ferritina y hemosiderina) la mayoría se recicla y vuelve a la médula ósea para
colaborar en la eritropoyesis, unido a la proteína transportadora transferrina.
El grupo hemo es degradado a bilirrubina que viaja al hígado unido a la albúmina, Se
elimina por la biblis.
Alteraciones clínicas relacionadas:
o Anemia falciforme: Hematíe en forma de hoz (drepanocito) que se forma
por una mutación puntual en el gen para la hemoglobina.
Si el individuo es homocigoto para la mutación sólo forma drepanocitos. Si es
heterocigoto su sangre tendrá mezcla de hemoglobina normal (HbA) y hemoglobina
mutada (HbS). Los individuos homocigotos muestran anemia hemolítica sea cual sea la
presión de oxígeno. En heterocigotos la anemia y hemólisis aparece cuando baja la
presión de oxígeno.
Los drepanocitos obstruyen el flujo sanguíneo. Se adhieren al endotelio, con lo que
aumenta la resistencia periférica. Al agolparse en los sinusoides se produce la hemólisis
por la rotura de la membrana del drepanocito y precipitación de HbS. Los macrófagos
fagocitan los restos.
o Anemia esferocítica: Los eritrocitos son esféricos y no pueden atravesar
los capilares esplénicos. Se degradan sin ser viejos. Ocasiona anemia y
puede ser necesario extirpar el bazo.
x Eliminación de bacterias y virus presentes en sangre (sepsis)
o Bacterias reconocidas por macrófagos y eliminada de manera directa o
tras ser recubierta por proteínas del complemento o Ig.
o Impide infecciones respiratorias, renales, meningíticas…
o Las bacteriemias son más frecuentes tras esplenectomía, especialmente
en niños. Supone un riesgo vital porque las bacterias circulantes se
distribuyen por todo el organismo.
En adultos la esplenectomía no tiene consecuencias graves porque ya poseen Ac. Se
recomienda vacunar contra neumococos y profilaxis preventiva con antibióticos
x Reservorio de plaquetas:
o 1/3 de total plaquetaria está en el bazo: pool de reserva
75
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
o No se ha localizado exactamente
x Reservorio de hematíes en otros seres vivos, donde la cápsula tiene propiedades
contráctiles.
Pulpa blanca
x Inmunológica:
La pulpa blanca se comporta principalmente como un nódulo linfoide, que produce
clones de linfocitos B en presencia de linfocitos T derivados del PALS. Esta función
tiene especial importancia durante la bacteriemia (presencia de bacterias viables dentro
de la circulación) porque los macrófagos pueden atrapar estas bacterias y presentar sus
antígenos a los linfocitos esplénicos para estimular una respuesta inmunitaria específica.
Los antígenos penetran en el bazo desde la sangre (no desde la linfa como en el caso de
los ganglios linfáticos) y llegan a la pulpa blanca a través de la arteria trabecular hasta
alcanzar la arteria central y el seno marginal. La arteria central y el seno marginal están
unidos entre ellos mediante arteriolas radiales.
Las células presentadoras de antígenos en la región de la corona detectan los patógenos
hematógenos, que son muestreados por las células derivadas del PALS. Los linfocitos T
interaccionan con los linfocitos B y esta interacción condiciona la proliferación de estos
últimos y su diferenciación a células plasmáticas.
Las células plasmáticas liberan inmunoglobulinas a la circulación sanguínea.
A modo de resumen:
o Macrófagos, linfocitos y CPA actúan detectando Ag que llegan vía
sangre
o El órgano linfoide secundario reaccionan ante ellos
o Ag captado por células dendríticas interdigitantes
o Migran a PALS
o Activan linfocitos T y B
o Se forman focos de linfocitos activados
o Migran a folículos primarios y originan centros germinales
TEJIDO LINFOIDE ASOCIADO A MUCOSAS (MALT)
Parte del sistema inmune que localizamos en las mucosas de órganos que son
vías de entrada:
x Tubo digestivo
x Vías aéreas
x Tracto urogenital
x FAE (epitelios asociados a folículos)
x SALT (piel)
76
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Componentes
Según su concentración, los linfocitos se encuentran:
x Linfocitos dispersos intraepiteliales
x Cúmulos difusos
x Folículos (por ejemplo en las criptas de Lieberkuhn)
x Amígdalas, placas de Peyer
Hay una enorme cantidad de células plasmáticas, por lo que la producción de Ac es muy
elevada, especialmente la de IgA (10g/día), pero también encontramos IgG, IgM e IgE.
Estructura histológica
No existen vías linfáticas aferentes, las uniones oclusivas del epitelio impiden
que los Ag lo atraviesen. Las células M permiten el transporte de Ag desde la luz hasta
el tejido linfoide adyacente.
Las células M son epiteliales, que provienen y se regeneran de las células madre
epiteliales. Forman parte de las FAE, pues se sitúan sobre nódulos linfoides. Pueden
estar en el epitelio del tubo digestivo, las vías urinarias y las vías aéreas. Se caracterizan
porque carecen de vellosidades, pero su membrana plasmática posee micropliegues en
la membrana apical y en la basolateral, donde son tan profundos que se les conoce como
invaginaciones o bolsillo intraepiteliales. En estos bolsillo albergan LB, LT y
prolongaciones de macrófagos y dendríticas. Además, expresan MHCII.
Las células M captan microorganismos y macromoléculas por endocitosis. Estas
viajan en vesículas por catepsina B y las vierten por la membrana basolateral. Se
comportan, por tanto, como células presentadoras de Ag a LB, macrófagos y
dendríticas. Este mecanismo puede ser usado por distintos patógenos como vía de
entrada.
Una vez que los Ag son expuestos y estimulan a las células, los LB migran a los
folículos y regiones parafoliculares y desarrollan plasmocitos que invaden toda la
mucosa del órgano, alcanzando incluso la de otros órganos y las glándulas exocrinas.
Esto permite una síntesis masiva de IgA.
Amígdalas
Acumulaciones masivas de nódulos linfáticos que se sitúa alrededor del istmo de
las fauces, en la orofaringe y en la rinofaringe. Forma el anillo de Waldeyer o
amigdalino, un anillo protector situado en la región anatómica inicial de los aparatos
digestivo y respiratorio. Está formado por:
x Amígdala lingual: cara dorsal de la raíz lingual.
x Amígdalas palatinas: a cada lado del istmo de las fauces.
x Amígdalas tubáricas: en las paredes laterales de la rinofaringe, tras la
desembocadura de las trompas de Eustaquio.
77
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Amígdala faríngea o adenoides: techo de la rinofaringe.
Estructura histológica
Son zonas con una superficie irregular y hendida por la presencia de criptas profundas y
abombamientos. El epitelio es difícil de diferenciar porque está invadido por linfocitos y
leucocitos. La luz de las criptas está taponada por moco y células epiteliales y
leucocitos.
Posee una cápsula de tejido conjuntivo que las delimita, haciendo fácil su extirpación.
Hay dos regiones: B (folículos) y T (parafolicular). Tienen VEA y linfáticos eferentes
que conducen la linfa hasta los ganglio linfáticos.
Amígdala lingual
Se encuentra en la mucosa de la cara dorsal de la raíz lingual. Es un epitelio
plano estratificado no queratinizado invadido por linfocitos, con criptas relativamente
cortas, poco ramificadas y separadas. Las glándulas salivales mucosas desembocan en
las criptas. Por debajo encontramos el músculo lingual.
Cada cripta está asociada a tejido linfoide difuso y nódulos linfoides, esto es un
folículo lingual. Entre los folículos linguales quedan bandas anchas de tejido conjuntivo
en conexión con la cápsula.
Amígdalas palatinas
Son pares, y están a izquierda y derecha de la entrada de la faringe. Es un
epitelio plano estratificado no queratinizado y grueso (1cm) con criptas profundas,
ramificadas y próximas; los linfocitos invaden tanto el epitelio como las criptas.
Su cápsula es gruesa y emite tabiques que lobulan. Las glándulas salivales
mucosas desembocan en la superficie, fuera de la cápsula. Asociado se encuentra el
músculo faríngeo.
Amígdala faríngea
Impar, en la mucosa del techo de la rinofaringe, adherida al periostio. Es un
epitelio respiratorio con parches de malpighiano invadido por linfocitos.
No hay criptas, pero sí pliegues y recesos irregulares. La cápsula presenta
abundantes fibras elásticas. Las glándulas seromucosas desembocan en la superficie.
Se inflama con facilidad, dificultando la respiración. Son extirpables, son las
denominadas vegetaciones.
GALT
Tejido linfoide asociado al tubo digestivo. La unidad se llama GALU. Las placas
de Peyer son engrosamientos de 2 cm de la mucosa y de la submucosa del íleon
terminal. Hay estructuras similares en el intestino delgado, en el apéndice y el colon
78
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Placas de Peyer
Se originan por el elevado número de folículos y tejido parafolicular en la región
opuesta a la inserción del mesenterio, normalmente. En niños puedes llegar a 300
folículos.
La estructura histológica está alterada: las criptas son escasas y la muscularis
mucosa está disgregada, invadiendo la submucosa. El epitelio intestinal que las cubre
carece de caliciformes. Los enterocitos cumplen menos funciones y degradan Ag en
lisosomas.
El epitelio forma prominencias aplanadas (epitelio cupular). Este FAE es
especial porque contiene gran cantidad de células M y de linfocitos Tc.
La región cúpular está formada por el epitelio y la cúpula, que es una banda
delgada de lámina propia con abundantes LB (no IgD), además de macrófagos,
dendríticas, plasmocitos y LT.
La placa está formada por la cúpula y el centro germinal, que contiene LB (IgA),
LT CD4+ y CPA. Los plasmoblastos surgen en el centro germinal y migran fuera. Los
plasmocitos colonizan toda la mucosa e incluso la mucosa de otros órganos como las
glándulas exocrinas.
Se segregan IgA, IgM e IgE. La IgE se une a mastocitos y se libera histamina.
La IgA entra por la lámina apical de enterocitos y sale por la basal como sIgA. La sIgA
se une a toxinas, Ag y microorganismos, que impide que se adhieran a enterocitos.
En la enteritis se producen procesos inflamatorios intestinales, en la que las
placas de Peyer reaccionan agrandándose y con activación del tejido linfoide.
79
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
BALT
Tejido linfoide asociado a las vías aéreas, su unidad es el BALU. Los folículos
son menos abundantes que en el tracto digestivo, pero hay muchos LB y LT dispersos.
SALT
Tejido linfoide asociado a la piel. La piel forma parte de la inmunidad innata
gracias a la capa córnea epidérmica. También forma parte de la inmunidad adaptativa.
En la piel hay linfocitos intradérmicos, células de Langerhans, queratinocitos, linfocitos
y macrófagos en la dermis.
80
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
SISTEMA TEGUMENTARIO
La piel es un órgano importante pues supone el 16 % del peso corporal. Si se pudiera
extender tendríamos 2 m2, pues es una superficie con zonas mucocutáneas. El grosor de
la piel es muy variable, puede ser de 0,5 mm a 3-4mm.
Es de fácil exploración y abordaje en biopsia. Además permite diagnosticar
enfermedades típicas de la piel y ver síntomas de otras enfermedades.
HISTOLOGÍA
La piel es sencilla, pues sólo tiene dos capas:
x Epidermis: La más externa, de origen ectodérmico y naturaleza epitelial
x Dermis o corion: más interna, de origen mesodérmico. Es conjuntivo.
Estas dos capas descansan sobre hipodermis o tejido subcutáneo (fascia superficial) y
ciertos anejos cutáneos pueden asentarse no sólo en el corion sino también en tejido
subcutáneo.
Hay que recalcar que el límite ente dermis y epidermis es irregular y muy marcado, sólo
pudiéndose observar un aumento de tejido adiposo.
FUNCIONES
x Protección:
o Mecánica: de cualquier daño físico, roce o fricción la piel protege toda la
superficie corporal.
o Química: protege de deshidratarnos, pues es vital mantener el equilibrio
hídrico y un buen nivel de hidratación. Es por eso por lo que en bebés es
peligrosa la deshidratación, ya que su barrera no está totalmente formada.
o Física: Por ejemplo de los rayos ultravioleta.
o Biológica: hay todo un sistema de vigilancia inmunológica
x Funciones metabólicas:
o Síntesis de Vitamina D: para muchos una hormona.
81
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
o Se excretan muchos catabolitos (agua y sales).
x Termorrecepción
x Termorregulación: Somos seres homeotermos y por ello debe haber un control
de la temperatura, a nivel local puede ser controlada por mecanismos que se
encuentran en la piel.
x Receptor de estímulos: La piel es el mayor órgano sensitivo que tenemos: recibe
calor, frio, cambios de presión…
PROPIEDADES
A través de ella se pueden absorber sustancias siempre que sean liposolubles o solubles
en alcohol. La piel, debido a su estructura histológica es buena para tratamientos tópicos
(cremas, pomadas, que atraviesan la piel).
La piel se suele clasificar en dos tipos: fina (o lampiña) y gruesa (glabra).
Histológicamente se considera que la fina es la epidermis, mientras que para los
dermatólogos la fina es la dermis.
En realidad, podemos distinguir 5 tipos de piel en función de la epidermis, de la dermis
y de los anejos:
x Lampiña (fina): Es la que tiene mayor extensión, de ella destaca que la dermis es
más gruesa. Es muy armónica, pues tiene gran cantidad de anejos, pero ninguno
predomina. No hay glándulas apocrinas, pues estas sólo existen en la piel
apocrina.
x Glabra (gruesa): Con extensión pequeña (en plantas de manos y pies). Tiene un
gran desarrollo de la epidermis y sobre todo del estrato córneo. No tiene
glándulas sebáceas y apocrinas, pero sí un gran número de ecrinas. Tiene una
altísima inervación que incluye tanto terminaciones nerviosas libres como
circulantes.
82
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Pilosa: el anejo principal son los folículos pilosos (numerosos y de donde crece
pelo largo y grueso) y también las glándulas sebáceas.
x Serborreica: Muchas glándulas sebáceas y con muchos folículos (de estos en vez
de pelo sale vello).
x Apocrina: lo principal son las glándulas apocrinas. Tiene pocas ecrinas, no
tantas sebáceas pero posee folículos pilosos abundantes. Dependiendo del sexo
puede salir vello largo o grueso, también depende de la edad. Las terminaciones
libres también son muy abundantes.
Superficie cutánea
Esta superficie no es lisa, sino que
muestra las arrugas epidérmicas (irregularidades, surcos, elevaciones, pliegues…). Esas
arrugas son características del individuo; son únicas, es genético y sirve como valor
diagnóstico (para medicina legal pues se utiliza para las huellas dactilares, palmares…).
Gente con trastornos hereditarios tiene alteraciones en sus huellas.
¿Por qué se presentan esas arrugas? Es un reflejo de la línea irregular que separa
epidermis de dermis, de la unión dermoepidérmica. Es irregular ya que la epidermis va
para abajo (formando las crestas epidérmicas) y la dermis va hacia arriba (formando las
papilas dérmicas).
Esta superficie irregular proporciona una mayor superficie para una mayor fijación
frente a tracciones mecánicas. De esta forma, en estas papilas dérmicas los vasos
pueden alcanzar niveles más superiores lo que es importante cuando la epidermis tiene
un grosor considerable (en la gruesa la superficie es mayor y está completamente lleno
de crestas y papilas).
83
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
DERMIS
Es tejido conjuntivo, de origen mesodérmico. Aparte de albergar los anejos cutáneos da
sostén. En ella encontramos los vasos y nervios. Histológicamente podemos distinguir
dos estratos:
x Estrato papilar: inmediatamente por debajo de la epidermis, se emite por las
papilas. Hay dermis papilar alrededor de cada uno de los anejos pilosos. Todo lo
demás corresponde a dermis reticular. Con el tejido conjuntivo laxo, rodea cada
uno de los anejos.
La superficie de contacto es estabilizada por los hemidesmosomas, que anclan los
queratinocitos basales con la lámina basal.
Hay predominio de la sustancia fundamental (destacando el ácido hialurónico y
condroitin sulfato). Además podemos encontrar fibrillas de colágeno tipo I y III.
Presenta un sistema fibrilar elástico complejo (con fibras de oxitalán, elaunina y
elásticas); para el anclaje epidermis-dermis.
x Estrato reticular: mucho más denso, con tejido conjuntivo denso. Posee fibras
elásticas más gruesas y el colágeno se encuentra incluso en haces. Las fibras
elásticas otorgan elasticidad y flexibilidad. El colágeno I da sostén y soporte
mecánico, con grandes haces y en todas direcciones. Junto a las fibras, hay
células libres (células típicas del SI).
En menor medida hay miocitos, siendo en su mayor parte lisos. Suelen estar dispersos,
formando a veces pequeños haces (el músculo horripilador). En algunas zonas hay una
gran cantidad de miocitos lisos (como en los genitales externos); en otras zonas hay
musculatura esquelética (cuello, cara, párpados…) que presenta en el corion miocitos
esqueléticos. A eso se le conoce como musculatura facial, mímica o de la expresión.
Debajo de la piel del corion, está el tejido subcutáneo (hipodermis) en el que aumenta el
número de adipocitos. Además encontramos la zona más profunda de los anejos. El
tejido subjuntivo se divide en lóbulos. Hay una distribución diferente según la edad y el
sexo.
La edad afecta a la piel, sobre todo a la dermis. Afecta especialmente al estrato reticular,
esto se debe a que se pierde agua, sustancia fundamental, fibras elásticas, aunque el
colágeno no disminuye tanto, se vuelve mucho más rígido y se pierde número de
células.
VASCULARIZACIÓN
La dermis está vascularizada por dos plexos:
x Plexo profundo que discurre por la dermis reticular.
x Plexo superficial que discurre por la dermis papilar del cual creemos que salen
una serie de asas que se meten en las papilas dérmicas de tal forma que la sangre
de esos vasos está mucho más próxima a la epidermis.
84
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
A partir de los vasos arteriales tenemos un primer plexo que está en la zona profunda de
la dermis reticular (frontera con la hipodermis) y la sangre sube hacia arriba hasta
alcanzar un plexo más superficial (plexo subpapilar) que inerva la dermis y las papilas.
Alrededor de cada folículo o glándula nos pintan un plexo papilar (cada anejo tiene
vasos y dominios capilares alrededor de él como apreciamos en los plexos
perifoliculares o periglandulares).
Clínica
La vascularización es importante en el diagnóstico de enfermedades inflamatorias de la
piel, por las infiltraciones diferenciales. Las enfermedades vasculares locales y
generalizadas pueden afectar a la red vascular cutánea. Las púrpuras no inflamatorias
(extravasación de sangre en la dermis a partir de vasos pequeños) pueden ser pequeñas
(petequias, menores de 3 mm de diámetro) o grandes (equimosis). Los trastornos de la
coagulación, las enfermedades de los eritrocitos (drepanocitosis) y los traumatismos son
causas frecuentes.
La urticaria aguda es una reacción transitoria debida a un aumento de la permeabilidad
vascular asociado a edema en la dermis. Las vasculitis son un conjunto de trastornos
caracterizados por inflamación y lesión de la pared vascular. La mayor parte de las
vasculitis cutáneas afectan a los vasos pequeños, principalmente a las vénulas.
Termorregulación
En cuanto a los vasos hay que ver que es frecuente anastomosis arteriovenosas que
permiten un cierto control de la temperatura a nivel local. De esta forma cuando las
anastomosis están cerradas la sangre va a los plexos más superficiales disipando calor
(termorregulación local). La limitación del flujo se realiza por el plexo superficial, para
reducir la pérdida de calor.
Glomos: Vienen de la zona reticular de lechos ungueales, almohadillas plantares o cutis
facial. Aquí encontramos anastomosis arteriovenosas, siendo una zona rica en la
inervación. Se debe hacer mención al Canal de Sucquet-Hoyer, que posee células
glómicas (miocitos lisos a modo de esfínteres). También se encuentran en la rama
venosa.
85
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Vascularización linfática
En la dermis encontramos abundantes vasos linfáticos que en principio carecen de
válvulas y cuesta muchas veces encontrarlos porque la luz esta ocluida. Es más típico/
fácil encontrarlas en inflamaciones o edemas (como en una muestra donde el paciente
tiene urticaria en la que los vasos están drenando y están dilatados). Recogen el exceso
de líquido intersticial. La linfa va a ser transportada hasta los ganglios y existen unas
células en la epidermis llamadas células de Langerhans que son capaces de migrar y
llegar hasta esos vasos linfáticos de manera que a través de la linfa que segrega la piel
pueden alcanzar ganglios.
UNIÓN DERMOEPIDÉRMICA
No hace referencia solo a lámina basal de dermis y epidermis sino también incluye a la
membrana basal de queratinocitos de la capa basal de la epidermis. Es muy irregular y
cumple muchas funciones:
x Cohesión entre epidermis y dermis: es fundamental el anclaje de las dos capas
por las fuerzas de tracción.
x Polaridad de queratinocitos: un queratinocito al ser una célula epitelial está
polarizada y se organiza de manera muy estricta. Es la unión dermoepidérmica
quien manda a la célula cómo debe orientar sus polos, la organización del
citoesqueleto, de las proteínas de membrana…
x Emitir señales para el desarrollo de tejido en situaciones de daño y reparación
x Barrera Semipermeable: es quien decide quién pasa y cuando.
Lámina Basal
Está formada por 3 estratos diferentes:
x Lámina lúcida: Constituye el primer estrato. Ofrece cohesión de la mano de
glucoproteínas (laminina, nidogen y fibronectina). Estas se unen a receptores
(integrinas) que están en la membrana plasmática basal de los queratinocitos.
x Lámina densa: Tiene dos funciones: dar soporte y flexibilidad (gracias al
colágeno IV que se organiza formando redes y en menor medida encontramos
colágenos VII que contribuye a seguir proporcionando anclaje) y actuar como
86
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
barrera o filtro (intervienen los proteoglucanos sulfatados y es atravesada por
células en migración). Esta lámina densa hace una especie de criba de lo que
puede o no pasar.
x Lámina reticular: es la zona más profunda. Se encuentra en contacto con la
lámina basal de la dermis. Está formada por todo tipo de fibras que se organizan
en haces no muy gruesos. Posee fibras elásticas, de oxitalán y colágeno I, III, V
y VI. Se encarga de la acomodación ante deformaciones sin alterar la integridad
de la piel, no es tan de anclaje.
EPIDERMIS
Epitelio plano estratificado queratinizado de origen ectodérmico. Su grosor es de
0,4 a 1,5 mm (piel fina o labra). Origina faneras (anejos cutáneos) y tiene renovación
constante. Se encuentran 4 tipos celulares: queratinocitos, melanocitos, células de
Langerhans y células de Merkel.
Queratinocitos
Constituyen un 80% del epitelio.
Son células epiteliales. Se
denomina de este modo porque su
principal producto es la queratina,
una proteína de tipo filamento
intermedio.
Su diferenciación conlleva
cambios estructurales,
ultraestructurales y metabólicos
que conducen a la apoptosis. Sufre
desplazamientos, lo que hace que
estén organizadas en estratos:
Basal, Espinoso, Granuloso y
Córneo.
87
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Posee tonofilamentos. Hay regulación génica de los genes para queratinas, que varía con
la diferenciación celular e implica la presencia de diferentes filamentos de queratinas en
los distintos estratos epidermis y en distintos epitelios. Hay queratinas que se expresan
juntas.
x Epitelios simples: par 7-17 o 8-18.
x Epitelios estratificados: par 5-14, o en menor medida 6-16, en el estrato basal.
En las capas superiores están los pares 1-10 y el 2-10.
x A nivel del esófago el 4-14, y en la capa córnea 14 el 3-12.
La diferenciación implica un aumento de tamaño celular, tendencia al aplanamiento,
aparición de nuevos orgánulos, junto a la expresión génica de queratinas.
Se produce, por tanto, una reorganización ultraestructural, cambios metabólicos,
alteraciones de la membrana que llevan a su desaparición. Finalmente hay apoptosis y
deshidratación, quedando sacos repletos de filamentos de queratina y matriz proteica,
una envuelta confinada bajo la membrana y una matriz lipídica.
Estrato basal
1 hilera de células cúbicas sobre el estrato basal, basófilas y con abundantes orgánulos.
Hay K5/K14 y proteína ligadora de Ca2+, asegurando que los niveles de calcio sean
bajas.
Hay muchas uniones de adhesión (α3β1, α6β4, α2β1, α5β1) y desmosomas y
hemidesmosomas. Este estrato determina la polaridad. La lámina basal se ensambla con
la MEC. Entre los queratinocitos hay espacios, los canales faciales.
x 10% células madre
x 50% células amplificadoras: mantinen la capacidad de división
x 40% células postmitóticas
Aquí se originan todos los queratinocitos. La unidad proliferativa epidérmica se
considera como un queratinocito y todos sus derivados.
88
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Estrato espinoso
Hasta 10 capas de células. Los queratinocitos de la capa espinosa tienen una forma
poligonal aplanada con un núcleo ovoideo definido. Haces de filamentos intermedios,
las tonofibrillas, se extienden en las prolongaciones citoplasmáticas a modo de espinas y
se unen con la placa densa del desmosoma.
Posee uniones comunicantes conocidas como espinas (nódulo de Bizzozzero), canales
faciales, desmosomas y desmogleína 3.
Puede sufrir alteraciones en el que hay separación de queratinocitos: edema, reacción
inflamatoria y vesículas y ampollas.
Las queratinas son K1/K10. Esto genera una mayor resistencia a tracciones, cohesión
del estrato e impide la destrucción y descamación. En epidermis hiperproliferativa
(heridas, psoriasis, queratosis...) hay K6/K16.
Aquí se produce un aumento del tamaño celular y aparecen los cuerpos de Odland,
lamelares o queratinosomas en capas superiores. Son gránulos con membrana y
contenido lamelar: glucoproteínas, glucolípidos, fosfolípidos, esteroles e hidrolasas.
Estrato granuloso
Hasta 10 capas. La capa granulosa corresponde a un conjunto de múltiples capas de
queratinocitos aplanados con núcleos, que tienen en su interior unos característicos
gránulos de queratohialina, basófilos, de forma irregular sin membrana limitante y
asociados a tonofilamentos.
Hay fosforilación de K1/K10 y expresión de nuevos genes, apareciendo K2/K11. La
matriz es proteica, con filagrina, que agrega los filamentos intermedios de queratina en
haces densos, lo que justifica el aplanamiento de las células. Es el lugar donde tiene
lugar el embebemiento y empaquetamiento de los filamentos de queratina.
Además, se produce un aplanamiento celular. Se distinguen claudinas 1 y 4 (sirven de
marcadores) y aumenta la concentración de Ca2+ al desaparecer la proteína ligadora de
calcio.
Bajo la membrana plasmática hay una banda marginal, es una capa densa de proteínas y
lípidos con uniones covalentes, que finalmente dará lugar a la envuelta cornificada,
insoluble, que forma un esqueleto rígido. Sus componentes son:
x Involucrina, quertolinina, loricrina, elafina, repetina, desmoplaquina,
envoplaquina, periplaquina, cistatina α, pancornulinas, prot peq ricas en prol
x Involucrina, proteína citosólica que se une a proteína de membrana en regiones
interdesmosómicas
x Loricrina (gránulos L). Tardío. 70%
Los cuerpos de Odland ocupan un 15% del volumen celular. Los cuerpos laminares, que
aparecen en los queratinocitos de la capa espinosa, son más numerosos en la capa
granulosa y el producto laminar, el glucolípido acilglucosilceramida, se libera hacia los
espacios intercelulares. En el espacio intercelular, el material lipídico laminar forma una
estructura con múltiples capas que se organiza en sábanas extensas, que revisten la
89
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
superficie de los queratinocitos de la capa superior, el estrato lúcido. Se forma una
probarrera, que madurará para dar lugar a la barrera hidrofóbica.
Es este estrato, en resumen, tiene lugar:
x Fosforilación de filamentos de queratinas
x Exocitosis de cuerpos de Odland Dependientes de Ca2+
x Formación de banda marginal
x Apoptosis: EndoNasas, fragmentación del ADN, pérdida de núcleo y orgánulos,.
Está mediado por hidrolasas á´cidas, esterasas, fosfatasas, proteasas, ADNasas,
ARNasas... Este rpoceso está bloqueado en estratos inferiores (...).
La transición del granuloso al córneo implica pérdida de peso. El ascenso por el estrato
de Malpigio dura 14 días y es ascenso por el estrato córneo dura otros 14 días.
Estrato córneo
Corneocitos o escamas córneas. Sacos repletos de queratina con membrana reforzada
por envuelta cornificada y compactada por MEC muy hidrófobo. Son grandes,
aplanados, sin núcleo y muy eosinófilos.
No es un estrato inerte, hay mantenimiento de
actividades metabólicas:
x Anclaje de corneocitos
x Maduración de la envuelta cornificada
x Transformación del MEC
Por tanto, cumple importantes funciones;
x Permite la descamación
x Barrera mecánica, protege capas
inferiores
x Barrera hídrica, evita deshidratación
90
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
El estrato lúcido es refringente, por lo que a mover el óptico brilla, viéndose en la piel
glabra.
x Estrato coyunto o compacto: Es el estrato lúcido descrito en la piel glabra. Los
corneocitos mantienen desmosomas y desmogleína 1. Hay uniones herméticas
con claudinas 1 y 4 y la envuelta córnea está debilitada.
x Estrato disyuntivo o descamativo
Disolución de desmosomas. Se producen huecos en la MEC que da lugar a una red de
canales, que actúan como poros hidrófilos. La envuelta cornificada es muy rígida (16
nm) y se disuelve la membrana plasmática.
En la MEC se degradan azúcares y lípidos polares, por lo que hay predominio de lípidos
neutros, esteroles y ceramidas. Hay máxima cohesión y sirve de barrera hídrica.
Hay alternancia de capas MEC y envuelta rígida. Con la descamación hay disolución de
desmosomas con presencia de enzimas que disuelven los desmosomas, la membrana y
la matriz.
Trastornos ampollosos
Como ya hemos visto en toda su diferenciación hay varios procesos y en todo ello
podemos encontrar alteraciones:
x Trastornos ampollosos: Se forman Ig contra alguna de las moléculas de los
hemidesmosomas (plaquinas = BPAG1 o plectina, integrinas o proteínas
transmembranales de la M.Basal del queratinicito) –
x Trastornos de queratinización: afectan a los genes que codifican para las
queratinas.
o Epidermolisis ampollosa simple (EAS): Mutación de K5-K14 o
o Hiperqueratosis epidermolítica: Mutación en K1-K10
o Queratodermia epidermolitica palmoplantar (QEEP): Mutación en K9-
K11 o
o Ictiosis ampollosa de siemens (IAS):Mutación en K2
x Trastornos de la cubierta:
o Queratodermia palmoplantar estriada que afecta a las desmoplaquinas.
o Ictiosis lamelar: Enorme sequedad donde esta alterada la enzima
glutaminasa 1 que interviene en la formación de enlaces entre proteínas,
así, cuando la banda marginal no se forma adecuadamente aparece la
ictiosis laminar.
x Síndrome de Vohwinkel y eritroqueratodermia progresiva simétricacuando está
mutada la loricrina y provoca hiperqueratosis
x Ictiosis ligada a X por mutaciones en enzimas que modifican la banda marginal.
x Síndrome de Sjögren Larsson por mutaciones en enzimas que modifican la
banda marginal
91
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Manto ácido
Capa de escamas y MEC junto a sebo y sudor ecrino. Lubrifica y tiene un pH
ácido. Permite la presencia de gérmenes saprofitos, creando un microambiente adecuado
para la vigilancia inmunológica.
Células claras
Fueron bautizadas con ese nombre por Ranvier
debido a su cromofobia, ya que su citoplasma claro
aparecía sin teñir y destacaba entre los
queratinocitos. Se hallan en el estrato basal,
espinoso y granuloso. No se trata de un único tipo
celular, sino que hablar de células claras es hablar
de un concepto que engloba tres tipos de células:
1. Melanocitos
2. Células de Langerhans
3. Células de Merkel
Melanocitos
Los melanocitos son células estrelladas
derivadas de la cresta neural, que migran
desde allí a la epidermis. A las 10-11
semanas de VIU ya hallamos en la dermis,
pudiendo localizarlas en la epidermis entre
las semanas 12 y 14 del desarrollo
embrionario. Cabe destacar asimismo en
relación a ellos la interacción entre el factor
célula madre y el receptor c-kit.
Los melanocitos no son exclusivos de la epidermis, pero constituyen en ella un tipo
celular muy importante, localizado en el estrato basal. Aun así, debido a una
enfermedad o con estimulación prolongada podemos provocar el ascenso de los
melanocitos a capas superiores de la epidermis. Los somas de los melanocitos se hallan
por tanto pegados a la lámina basal, aunque sus prolongaciones o dendritas se proyectan
en sentido ascendente, de manera que se localizan en el estrato espinoso, entre los
queratinocitos.
Los melanocitos no poseen entre sí ni con los queratinocitos vecinos uniones de tipo
desmosoma ni hemidesmosomas, pero sí placa basal electrodensa. Carecen de
tonofilamentos, aunque sí tienen filamentos intermedios y muchos microtúbulos,
esenciales para el transporte de los melanosomas. Estos son los orgánulos
característicos de los melanocitos, encargados de la síntesis y el depósito de melanina,
que puede ser de dos tipos:
x Eumelanina: marrón
x Feomelanina: color rojizo
Un mismo individuo sintetiza los dos tipos de melanina, pero en distinta proporción.
Los melanosomas que contienen eumelanina son alargados y con matriz estriada. Los
92
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
melanosomas que contienen feomelanina son redondos y con matriz granular. En ambos
casos, su pH es ácido.
La melanina es argirófila, por lo que podemos visualizar los melanosomas que se hallan
tanto en los queratinocitos como en los melanocitos con técnicas de impregnación
argéntica. También los podemos ver mediante técnicas histoquímicas (tirosinasa
encargada de oxidar la tirosina a DOPA, que posteriormente será convertida a tirosina).
Algunos de los marcadores que podemos usar para localizar los melanosomas son:
S100, Melan-A, la glucoproteína HMB-45…
Las reacciones de síntesis de la melanina y formación de melanosomas maduros ocurren
en 4 fases. La maduración muestra cambios en matriz proteica.
1. Premelanosoma con tirosinasa: posee una estructura interna muy ordenada como
consecuencia de su contenido
2. Premelanosoma con membrana interna
3. Depósito de melanina en estriaciones unidas a la membrana interna
4. Melanosomas maduros electrodensos con diferente distribución de la membrana
interna
Transporte de melanosomas
Se trata de un transporte bidireccional vía microtúbulos, en el que tienen una
importancia vital las cinesinas. Como ya hemos dicho, los melanosomas van a acabar
siendo transferidos desde los melanocitos a los queratinocitos, para lo cual es
indispensable un proceso denominado retención periférica de los melanosomas,
controlado por las proteínas Rab27a, miosina-Va y melanofilina. En ausencia de esta
retención periférica, se desarrolla síndrome de Griscelli, un albinismo parcial que lleva
asociados trastornos neurológicos e inmunológicos.
La transferencia a los queratinocitos se realiza por un proceso denominado donación
pigmentaria, que comprende la fagocitosis del extremo de la prolongación melanocítica
por el queratinocito, y se trata en realidad de una secreción citocrina porque también se
fagocita una pequeña porción de citoplasma del melanocito existente alrededor del
melanosoma.
Una vez transferidos a los queratinocitos los melanosomas no se almacenan libres en el
citoplasma, sino que se engloban en lisosomas fagocíticos que contienen también
mucho citoplasma, debido al proceso de internalización ya descrito. En los individuos
de raza blanca, los melanosomas que hay dentro del lisosoma son pequeños, pero se
encuentran en gran cantidad; mientras que en las personas de raza negra a veces
encontramos un solo melanosoma, pero de un tamaño considerable. Su almacenamiento
en lisosomas implica que van a comenzar a ser degradados.
93
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
La melanina está destinada a la protección frente a la radiación, con lo que los
melanosomas se localizan en la zona supranuclear. Los queratinocitos con melanosomas
en personas de raza blanca se encuentran en todas las capas de la epidermis excepto en
la córnea, mientras que en individuos de raza negra sí que podemos encontrarlos en este
último estrato.
Concepto: unidad melanoepidérmica
Formada por un melanocito y los 36 queratinocitos con los que se relaciona debido al
transporte de melanosomas.
94
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Clínica
x Léntigos solares: son los lunares comunes. Se trata de acúmulos de melanocitos
hiperplásicos con una gran cantidad de melanina.
x Melanomas cutáneos: Constituyen solo un 4% de los cánceres de piel, pero un
80% de los fallecidos por cáncer de piel lo tenían de tipo melanoma, con lo que
es muy agresivo. Se relaciona con la sobreexpresión del factor MITF, que
interviene en el ciclo celular y estimula genes que participan en la producción de
melanina.
Coloración de la piel
El color de la piel en los seres humanos está determinado por la melanina. El color
constitutivo depende de:
x Número de melanosomas: estos no solo se hallan en los melanocitos, sino que se
transfieren a los queratinocitos. Por lo tanto, el número total de melanosomas
será la suma de los melanosomas que hallemos en queratinocitos y los que
encontremos en los melanocitos. Es distinto en las distintas razas.
x Velocidad de transporte de los melanosomas por microtúbulos
x Índice de melanogénesis
Este último factor se puede estimular, con lo que forma parte también junto con la
proliferación de los melanocitos (estimulable) de la tonalidad facultativa de la piel.
Existen diferencias zonales en el número de melanosomas en un individuo (además de
las interetnia). La cantidad de melanosomas y por tanto la tonalidad de la piel depende
de factores genéticos, y tiende a descender con la edad. En el embarazo y en
enfermedades endocrinas como la enfermedad de Addison se produce una
hiperpigmentación, que remite cuando lo hace el estímulo.
Células de Langerhans
Fueron descubiertas por este estudiante de
segundo de Medicina en 1868. Se visualizan
también con técnicas de impregnación argéntica,
histoquímicas (ATPasa) e inmunoquímicas
(marcadores: CD1a, CD46 y S100). Proceden de
precursores migratorios de la médula ósea que
alcanzan la epidermis entre la 6ª y la 10ª
semana, guiados por citoquinas.
Estas células se encuentran en el estrato espinoso, aunque no son mayoritarias, están en
bajo porcentaje. Dado que los queratinocitos se encuentran en proceso de ascenso, para
permanecer en esta capa las células de Langerhans se mueven contracorriente, no están
estáticas, si lo estuvieran serían desplazadas.
Se trata de células de aspecto dendrítico, con el soma estrellado, cuyas prolongaciones
se proyectan hacia los canales faciales existentes entre los queratinocitos. No presentan
desmosomas, sino uniones por cadherinas. Su citoplasma es electrolúcido, con un
núcleo lobulado y una gran densidad de orgánulos, entre los que destacan especialmente
los gránulos de Birbeck. Estos gránulos son vesículas de tamaño relativamente pequeño
95
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
que están aplanadas y se ven como bastoncillos con una expansión bulbosa en un
extremo (forma de raqueta de tenis). Contienen langerina, una lectina encargada del
reconocimiento de hidratos de carbono.
Las células de Langerhans se encuentran en todos los epitelios estratificados y en los
ganglios, no solo en el estrato espinoso, donde además tienen distinta densidad según la
zona. Son APCs, reconocen, internalizan y exponen el Ag. Cuando lo hacen, atraviesan
todas las capas hasta llegar a la dermis, donde se encuentran los vasos linfáticos, y
migran por ellos a los ganglios linfáticos, donde activan a los linfocitos desencadenando
la respuesta inmunológica: los linfocitos activados migran a la zona de procedencia de
la célula de Langerhans para la defensa (SALT de la piel, compuesto por células de
Langerhans, linfocitos, queratinocitos y vasos linfáticos). Algunos de ellos se quedan
como residentes (linfocitos intraepitelialesÆ relación con alergias de contacto). Por
tanto, estas células:
x Inducen respuesta específica por formación de LTc
x Rechazo en injertos
x Papel en defensa contra gérmenes
Cuando una célula de Langerhans reconoce a un Ag se producen en ella cambios
funcionales y fenotípicos:
x Aumento de tamaño
x Cambio de marcadores
x Pérdida de la capacidad presentadora: ya tienen un Ag
x Desaparición de las uniones por cadherinas y de los gránulos de Birbeck.
Clínica: psoriasis
La psoriasis es una enfermedad dermoepidérmica crónica caracterizada por:
x Hiperplasia persistente de la epidermis por una proliferación celular anormal
x Microcirculación anormal en el plexo capilar de la dermis. Los plexos que
rodean las papilas dérmicas adquieren predominancia venosa, con lo que las
células inflamatorias migran a la dermis. De ahí, estos linfocitos
(fundamentalmente neutrófilos) migran a la epidermis, donde se forman
microabscesos que desplazan la capa córnea.
96
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Inervación
Cuando hablamos de la inervación debemos distinguir dos componentes:
x Eferente: en el caso de la piel, corre a cargo de fibras amielínicas del sistema
nervioso simpático, que se dirigen a inervar las glándulas ecrinas y apocrinas, el
músculo piloerector y los vasos sanguíneos. Estas fibras nerviosas no terminan a
nivel de la dermis, sino que atraviesan la lámina basal y penetran entre los
queratinocitos de la epidermis (fundamentalmente los del estrato basal, pero
también los del estrato espinoso).
x Aferente: es la inervación sensitiva, en el caso de la piel es especialmente
importante porque es el órgano sensorial más extenso del cuerpo. En él reside la
información del tacto gracias a múltiples receptores, aunque hemos de recordar
que los receptores no son exclusivos de la piel, también los hallamos en
músculos, tendones, articulaciones y vísceras. Podemos clasificarlos en:
o Nociceptores: son los receptores del dolor. Podemos entender el dolor como
dolor mecánicos (aplastamiento, pinchazo) o térmico (aquel que sentimos a
temperaturas muy bajas, de 2-3 grados, o muy altas, de 43-45 grados). Estos
receptores no están encapsulados, y están en contacto con fibras mielínicas
para facilitar una respuesta inmediata. En una imagen a microscopía, los
observaremos como unas fibras que atraviesan la dermis y pasan a la zona
basal de la epidermis.
o Termorreceptores: hay dos tipos de termorreceptores, ambos encapsulados:
x Corpúsculos de Krause: detectan temperaturas mínimas, inferiores a
los 10 grados. Se hallan tanto en la dermis como en las mucosas de
otras zonas del cuerpo. Son esféricos y pequeños, y en ellos las fibras
amielínicas se acaban entremezclando con elementos conjuntivos.
x Corpúsculos de Ruffini: detectan temperaturas máximas, superiores a
25 grados. Se localizan en la dermis y en las articulaciones. Son
ovalados, y tienen un tamaño relativamente grande, de 1 mm. En
ellos, las fibras amielínicas también acaban mezclándose con
elementos conjuntivos.
o Mecanorreceptores: captan la presión, la deformación de la piel o el
movimiento de los folículos pilosos. Pueden ser:
x De adaptación lenta: envían información sin que haya movimiento, y
están constituidos por células de Merkel
x De adaptación rápida: solo mandan información si hay movimiento, y
están constituidos por corpúsculos.
Hallamos tres tipos de células que conforman los mecanorreceptores:
Células de Merkel
Se descubrieron utilizando impregnaciones aúricas. Se hallan en la capa basal de la
epidermis, aunque su número es bajo (siendo mayor en pieles glabras). Su origen es
ectodérmico, y alrededor de la 8ª-10ª semana de desarrollo las podemos hallar en piel
glabra, mientras que en la piel fina no las encontramos hasta la 12ª semana. Durante el
desarrollo, sabemos que parte de estas células vuelven a la dermis, aunque no se conoce
97
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
la razón exacta. Sus características fundamentales, como células epiteliales que son,
son:
x Citoplasma claro enfrentado a terminal aferente
x Núcleo muy lobulado y en vertical
x Tonofilamentos: tienen las queratinas K8, K18 o K19
x Desmosomas y hemidesmosomas
x Gránulos con membrana con un halo periférico claro y un centro denso. Son más
frecuentes en la zona próxima al terminal aferente
x Contiene neurotransmisores: VIP, met-encef, bombesina…
x Prolongaciones cortas que se meten entre los queratinocitos adyacentes
Las células de Merkel son receptores táctiles que van a transmitir la información a fibras
aferentes. Su papel en el desarrollo es esencial, porque constituyen el foco de atracción
de dichas fibras. Desde el punto de vista clínico, pueden ser causa de carcinomas
neuroendocrinos
Corpúsculos de Paccini, de Vater-Paccini o de Golgi-Mazzoni
Detectan diferencias de presión. Responden a estímulos vibratorios en el rango de 20-
500 Hz. Son grandes (1mm) y se localizan en la dermis reticular, hipodermis,
articulaciones, ligamentos, etc.
Su cápsula es muy gruesa, y está constituida por capas concéntricas de elementos
conjuntivos por debajo de las cuales vamos a encontrar células de la teloglía. Una fibra
nerviosa actúa a modo de eje del conjunto, entra perpendicular. Esta fibra se ramifica
en dos.
Corpúsculos de Meissner
También son receptores táctiles que responden a estímulos vibratorios, pero en un rango
inferior, entre los 10 y los 400 Hz. Se localizan en la dermis papilar, y abundan en la
piel glabra.
Son ovalados, y su cápsula es también conjuntiva y glial, aunque de menor grosor. La
diferencia más significativa con los corpúsculos de Paccini es que la fibra nerviosa no se
sitúa a modo de eje sino en zig-zag.
98
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
FANERAS O ANEJOS CUTÁNEOS
Las faneras o anejos cutáneos son estructuras asociadas a la piel, derivadas de la
epidermis. Son las uñas, las glándulas sudoríparas ecrinas y el aparato pilosebáceo
compuesto a su vez por el folículo piloso, las glándulas sebáceas, las glándulas
sudoríparas apocrinas y el músculo erector.
Folículo piloso
Desarrollo embrionario y distribución
Los folículos pilosos son derivados epidérmicos, procedentes del ectodermo.
Comienzan a formarse a partir del tercer mes de desarrollo: se observan invaginaciones
de la epidermis, que se aprecian primeramente en cejas, barbilla y labio superior y van
apareciendo por toda la epidermis.
En el mes 5 o 6 sale el primer tallo, los primeros pelos, el lanugo, un pelo fino y
no pigmentado. Este pelo cae al nacer, excepto en las cejas, las pestañas y el cuero
cabelludo, donde es sustituido por el pelo definitivo o terminal, pigmentado y más
grueso, alcanzando la hipodermis. En el resto del cuerpo, cae el lanugo y nace el vello,
un pelo fino y no pigmentado, que llega hasta la dermis.
Al llegar a la pubertad, parte de las células donde había vello comienzan a
formar pelo terminal. Esto sucede en axilas, región ano-genital. Además en el caso de
los varones ocurre también en bigote, barba, extremidades, tronco… Aparecen por todo
el cuerpo excepto palmas, plantas, lados de los dedos, superficie lateral de los pies (bajo
el tobillo), labios, glande, prepucio, clítoris, labios menores, superficie interna de los
labios mayores.
Tenemos 2 millones de folículos al
nacer, 100.000 de los cuales se encuentran
en el cuero cabelludo. La densidad de los
folículos es variable según la zona, desde
50 folículos por cm2 hasta más de 500. La
distribución de estos folículos pilosos
presenta diferencias según el sexo debido
a la distinta herencia genética y al control
hormonal.
Tallo (pelo)
El pelo o tallo es un filamento de longitud muy variable (desde 2 mm hasta más
de 1 m.) y grosor variable. Siendo distinto según la raza. Sale de la superficie de la piel
con orientación oblicua. Está queratinizado, con una queratina más dura que en
epidermis. Con abundantes residuos de azufre, de manera que las células no se pueden
descamar (esto se debe a un alto contenido en puentes con azufre), para eliminarlo habrá
que cortar. Emerge de la raíz.
El vello es un pelo más fino, blando y corto. No tiene médula ni pigmento, y se
introduce hasta la dermis. Normalmente cubre el tronco y los miembros.
99
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
El pelo terminal es más grueso y firme, crece mucho, tiene médula y está
pigmentado. Se aloja en la hipodermis. Se encuentra en el cuero cabelludo, pestañas,
cejas, etc. Además nace en zonas muy profundas de la hipodermis.
Existen tres grandes tipos de pelo terminal según diferencias raciales: grueso
lacio, fino ondulado y crespo corto.
El color del pelo terminal es muy variable y depende de dos factores: la cantidad
de melanina y la distribución de los melanosomas:
x Pelo cano: melanocitos y melanina escasos. Además la médula empieza a
llenarse de aire, lo que hace que se vuelva más traslúcido.
x Rubio escasos: melanosomas normal melanocitos.
x Pelo rojo: por feomelanina.
En el corte trasnversal observamos:
x Cutícula: Es epitelio plano estratificado queratinizado (queratinas duras), las
queratinas se disponen como tejas; eso asegura que la cutícula selle el interior
del tallo.
x Corteza: epitelio cúbico queratinizado, con melanosomas en nº variable. Estos
melanosomas son captados desde el bulbo.
x Médula: epitelio queratinizado, las células dejan grandes espacios llenos de aire
entre sí. Con el paso de los años, hay más aire y menos células y hace que el
pelo salga cano.
100
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Raíz
Consiste en una invaginación tubular de la epidermis. Es la parte esencial para
que se forme el tallo. Lo habitual es que la raíz se vaya ensanchando conforme
avanzamos en profundidad. A partir de ella se genera el tallo, se renueva. En un corte
longitudinal observamos:
x Infundíbulo: desde la superficie hasta la desembocadura del conducto de la
glándula sebácea.
x Istmo: desde la desembocadura del conducto de la glándula sebácea hasta la
inserción del músculo erector.
x Segmento inferior: desde la inserción del músculo erector hasta el final.
x Bulbo: es el engrosamiento de la base, donde hay una entrada de conjuntivo.
El infundíbulo y el istmo son siempre permanentes, mientras que el segmento
inferior sufre cambios cíclicos. La secreción de la glándula sebácea sale por el
infundíbulo, que es por tanto vía conductora de la secreción sebácea.
Histológicamente, la raíz presenta cuatro capas, desde la parte más externa hasta
el tallo: la vaina conjuntiva, la membrana/lámina vítrea, la vaina radicular externa y la
vaina radicular interna.
Vaina Conjuntiva (raíz)
Esta vaina es un tejido conjuntivo que rodea periféricamente a la raíz. Se divide
en una capa más externa, con fibras elásticas y colágenas orientadas longitudinalmente;
y una capa interna, con fibras colágenas en disposición perpendicular. Entre ambas
capas se localiza el plexo nervioso peritriquial: fibras aferentes que recogen el cambio
de orientación del tallo (movimiento).
La membrana vítrea es la lámina basal que separa esta vaina conjuntiva de las
siguientes capas de carácter epitelial.
Vaina radicular externa (raíz)
Es un epitelio que se halla en continuidad con la epidermis. Las células están
queratinzadas con queratina blanda (no dura, como la del tallo).
En la zona del istmo se estrecha y pierde los estratos córneo y granuloso.
Profundiza, se adelgaza y se continúa con una región que se denomina matriz. En la
zona del infundíbulo, esta vaina no está en contacto con el tallo, sino que media una
separación. En el istmo y el segmento inferior, encontramos la vaina radicular interna a
continuación de la externa. Eso es por debajo, y aunque por arriba deberíamos encontrar
que se encuentra conectado con el tallo, está separada de él.
En la zona final del istmo, forma un engrosamiento que recibe el nombre de
rodete. Contiene células madre que tienen 3 destinos:
1. Ascender hacia epidermis para ir a la capa basal.
2. Descender hacia el bubo y meterse en la matriz.
3. Irse por el conducto de la glándula sebácea y repoblar las glándulas sebáceas.
101
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Vaina radicular interna (raíz)
Es un epitelio que no está en continuidad con epidermis, sino que deriva de la
matriz y se encuentra rodeando el tallo en el istmo y el segmento inferior. La matriz
genera muchas células y hace que la vaina se desplace hacia arriba. Por eso la vaina
interna se relaciona de esa manera con el tallo y el segmento inferior. Por encima de
éstos, se desintegra y se pierde. Sus células presentan queratina blanda. Esta vaina
también se descama.
Histológicamente, y en un corte transversal, se distinguen en la vaina tres capas,
de más externa a más interna:
x Capa de Henle: epitelio simple cúbico bajo o plano.
x Capa de Huxley: epitelio estratificado plano. Presenta gránulos eosinófilos de
tricohialina.
x Cutícula de la vaina radicular interna: La más interna. Es epitelio estratificado
plano queratinizado, en contacto con la cutícula del tallo. Se asegura que el tallo
esté bien emitido en la zona profunda, en la raíz del pelo.
Las capas se fusionan en el infundíbulo y acaban descamándose como una masa
eosinófila que se describe por encima del istmo.
Bulbo
En él se ubica la matriz: una capa
basal de células epiteliales en mitosis
constante. La matriz se asienta sobre una
papila dérmica de conjuntivo que se
introduce en el bulbo y depende de los
fibroblastos de la papila.
Las células epiteliales de la matriz
proceden de las células madre que emigran
en el rodete, sobre la papila dérmica. Entre
las células madre encontramos melanocitos
con gran cantidad de dendritas.
La papila emite una serie de
sustancias que estimulan la mitosis en la
matriz. Esta mitosis se produce por influencia
de fibroblastos de la papila. Si ésta se daña,
la matriz queda inerte, no se divide y eso
afecta al crecimiento del tallo.
Las células de la matriz originan la vaina radicular interna y el tallo, pero no lo
hacen de manera homogénea: las células de la zona central originan el tallo y las más
periféricas (de la capa basal que recubre la papila) originan la vaina radicular interna. La
capa basal que recubre la papila da la vaina radicular interna
102
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Cuando las células de la matriz
comienzan a ascender, pasan a una zona de
transición, la zona queratógena (entre la
matriz y las células que ya muestran queratina
dura, que solo está en el tallo), donde se
produce la transferencia de los melanosomas a
las futuras células del tallo.
Esta zona queratógena es una zona de
transición entre las células de la matriz y las
que formarán el tallo pero se empezaran a
sintetizar queratinas duras. Los melanocitos trasnfieren melanina a las células que
formarán el tallo, y así éste tendrá color.
Como la matriz se divide contantemente, asegura que las células que suban y
empujen al tallo, haciendo que se alargue y crezca. El crecimiento no es continuo, está
sometido a un ciclo.
Crecimiento del tallo
El crecimiento del pelo es dependiente de la matriz, de modo que el tallo crece
en longitud, paro lo cual la vaina radicular desaparece a la altura del infundíbulo. Es un
crecimiento cíclico, con tres fases:
1. Fase anagénica (hasta 8 años):
Las células de la matriz se dividen, haciendo el tallo cada vez más largo. La
vaina radicular interna crece, pero se descama. En las mujeres el pelo que se
origina puede llegar a los 2m, mientras que en el hombre no tanto. El segmento
inferior es muy profundo y alcanza la hipodermis. Al cabo de los años el folículo
entra en involución (fase catagénica). La papila dérmica muy bien formada y la
matriz es muy activa. Aquí el pelo tiene todas las capas.
2. Fase catagénica (2 ó 3 semanas):
Se produce un cese en el crecimiento y el folículo involuciona. Se observa una
retracción del segmento inferior, atrofia del bulbo y reducción drástica de la
actividad mitótica en la matriz y de la síntesis de melanina en melanocitos (deja
de haber). El tallo no va a crecer y se desarrolla más la vaina radicular interna.
Después hay un período de reposo en la que de esa raíz sólo queda el
infundíbulo y el istmo. El segmento inferior se ha emitido tanto que está a
continuación del istmo. No hay matriz, ni mitosis, melanina, proliferación o
queratinización.
3. Fase telogénica (3 ó 4 meses):
El segmento inferior es muy corto, pues se ha emitido tanto que está a
continuación del istmo y no se observa apenas. El tallo persiste y queda sólo el
infundíbulo y el istmo. No hay proliferación ni queratinización. El cambio lo
sufre la raíz, siendo el pelo fino y que no crece. Acaba con la activación de las
células madre del rodete que migran hacia abajo y forman una nueva matriz.
4. Nueva fase anagénica:
103
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Se inicia un nuevo periodo de crecimiento por activación de las células madre
del rodete de la vaina radicular externa. Las células madre migran y crece de
nuevo el segmento inferior, se forma un nuevo bulbo, una nueva papila dérmica
y entonces se forma una matriz donde las células se dividen activamente. En esta
nueva fase anagénica se forma un nuevo tallo y hace que caiga el viejo. Todo
esto hace que volvamos a tener un folículo con la raíz completa.
La duración de las fases varía según las zonas. Es un crecimiento asincrónico
(no en todos los folículos a la vez), con patrón en mosaico (sincronización por zonas).
En un momento dado, el 90% de folículos está en fase anagénica.
Clínica
La alopecia se produce cuando el folículo deja de formar pelo terminal y forma
vello. Los folículos presentan una fase telogénica muy prolongada. La alopecia se debe
a factores genéticos (se transmite de forma dominante en varones y recesiva en mujeres)
y endocrinos, estimulada por hormonas sexuales masculinas e inhibida por las
femeninas.
Glándula sebácea
Se forma de células madre de la vaina radicular
externa (mismo origen que la epidermis) y se asocian casi
siempre a un folículo piloso. Pueden estar a ambos lados
del folículo, pero suelen encontrarse en el ángulo obtuso
del tallo. Los folículos van oblicuos.
Son particularmente abundantes en la piel
seborreica. Drenan por un conducto que drena al
infundíbulo. Puede haber más de una glándula por folículo.
En algunas zonas no se asocian a folículos, como en labios,
labios menores, pliegues internos del prepucio y la glándula
de Meibomio. En estas zonas drenan directamente a la piel.
104
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Presentan células pequeñas basales aplanadas, ricas en orgánulos, de activa
división. Posteriormente ascienden y se cargan de la síntesis de lípidos, por lo que
desarrollan REL y crestas tubulares. Finalmente entran en apoptosis, es decir, siguen un
mecanismo de secreción holocrina con los restos celulares. La célula tarda 8 días en
realizar este ciclo y morir.
La secreción de estas glándulas es el sebo y presentan una secreción continua
(producción en 8 días). El sebo es conducido por un conducto excretor de epitelio
estratificado plano, en continuación con la vaina radicular externa saliendo por el
infundíbulo. El sebo es lubricante, emoliente (algo que reblandece), protege frente a las
temperaturas extremas y tiene propiedades bacteriostáticas.
La secreción de sebo aumenta con la pubertad, pues las hormonas estimulan más
las glándulas sebáceas. Particularmente lo hacen más las masculinas que las femeninas
(estas últimas inhiben).
Clínica
El acné es una patología que se aprecia en cara, cuello, pecho y espalda. Se
produce porque los trigliceroles son transformados por bacterias epidérmicas en ácidos
grasos irritantes. Además, se retiene sebo en el istmo debido a una secreción excesiva y
a una dificultad en el drenaje. De forma que el acné presenta además infiltración de
linfocitos en la epidermis, llegando al istmo y dando lugar a una reacción lÑinfocitaria.
105
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Músculo Horripilador
El músculo horripilador o erector del pelo es un haz de miocitos lisos. Tienen
dos puntos de inserción: uno está en la capa papilar de la dermis (zona superior de la
dermis) y el otro extremo se inserta en la vaina perifolicular (es conjuntiva), a nivel del
istmo. Cuando los miocitos se contraen,la dermis se hunde hacia abajo, se acorta el haz
y se erecta el tallo, el folículo (erizamiento). Además, cuando se contrae se abre más el
infundíbulo, facilitando la salida del sebo. La contracción está regulada por el sistema
nervioso autónomo, respondiendo a estímulos tanto emocionales como térmicos.
Glándula sudorípara apocrina
A partir del brote epidémico del folículo piloso. Se encuentra en la piel apocrina:
aréola, axila, púbica y perianal. Tienen gran tamaño y son tubulares enrolladas, a veces
ramificadas. Su conducto excretor drena por encima del conducto de las glándulas
sebáceas.
Las glándulas sudoríparas apocrinas contienen ácinos secretores de mayor
tamaño que las ecrinas. La porción secretora se localiza en la dermis e hipodermis. El
conducto excretor desemboca en el folículo piloso. Las glándulas sudoríparas apocrinas
se vuelven funcionales después de la pubertad y están inervadas por nervios
adrenérgicos.
Dos ejemplos de glándulas apocrinas especiales son las glándulas ceruminosas
del conducto auditivo externo y las glándulas de Moll del margen parpebral.
x Las glándulas ceruminosas producen cerumen, un lípido pigmentado; el
conducto excretor se abre, junto con los conductos de las glándulas sebáceas, en
los folículos pilosos del conducto auditivo externo.
x El conducto excretor de las glándulas de Moll desemboca en la superficie libre
de la epidermis del párpado o en las pestañas.
Los adenómoros tienen una luz muy amplia tapizada por epitelio simple cúbico-
plano, con células epiteliales y mioepiteliales. Las primeras son las que segregan y
106
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
almacenan el sudor (secreción lechosa, viscosa y rica en proteínas, con azúcares,
lípridos, pigmentos, lipofucsina, hierro y amonio). Es incolora, pero la descomposición
de bacterias produce olor.
Las células epiteliales son cúbicas o planas, con citoplasma eosinófilo abundante
en lipofucsina y lisosomas que parece perder la poción apical (glándula apocrina), pero
en realidad su secreción es merocrina, de composición variable según la localización.
Las células mioepiteliales se encargan de la expulsión del sudor.
El conducto excretor tiene luz estrecha y trayecto recto. Es un epitelio
estratificado cúbico, sin reabsorción, que abre al infundíbulo.
El sudor contiene bacteriostáticos y barteriocidas. Es transformado por bacterias
en sustancias odoríferas. Además, actúa como feromonas masculinas y femeninas que
cambian con el momento del ciclo menstrual y con los sexos.
Reciben estímulos emocionales y sensitivos, con control adrenérgico, y no
responden ante el calor.
Glándulas sudorípara ecrina
107
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Se encuentran por todo el cuerpo, excepto en los labios genitales externos. Son
muy abundantes en la piel gruesa. Están activas desde el nacimiento. Son
independientes del folículo piloso. Son glándulas muy pequeñas de tipo tubular simple
enrollada con fondo ciego, y drenan por el segmento canalicular.
Sus adenómeros se encuentran en la dermis profunda o hipodermis, con epitelio
psudoestratificado. Hay células mioepiteliales, claras y oscuras, que secretan el sudor,
una solución hipotónica clara con cloro, sodio, potasio, amonio, urea, lactato y pobre en
proteínas. Es considerado un órgano excretor, pues contribuye a la pérdida electrolitos
(K+, Mg2+) y la deshidratación
El segmento canalicular es más eosinófilo. Sus células se caracterizan por:
x Las células claras son bajas, con escaso contacto con la luz. Poseen pliegues
apicales, laterales y basales, abundantes mitocondrias y REL. Secretan
glucógeno (PAS+). Secretan componente acuoso por las membranas laterales.
Entre las células oscuras forman canalículos o canales secretores hacia la luz.
x Las células oscuras se localizan encima de estas células claras y secretan
glucoproteínas, además de ejercer un efecto bacterióstatico. Son basófilos por su
abundante RER y aparato de Golgi. Contienen gránulos de secreción.
x Las células mioepiteliales se localizan entre la lámina basal y las células claras.
Tienen filamentos actina y se ocupan de la expulsión rápida del sudor.
El canalículo empieza a ahuecarse y permite la salida del sudor ecrino, por lo
que no hace falta conducto. El segmento canalicular está recubierto por un epitelio
estratificado cúbico, con enrollamiento menos tortuoso y luz menor que el adenómero.
En él encontramos:
x Células basales con núcleo redondo, nucléolo y mitocondrias.
x Células luminales con tonofilamentos y citoplasma vítreo.
Aquí se reabsorben de forma parcial NaCl y agua bajo la influencia de la
aldosterona. La reabsorción de NaCl por el conducto excretor es deficiente en pacientes
con fibrosis quística y el sudor isotónico se hace hipotónico. El conducto sigue un
trayecto helicoidal cuando se aproxima a la epidermis y se abre en su superficie a través
del poro sudoríparo. Dentro de la epidermis, el conducto excretor pierde su
revestimiento epitelial y se rodea de queratinocitos.
La sudoración es termorreguladora
x Evaporación del sudor: disminución de temperatura
x Frente y cuero cabelludo, cara y resto del cuerpo, palmas y planta se van
activando por este orden
x Control colinérgico
x Mayor perfusión cutánea.
x Van a producir una secreción protectora frente al sobrecalentamiento
También hay una sudoración emocional, principalmente en palmas, plantas y
axilas. En este caso el control es adenérgico.
Cínica
x Uremia: cristales de urea (visible en labio superior): escarcha ureica
108
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Fibrosis quística: Canal CFTR defectuoso, sudor salado
Uñas
Placas córneas curvas en la cara dorsal de falanges terminales de dedos de pies y
manos. El lecho ungueal de la epidermis tiene estratos basal y espinoso. Las papilas
dérmicas son grandes y muy vascularizadas, lo que le aporta un color rosáceo.
Son visibles el surco ungueal y el rodete ungueal. La placa ungueal es
transparente, con escamas córneas muy apretadas (transparente) que contienen queratina
dura, por lo que no se descama.
x Raíz: Zona de la placa córnea debajo de la piel, con nuevas células que se van a
queratinizar, procedentes de la matriz. Se asegura un crecimiento continuo, por
lo que la placa ungueal se desliza, pero el lecho no contribuye.
x Lúnula: Semiluna blanca, con células parcialmente queratinizadas.
x Eponiquio (cutícula): Zona de piel que cubre la raíz. Si se pierde favorece la
inflamación e infección de la matriz (distrofias de la placa ungueal).
x La parte distal crece sobre hiponiquio, que asegura la permeabilidad del lecho
unqueal. Protege de bacterias y hongos. Si se daña puede producirse
onicomicosis.
Su función es protectora. Su ritmo de crecimiento variable, siendo mayor en
infancia, en las manos y en los dedos índices.
Se pueden deteriorar o dañar por enfermedades, mala alimentación,
inmovilización y desequilibrios endocrinos.
109
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
MAMA
Son órganos exclusivos de los mamíferos diseñados para nutrir a las crías, muy
inmaduras. Son glándulas cutáneas modificadas
DESARRROLLO EMBRIONARIO
El lactógeno placentario y los estrógenos estimulan el desarrollo de la glándula
mamaria. En este desarrollo participan interacciones entre el epitelio y el mesénquima, y
se divide en dos fases: 1) la formación del pezón y 2) el desarrollo de la glándula
mamaria.
En la 6ª semana de vida se acumulan células epiteliales formando una depresión
(pezón invertido). Se producen engrosamientos lineales a los largo de dos líneas
mamarias, desde la axila hasta la región inguinal (cara ventral de tórax y el abdomen).
Durante el desarrollo de la glándula mamaria, una yema de células epiteliales
ectodérmicas, la yema o botón epidérmico primario, entra en el mesodermo subyacente
En el primer trimestre se ramifican, dando lugar a 15-20 cordonas epiteliales
macizos (botones secundarios). En el segundo trimestre se ahuecan. En el tercer
trimestre aparecen los alveolos y empieza a denominarse conducto galactóferos.
110
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
El mesénquima va a inducir la proliferación de las células epiteliales, y los
adipocitos van a inducir su ramificación. Las células epiteliales son precursoras de las
células mioepiteliales, que van a migrar a la periferia. El mesodermo se diferencia en
respuesta al receptor de paratohormona (PTH-RC), segregada por el botón epidérmico
Si hay una mutación en el gen PTH se para el desarrollo del botón epidérmico.
El mesodermo se diferencia en un estroma conjuntivo y adiposo, además de en
el tejido muscular liso del pezón.
x Cada botón primario origina una mama
x Cada botón secundario origina un lóbulo mamario
En los varones los sistemas de conductos involucionan por la testosterona, pero
puede producirse ginecomastica: desarrollo pronunciado de mamas en el varón por
proliferación y ramificación de conductos y por aumento del tejido fibrocartilagenoso
periductal. Se debe a falta de Rc de testosterona (síndrome de sensibilidad androgénica),
en caso de desequilibrio entre andrógenos testiculares y estrógenos de la costeza
suprarrenal (cirrosis hepática, tratamiento de cáncer de próstata) o por síndrome de
Klinefelter (47, XXY).
Hay una involución a excepción a nivel pectoral (2), aunque puede haber
polimastia o politelia, que se extirpa quirúrgicamente. Tras el nacimiento, el pezón
protruye y la aréola se eleva según se desarrollen glándulas areolares
En el nacimiento el tamaño va a ser mayor que en la infancia, y puede haber
secreción (leche de brujas: α-lactoalbúmina, grasa, leucocitos) debido al efecto de los
estrógenos placentarios.
Tras el nacimiento involuciona el parénquima. En el varón hay desarrollo escaso
y en la mujer, desarrollo bajo influencias hormonales
En la pubertad los estrógenos en presencia de prolactina estimulan el desarrollo
de conductos e hiperplasia de adiposo. Hay cambios cíclicos por progesterona, que
111
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
estimula la formación de nuevas yemas alveolares: Las células epiteliales que revisten
los conductos galactóforos contienen receptores estrogénicos citosólicos y nucleares; la
progesterona estimula la formación de nuevas yemas alveolares, que sustituyen a las
yemas antiguas que involucionan y que acaban desapareciendo al final del ciclo ovárico.
Estos cambios cíclicos se producen en cada ciclo menstrual. El desarrollo máximo solo
se alcanza en gestación y lactancia.
ESTRUCTURA
Papila y aréola mamarias
Tiene epidermis arrugada, con epitelio estratificado plano queratinizado. Sus
papilas dérmicas son muy profundas por la unión dermoepidérmica marcada, de una
tonalidad rosa.
En la pubertad y gestación el pezón es promienente y aumenta la pigmentación
(melanocitos estimulados por estrógenos).
Hay una rica vascularización sanguínea y linfática. Los miocitos forman haces
musculares lisos longitudinales y radiales en la dermis profunda. La inervación sensitiva
es rica, permitiendo la erección de la papila estimulada por la prolactina y la oxitocina.
En la papila mamaria hay 15-20 orificios en disposición anular. Se continúan
con 15-20 conductos galactóforos, que drenan un lóbulo mamario cada uno. En una
mama inactiva, los orificios están tapados por queratina.
En la aréola mamaria hay glándulas sebáceas no asociadas a folíclos pilosos,
sudoríparas y areolares o de Montgomery (intermedias entre las dos anteriores). Estas
glándulas de Montgomery sobresales y su secreción es grasa, lubricante y protectora de
la aréola en la lactancia.
Lóbulos mamarios
Cada botón primario origina una mama, y cada botón secundario origina un
lóbulo mamario. Por tanto, una mama contiene entre 15 y 20 lóbulos mamarios.
Los lóbulos mamarios tienen una disposición radial desde el pezón, y están
separados por tabiques de conjuntivo denso denominado conjuntivo interlobular, que
contiene abundantes adipocitos. Además, entre los lóbulos hay bandas de conjuntivo
fibroso, algunas de las cuales unen los lóbulos a la dermis insertándose en la fascia que
cubre el músculo pectoral, denominándose ligamentos suspensorios o de Cooper.
Después de la gestación y el periodo de lactancia, estos ligamentos no recuperan su
longitud inicial, ya que son fibrosos, no elásticos, lo cual hace que la mama se acabe
cayendo en la mujer.
112
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
En cada lóbulo hallamos también una glándula tubuloalveolar ramificada, que es
en realidad una glándula sudorípara modificada. Por tanto, si en cada lóbulo hay una
glándula y en cada mama 15-20 lóbulos, tenemos que cada mama dispone de 15-20 de
estas glándulas independientes.
Lobulillos mamarios
Los lóbulos se subdividen en numerosos lobulillos separados por tejido
conjuntivo denso interlobulillar. Este tejido interlobulillar se entrecruza con el
interlobar, y por ello tienen características similares. Resulta clave conocer esto cuando
queremos realizar una extirpación quirúrgica, pues esta habrá de realizarse a nivel del
lóbulo.
MAMA A LO LARGO DE LA VIDA
Mama inactiva
En la mama inactiva distinguimos un estroma y un parénquima.
Estroma
Es muy abundante, y está formado por tejido conjuntivo. Comprende tanto el
conjuntivo interlobar como conjuntivo inter e intralobulillar.
Este último no se ha mencionado antes: se trata de tejido conjuntivo laxo (los
otros dos son densos) que carece de adipocitos, y en el cual encontramos escasas células
libres (linfocitos, mastocitos y macrófagos, más adelante se verá su importancia). Es
muy rico en capilares, lo cual constituye una dificultad ante la diseminación tumoral a
este nivel.
Parénquima
Está constituido por el sistema excretor, que en la mama inactiva es escaso. Las
yemas alveolares dependen de progesterona, sin embargo, el sistema de vías excretoras
depende de estrógenos (MUY IMPORTANTE).
Este sistema de vías excretoras dependiente de estrógenos tiene una constitución
particular. El primero de los elementos de estas vías es el conducto galactóforo, que
desemboca en una porción dilatada denominada seno lactífero. El seno lactífero a su vez
origina ramificaciones dicotómicas que provocan la existencia de un conducto terminal
por lobulillo, el cual se vuelve a ramificar (esta vez de forma irregular, no dicotómica)
para originar conductillos intralobulillares, separados por conjuntivo intralobulillar. Este
sistema de ramificaciones del parénquima se conoce también con el nombre de sistema
ductal.
113
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
1. Conducto galactóforo: presenta un epitelio estratificado cúbico, que si se abre a
un orificio de luz estrecha se convierte en estratificado plano. Tiene un diámetro
de unos 2-4 mm.
2. Seno lactífero: como ya se ha dicho, esta porción se halla dilatada. El epitelio
sigue siendo estratificado cúbico, y es aquí donde se acumula la leche,
constituyendo por tanto un reservorio.
El epitelio estratificado cúbico se va convirtiendo a medida que avanzan las
ramificaciones del sistema ductal en simple prismático, y tras ello la altura de las células
disminuye y encontraremos epitelio simple prismático rodeando los conductos, y por
fuera de ellos células mioepiteliales con capacidad contráctil.
3. Conducto terminal: cada lobulillo drena en uno de estos conductos
4. Conductos intralobulillares: formados por ramificaciones del conducto terminal.
Presentan un fondo ciego.
En la mama inactiva, no encontramos unidades secretoras (alveolos), el sistema
acaba con el ciego de los conductos intralobulillares.
Ciclo menstrual
A pesar de que la mama esté inactiva, se pueden producir mínimas
modificaciones debido a los cambios hormonales originados por el ciclo menstrual.
Hacia el día 16 del ciclo, el cuerpo lúteo induce un aumento de progesterona y
estrógenos.
x Con la estimulación estrogénica, la altura de las células epiteliales de los
conductillos intralobulillares aumenta, y podemos hallar signos de secreción. En
los conductos terminales se abren luces mínimas conforme se acumula dicha
secreción, y en el conjuntivo intralobulillar se acumula líquido y
glucosaminoglucanos.
x Con la estimulación de la progesterona se desarrollan nuevas yemas alveolares y
las antiguas involucionan
Estos cambios ocasionan la hipertrofia lobulillar, y pueden llegar a provocar un
ligero aumento del tamaño de la mama y molestias.
Si no hay fecundación, los niveles de progesterona caerán y la estructura de la
mama se mantendrá como se ha descrito en los apartados de parénquima y estroma. Si
hay fecundación, los altos niveles de progesterona promoverán el desarrollo de alveolos
o unidades secretoras.
114
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Mama en el embarazo
En el embarazo, en preparación para la lactancia, tienen lugar en la mama una
serie de cambios que llevan al desarrollo de los extremos de los conductos, antes ciegos.
Estos cambios se hallan estimulados por hormonas placentarias (lactógeno) y maternas
(progesterona, estrógenos y prolactina). Entre ellos destacan:
x Disminución del tejido conjuntivo y adiposo que constituye el estroma
x Aumento del parénquima
o Ramificación de los conductos
o Aparición de alveolos
x Infiltración de linfocitos y eosinófilos
115
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Primer trimestre
Durante el primer trimestre aumenta la vascularización del pezón y la areola
circundante, con lo que también lo hace su pigmentación.
Las células epiteliales de los conductillos terminales sufren una proliferación
hiperplásica, y en su citoplasma aparecen vacuolas.
Segundo trimestre
Las células epiteliales se vuelven secretoras, y al igual que ocurría en el ciclo
menstrual esto causa la apertura de las luces de los conductos. El tejido conjuntivo
intralobulillar se hipertrofia, y aumenta el número de células inflamatorias.
Tercer trimestre
La total conversión de las células epiteliales en secretoras hace que una cantidad
copiosa de secreción se acumule en los conductillos terminales, cuyas luces son por ello
muy amplias. La hiperplasia de las células epiteliales secretoras llega a su máximo
nivel, y se forman los alveolos.
Cambios celulares en la mama durante el embarazo
1. Las células se vuelven eosinófilas
2. Aparecen abundantes RER y AG supranuclear, así como mitocondrias, lo cual
responde a la nueva función secretora, que requiere la formación de vesículas y
energía para ello.
3. Lisosomas densos.
4. Aparecen vesículas de secreción que contienen proteínas y glucoproteínas para
la futura leche.
5. Aparecen inclusiones lipídicas que otorgarán a la leche el componente graso.
Mecanismo de secreción de la leche
En relación con las inclusiones lipídicas y las vesículas de secreción proteica se
ha hablado tradicionalmente de dos tipos de secreción:
x Merocrina, típica de los componentes proteicos
x Apocrina, típica de los componentes grasos. Las inclusiones lipídicas en
apariencia se fusionan formando grandes lóbulos que viajan al citoplasma apical
y parecen liberarse con una mínima cantidad de citoplasma. Posteriormente, se
ha visto que no se liberan con citoplasma, con lo que la apocrinia no es tal.
Como ya sabemos, la leche materna constituye un mecanismo de inmunidad
pasiva, puesto que tiene un alto contenido en Igs. Estas son sintetizadas por las células
plasmáticas que se infiltran en el segundo trimestre del embarazo en el conjuntivo
intralobulillar. Por un mecanismo de transcitosis consiguen pasar y son segregadas a la
luz alveolar.
116
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Mama en la lactancia
Tras el parto, se produce una caída súbita
de progesterona y estrógenos, y va a entrar en
acción la prolactina, que tiene efecto
lactogénico, y tarda tan solo unas horas o unos
días en estimular la secreción. Aun así, la
producción de leche requiere también la
intervención de glucocorticoides suprarrenales,
PTH y GH. Estamos hablando de producción,
esto es, de secreción. La excreción de la leche
depende de la oxitocina, que se encarga de
estimular a las células mioepiteliales existentes
en los alveolos (que son 20 veces más sensibles
a ella que el miometrio uterino) provocando la
eyección de la leche.
117
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Con fines secretores, durante la
lactancia se produce un desarrollo
máximo de los alveolos formados en el
último trimestre de gestación. Estos
alveolos están constituidos por un
epitelio cúbico simple rodeado de
células mioepiteliales que facilitan la
secreción, que los primeros días tras el
parto es el calostro y después será leche
propiamente dicha.
Incluso en un mismo lobulillo mamario, los alveolos se hallan en distintos
estados funcionales: podemos hallar alveolos cuya luz está dilatada y llena de leche, que
disponen de un epitelio cúbico más bajo; y otros cuya luz es estrecha debido a que
apenas tienen leche y que poseen un epitelio más alto.
Calostro
Como ya se ha avanzado, el calostro es la secreción láctea propia de los primeros
días posteriores al parto. Se trata de una secreción amarillenta alcalina que contiene más
proteínas, vitamina A, Na+ Y Cl- y menos lípidos, carbohidratos y K+ que la leche
definitiva. También contiene una cantidad considerable de anticuerpos, sobretodo IgA,
que otorgan inmunidad pasiva al recién nacido. De ahí que el destete precoz y
alimentación con biberón priven al bebé de
Leche madura
Es un alimento específico de cada especie mamífera. En un embarazo normal, la
mujer puede secretar hasta 1,1 L de leche al día, más de 2 L incluso si se trata de un
parto múltiple. Tiene componentes nutritivos, inmunológicos, protectores y
estimuladores del crecimiento; entre los que destacan:
x Proteínas: caseína, alfa-lactalbúmina, PTH-RP, lactoferrina, lisozima
x Lípidos: triglicéridos, colesterol
x Azúcares: lactosa, atrae agua hacia las vesículas secretoras haciendo la leche
líquida
x Igs: Ig A dimérica. De ahí que el destete precoz y alimentación con biberón
priven al bebé de inmunidad pasiva. Incluso después de 5 meses de lactancia, la
cantidad de Ig A aportada al bebé vía leche materna puede llegar a los 0,5 g
diarios.
Ciclo secretor de la mama en la lactancia
Inicio del ciclo secretor
Al inicio del ciclo secretor, la situación es la siguiente:
118
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Alveolos pequeños y con la luz vacía, cuyas células epiteliales son más altas,
prismáticas en vez de cúbicas. El epitelio, además de ser prismático, se
encuentra además plegado.
x Células epiteliales con núcleo basal y citoplasma eosinófilo
Progresión del ciclo secretor
x Alveolos crecen, su luz se llena de un material granulado eosinófilo con gotas
lipídicas. Las células epiteliales se hacen cúbicas bajas y el epitelio deja de estar
plegado
x Células contienen RER, AG, mitocondrias y grandes vacuolas vacías.
Regulación hormonal del ciclo secretor
x Secreción inicial: está regulada por la prolactina, PTH, GH y glucocorticoides.
x Expulsión: regulada por la oxitocina
Los niveles de prolactina caen tras el parto. Sin embargo, resulta imprescindible
amamantar al bebé, con lo que la producción de prolactina se halla estimulada por el
bebé. La succión por su parte estimula receptores sensitivos en la papila mamaria. Los
impulsos sensitivos llegan al hipotálamo, desde donde se inhibe la liberación del factor
inhibidor de la prolactina, logrando que la adenohipófisis libere prolactina en una
cantidad 10 veces mayor, logrando una elevación temporal de los niveles de esta
hormona. Otros efectos de la prolactina son:
1. Inhibición de la hormona liberadora de gonadotropina y de LH
2. Amenorrea de la lactancia: en un 50% de los casos, no hay ovulación, lo cual
constituye el principal método anticonceptivo en culturas en las que se
amamanta al bebé durante dos o tres años.
A su vez, la neurohipófisis libera oxitocina, que actúa sobre las células
mioepiteliales asegurando la expulsión de la leche, lo que resulta imprescindibles para
reiniciar el ciclo.
La producción y la secreción son continuas. La leche se acumula en la luz de los
alveolos y los conductos, y su eliminación es episódica y depende de la participación
del lactante. Si el bebé mama regularmente, la lactancia puede mantenerse entre 2 y 3
años.
Si no hay succión por parte del bebé, se acumula el producto en la luz de los
alveolos y conductos, comprimiendo los vasos sanguíneos circundantes e impidiendo la
llegada de la oxitocina procedente de la neurohipófisis, con lo que la expulsión de leche
cesa. Al cabo del tiempo la secreción alveolar es reabsorbida y los alveolos colapsan, a
la par que las epiteliales degeneran por acción de vacuolas autofágicas y se sintetizan
nuevas enzimas lisosomales. Esta degeneración epitelial hace que aparezcan macrófagos
intraepiteliales, aunque este proceso no se considera una reacción inflamatoria.
119
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Así, la mama involuciona y retorna al estado de reposo: aumenta de nuevo la
cantidad de tejido conjuntivo y adiposo y los alveolos retoman su aspecto de conductos
macizos, aunque no desaparecen del todo, y los restos de su secreción pueden
permanecer en los conductos.
Involución de la mama en menopausia
La mama retorna al estado prepuberal:
x Escasas fibras colágenas
x Sin fibras elásticas
x Atrofia epitelio secretos y sistema ductal
x Pobreza celular del tejido
Clínica: trastornos mamarios
Benignos
x Mastopatía fibroquística: Los desequilibrios hormonales causan la proliferación
del tejido conjuntivo y la aparición de quistes en los conductos. Mastalgia.
x Fibroadenoma, displasia mamaria benigna: Se forman masas de tejido epitelial y
conjuntivo de crecimiento lento. Es indoloro, pero los conductos mamarios se
dilatan y aparecen nódulos.
Malignos
x Carcinomas: La mama es el lugar más frecuente de cáncer en la mujer. El 80%
de los cánceres de mama tiene su origen en el epitelio de los conductos
galactóforos. Tanto estas células como las cancerosas muestran receptores de
estrógenos. El 10% tiene su origen en el epitelio de los conductillos terminales y
en el epitelio alveolar (carcinomas intraductales). Es decir, en conjunto, el 90%
120
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
son carcinomas ductales, puesto que tienen su origen en el sistema ductal.
Además, la rica vascularización de la mama facilita las metástasis.
Los genes BRCA1 y BRCA2 codifican para proteínas supresoras de tumores. El
20-30% de los carcinomas de mama expresan estos genes. Las mujeres con
mutaciones en ellos tienen un alto riesgo de sufrir cáncer de mama y de ovario
infiltrantes, por lo que es recomendable una mastectomía total profiláctica
bilateral.
El tratamiento de sustitución con estrógenos en mujeres postmenopáusicas
también se ha relacionado como factor de riesgo en el cáncer de mama.
121
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
APARATO DIGESTIVO
INTRODUCCIÓN
El aparato digestivo es un aparato que muestra una importante diferenciación tanto
funcional como estructural. El aparato digestivo consta de las siguientes partes: cavidad
oral, faringe, esófago, estómago, intestino delgado, intestino grueso.
En la cavidad bucal se inicia el proceso de digestión. Estamos refiriéndonos a una
digestión tanto mecánica como enzimática. La digestión mecánica comienza con los
dientes que cortan y muelen los alimentos que ingerimos. Para esta función es también
muy importante humedecer los alimentos de manera que aquí van a intervenir las
secreciones glandulares, las cuales también van a aportar una serie de enzimas que
posibilitan el inicio de la digestión enzimática.
En esta función digestiva juega un papel importante la lengua que permite envolver los
alimentos y masticarlos de una manera más activa. Como consecuencia de este proceso,
los alimentos son triturados en pequeñas porciones de aproximadamente 2 cm, a las que
llamamos bolo alimenticio. Cuando esto está listo, el alimento está preparado para ser
deglutido.
El alimento se deglute vía esófago y alcanza el estómago. Esta es la parte del tubo
digestivo que se va a encargar del proceso digestivo. Para ello segrega una serie de
enzimas (así como ácido clorhídrico) y junto con los movimientos de su túnica muscular
va a poder degradar los alimentos en nutrientes.
En un momento dado, estos nutrientes van a alcanzar la primera porción del intestino
delgado, es decir, el duodeno. En esta zona observamos unas secreciones de pH básico,
es decir, secreciones alcalinas. Estas secreciones están destinadas a neutralizar el ácido
que llega del estómago y a su vez van a permitir la activación de secreciones
pancreáticas.
De tal forma que la digestión se ultima en esta porción del intestino (no termina a nivel
del estómago); y para terminar la digestión también es fundamental la llegada la
duodeno de la bilis. Esta bilis va a permitir la emulsión de los ácidos grasos, lo cual es
clave para una posterior absorción.
La absorción de los nutrientes tiene lugar sobre todo a nivel del yeyuno. Estos nutrientes
van a pasar a la sangre y son conducidos en primer lugar al hígado (la principal glándula
aneja al tubo digestivo) que se va a encargar de procesar estos nutrientes, distribuirlos,
sintetizar otros más complejos y también lleva a cabo un papel esencial de
detoxificación.
Si volvemos al tubo digestivo, aquellos elementos que están en la luz y que no se
absorben, van a comenzar a perder agua. Se reabsorbe agua ya desde el ilion, y esta
función va a terminar de cumplirse en el intestino grueso. Además, estos elementos que
están en la luz del intestino grueso se van compactando para lo cual cumple un papel
fundamental la secreción mucosa.
122
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
CAVIDAD BUCAL
Es el lugar en el que comienza el proceso de
digestión con una digestión mecánica por los
dientes que cortan, desgarran y muelen. El
proceso continúa con una humectación de las
secreciones glandulares y una enzimática por
las enzimas. Para todo esto es fundamental la
lengua para la mezcla de alimentos con las
secreciones y para formar los bolos
alimenticios.
Consta de dos zonas:
x Vestíbulo: Delimitado por los labios,
las mejillas, las encías y los dientes. Es en este espacio en el que donde vierten, a
través de conductos excretores, las secreciones de las glándulas menores
yugales, labiales y molares y una glándula salivar mayor que es la parótida.
x Cavidad oral: Se encuentra detrás de los dientes. Está limitada por los dientes,
las encías, el paladar duro y blando por arriba; por abajo el piso y la lengua y por
atrás por el istmo de las fauces. Este es el lugar en el que vierten las secreciones
de las glándulas sublinguales y las submaxilares.
Se debe mencionar que el rafe divide en dos cavidades simétricas el paladar. La cavidad
oral tiene tres zonas distintas, por lo que no es una estructura uniforme. Estas zonas son:
mucosa masticatoria, mucosa no masticatoria y mucosa especializada.
Mucosa masticatoria
Recubre las encías y paladar duro. Tiene dos únicas capas:
x Epitelio: Es plano estratificado queratinizado, siendo en algunas zonas de la
encía paraqueratinizado. Posee cuatro estratos, pero no posee estrato lúcido.
x Lámina basal propia: Se encuentra bajo el epitelio y es similar a la dermis, con
las dos zonas (papilar y reticular). En esta lámina propia tenemos papilas
dérmicas y crestas epiteliales muy profundas y como la unión entre la lámina
propia y el epitelio están muy unidas, los capilares llegan hasta arriba. Además,
presentan muchas terminaciones nerviosas libres así como corpúsculos (de
Meissner).
Esta mucosa es muy gruesa y espesa. No se estira, y no es elástica debido a que está
muy unida al hueso de debajo. Ofrece mucha resistencia a cualquier daño o traumatismo
(resistencia frente a las tracciones y/o estrés mecánico).
En el caso de la mucosa masticatoria de las encías, está adherida al diente o al hueso
alveolar, es decir, que debajo de la lámina propia no hay submucosa, yendo
directamente el diente o los tejidos a donde se sujeta el diente. Por esto aquí no tenemos
glándulas.
El epitelio de la mucosa de las encías puede ser de fijación o cervicular. El epitelio de
fijación es la parte del epitelio de la mucosa (masticatoria) que está justo pegado al
diente (es la zona que se encuentra en el extremo del surco). Está enormemente sellado,
123
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
con muchas uniones, siendo esto fundamental pues es importante para evitar que se
puedan colar agua o restos de comida, causando si no infiltración de restos y gingivitis
(inflamación de encías)
El epitelio cervicular es la parte del epitelio de la mucosa que tapiza el surco gingival,
es decir, tapiza la cavidad del alveolo dentario donde están insertos los dientes. Esta
zona no tiene epitelio queratinizado y es más fino que el del resto de la mucosa de la
encía.
También hay masticatoria en el paladar duro, el cual es la zona más anterior del techo.
Esta mucosa contiene abundantes papilas. Su epitelio progresa a epitelio respiratorio en
la zona profunda. Aquí si hay submucosa, luego la mucosa descansa en tejido
conjuntivo rico en colágeno, con haces que se entrelazan con otros haces del colágeno
del periostio.
En esta submucosa, en la parte posterior del paladar duro, hay glándulas de tipo
mucoso. En la parte anterior hay tejido conjuntivo, con fibras colágenas y muchos
adipocitos. La zona de la submucosa, no existe ni junto a las encías, ni en la zona del
rafe.
Mucosa No Masticatoria
Se encuentra en labios, mejillas, piso de la boca, cara ventral de la lengua, mucosa
alveolar y paladar blando. Hay dos estratos: epitelio y lámina propia. Es una mucosa
con menos capas (solo la basal, espinosa y superficial) y más fina (más permeable) que
la masticatoria. Es un epitelio estratificado no queratinizado con solo tres estratos y que,
en algunos puntos, como en el borde libre del labio, se puede queratinizar.
En algunas zonas es excesiva la permeablidad. La lámina propia de estas zonas tiene
una mayor elasticidad lo que consigue que sea una mucosa más fina y que se adapte
mejor a los movimientos de la cavidad oral. El hecho, además, de que este epitelio sea
más fino, permite la difusión de fármacos por la existencia de zonas más permeables (se
absorben mejor) como el suelo de la boca. Por ejemplo las pastillas que tomamos se
deben poner debajo de la lengua.
En los labios, histológicamente, presenta tres zonas: una externa o cutánea, una interna
o mucosa oral, y otra roja o del bermellón.
x Cara externa o área cutánea: con piel y dermis y epitelio plano estratificado que
presenta folículos pilosos, glándulas sebáceas y sudoríparas.
x Cara interna o mucosa oral: Es la que presenta la mucosa no masticatoria y en su
lámina propia están contenidas las glándulas salivales labiales menores (que se
abren al vestíbulo). Bajo esta lámina propia encontramos los músculos
orbiculares de los labios.
x Área roja o bermellón: Esta zona tiene epitelio tremendamente fino donde las
células se hacen translúcidas y presentan unos gránulos eosinófilos con eleidina
(son característicos). Esta zona está firmemente anclada, con papilas muy
pronunciadas y profundas a su lámina propia, que presenta un forma irregular y
papilas muy vascularizadas, lo que dota de su color rojo característico a los
labios.
124
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Como todos sabemos, los labios se nos secan fácilmente porque no tenemos glándulas
de ninguna clase. Además, tampoco hay folículos pilosos y puede presentar
paraqueratinización (no hay pérdida de núcleos y el citoplasma no se tiñe tanto).
Las mejillas poseen mucosa no masticatoria que se asienta sobre una submucosa muy
desarrollada, fibrosa (con mucho colágeno y fibras elásticas para que cuando movamos
la boca no nos mordamos los carrillos). Aquí tenemos glándulas salivales yugales y
molares y además tenemos glándulas sebáceas bastante grandes no asociadas a folículos
pilosos conocidas como gránulos de Fordyce que están en la parte posterior de las
mejillas y son muy abundantes. Debajo de la submucosa encontramos el músculo
buccinador.
El paladar blando conforma la parte posterior del techo de la cavidad oral y es muy
movible debido a que no está asentado sobre hueso. Termina en una proyección
posterior conocida como la úvula.
En él tenemos una mucosa no masticatoria con papilas abundantes. Se pasa de epitelio
plano estratificado no queratinizado a epitelio respiratorio en la zona nasal con
linfocitos y glándulas mixtas. Además, bajo la mucosa no masticatoria de la zona oral
tenemos una submucosa con glándulas mucosas y debajo de esta hay una capa
muscular.
Mucosa Especializada
Cubre la cara dorsal de la lengua, las porciones laterales.
LENGUA
Es un órgano muscular esquelético con fibras
musculares longitudinales, verticales, horizontales y
oblicuas. Esta gran variación en la disposición dota
al órgano de mucha flexibilidad y movilidad en la
porción anterior porque la posterior está anclada al
hueso hioides.
La musculatura se divide en:
x Extrínseca: Compuesta por miocitos que se
insertan fuera de la lengua y que permite
mover la lengua.
x Intrínseca: Compuesta por miocitos que se
insertan en la propia lengua y que permite
alterar la forma de la lengua.
La cara ventral está formada por una mucosa no masticatoria bastante fina (lámina
propia fina) y sin submucosa (es muy permeable). Mientras que la cara dorsal se
denomina mucosa especializada.
Esta cara dorsal la podemos dividir en tres tercios: Los dos tercios anteriores constan de
papilas linguales y el tercio posterior denominado raíz o base consta de amígdalas
125
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
linguales, pero no de papilas linguales. Entre ellos tenemos un surco terminal al final
del cual tenemos el agujero ciego que son los restos del conducto tirogloso.
Papilas linguales
Irregularidad de la mucosa de la cara doral de la lengua. Tiene una zona central de
tejido conjuntivo que está recubierto por un epitelio plano estratificado que puede
variar. Al conjunto de epitelio más conjuntivo se le denomina papila lingual. Hay varios
tipos:
x Filiformes: Son las más abundantes, por toda la cara anterior. Son proyecciones
en forma cónica, con un tejido conjuntivo recubierto por epitelio estratificado
MUY queratinizado. Este epitelio con forma cónica apunta hacia la raíz. Están
como alineadas. Tienen SÓLO función mecánica (ayudan a romper el alimento,
degradarlo, sin que actúen botones gustativos). Si la lengua esta blanquecina por
algún trastorno, son estas papilas las que acumulan los restos de células
escamadas.
x Fungiformes: están entremezcladas, con forma de seta u hongo. Presenta un eje
de tejido conjuntivo MUY vascularizado (por eso se ven como puntos rojos).
Rodeándolo hay un epitelio plano estratificado NO queratinizado. Estas sí tienen
botones gustativos y se encuentran en las caras laterales. Muy numerosas,
sobresaliendo entre las hileras de papilas filiformes.
x Foliadas: muy abundantes en algunas especies (en la nuestra no, en el adulto
menos que en el niño) y se sitúan en las zonas laterales de la cara dorsal de la
lengua. Tienen un eje conjuntivo cubierto por epitelio estratificado no
queratinizado con botones gustativos que se puede ramificar, dando lugar a
crestas alineadas y perpendiculares al eje principal. Estas papilas foliadas están
separadas por hendiduras o surcos profundos en cuyo fondo está el drenaje de
las glándulas serosas.
x Caliciformes: (o circunvaladas). Son las más grandes de todas, son de tipo
gustativo y tienen abundantes botones. De aspecto redondeado, muy globoso.
No son elevaciones de la mucosa sin más, donde hay una papilas hay una
depresión. Al fondo de la depresión es donde se alza la papila caliciforme. En
esa emisión del fondo se abren los conductos excretores de las glándulas de von
Ebner (de tipo seroso y limpia la zona deprimida para que no queden estos). No
tenemos muchas (8-12 papilas) y están justo en paralelo al surco terminal
(alineadas justo por delante de él).
El surco que separa la papila del resto de la lengua (entre la papila y la depresión) es
bastante profundo y amplio, llamándose a veces valle/vallum y en él desembocan los
conductos de glándulas serosas de Von Ebner limpiándolo.
El epitelio no tiene una queratinización marcada pero es más grueso que el de las
foliadas y el de las fungiformes
126
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Botones gustativos
Estructuras ovaladas, redondeadas, pálidas a microscopia óptica con una tinción
convencional, que ocupan todo el grosor del epitelio en el que se encuentran. Tenemos
unos 3000 botones que están en las papilas fungiformes, foliadas y caliciformes además
de en la lengua (arco glosopalatino, paladar blando, parte superior posterior de la
epiglotis y parte posterior de la faringe).
En ellas distinguimos a M.O la fosita, poro gustativo (zona más hundida) y tres tipos de
células:
x Basales: Pequeñas que no alcanzan la luz.
x Oscuras periféricas o de sostén: Altas, muy heterocromáticas con
heterocromatina densa que se disponen en la periferia.
x Claras sensoriales, neuroepiteliales o gustativas: Altas, con eucromatina clara,
que se disponen en el centro del botón.
127
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
En la zona apical podemos observar un material eosinófilos con unos filamentos
denominados pelos gustativos.
A M.E.T, en cambio, en un poro gustativo encontramos cuatro tipos de células:
x Tipo IV (llamadas antes basales):
encargadas de la regeneración del botón,
formando células de sostén o gustativas.
x Tipo I (oscuras): de sostén periférico, con
capacidad también secretora teniendo
gránulos densos en el citoplasma que
secretan a la fosita por exocitosis el
material amorfo que es eosinófilo.
Presentan abundante RER, mitocondrias y
aparato de Golgi, así como
microvellosidades.
x Células II (claras): Con función de soporte
pero sin gránulos. Se cree que son de tipo
I pero que ya no segregan (en un estado
funcional distinto). Tienen
microvellosidades y REL.
x Células III (claras): Corresponden a otra parte de las células clara que serían las
sensitivas o gustativas, es decir, son neuronas con una prolongación apical que
invade el material amorfo y que terminan en una expansión o maza que
sobresale esa matriz amorfa. En cambio, en su base, se aprecian fibras nerviosas
de naturaleza sensitiva en proximidad a ellas (con lo cual hacen sinapsis en la
zona basas).
Las células se encuentran muy estrechamente unidas ente sí, con uniones occludens,
fibras aferentes o sinaptando.
Los botones gustativos son, por tanto, órganos sensitivos dentro del epitelio de la lengua
que se encargan de percibir sabores (en las células claras vamos a encontrar vesículas
con neurotransmisores) lo que es importante para para saber si lo ingerido está en buen
estado. Además, cuando los botones gustativos se activan van a desencadenar la
secreción de saliva y de jugo gástrico.
Estos botones están inervados por los pares craneales dependiendo de la localización:
VII (facial, que recoge información de los dos tercios anteriores), IX (glosofaríngeo que
recoge la información del tercio posterior) y X (vago, que recibe información de otras
zonas que no son la lengua).
Los botones perciben 5 sabores, y todos los botones pueden percibir todos los sabores
(no existen botones gustativos para cada sabor). Estos cinco sabores son dulce, salado,
amargo, ácido y umami. El umami es un sabor que se debe a los glutamatos, presentes
en carnes, sopas, conservas (es el sabor de los Risketos) y a los ribonucleótidos.
Para poder saborear las sustancias, éstas deben estar disueltas en agua. Sabemos que
existen diferentes receptores (TR1, TR2 y TR3) que están en la membrana apical de las
células gustativas.
128
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Cuando el receptor se une a una sustancia sápida, se altera una proteína G denominada
gustductina y va a haber diferentes vías según el tipo de sustancia percibida. En
cualquier caso se desencadena un impulso nervioso que viaja en primer lugar al tronco
cerebral pudiendo viajar posteriormente a la corteza (siendo conscientes del sabor). Los
sabores que mejor se detectan son el salado y el ácido, en los cuales el mecanismo se
produce por canales.
El sabor salado provoca apertura de los canales de Na+ que pueden bloquearse con
diuréticos pero no siempre.
El sabor ácido está relacionado con los protones produciéndose un bloqueo de los
canales de K+. En otras especies parece que los protones abren canales de sodio y son
capaces de atravesarlos.
El sabor dulce es más complejo porque no está ligado a un ion concreto sino que hay
muchas moléculas, desde grandes a pequeñas (como la glucosa, la sacarina o el
aspartamo). El receptor del sabor dulce está ligado a una proteína G que conduce a un
aumento del AMPc bloqueando canales de Ca2+.
El sabor amargo es más importante porque muchas de las toxinas son amargas. En este
caso los RC de las sustancias amargas actúan a través de proteínas G modificando los
niveles de fosfolipasa C, PIP, IP3 y DAG lo que produce una cascada que finalmente
causa la salida de Ca que es la misma vía que se produce con las sustancias de sabor
umami. Es decir, a diferencia del sabor dulce, estos dos sabores no bloquean los canales
de calcio sino que aumentan sus niveles porque se libera desde dentro (RE).
Clínica
La ausencia del gusto se denomina AGEUSIA y puede tener diferentes causas:
x Disautonomia familiar (síndrome de Riley-DAY): es una enfermedad
hereditaria, que presenta ausencia de papilas fungiformes y botones gustativos,
lo que da lugar a la ageusia (no percepción de sabores).
x Impedimentos que hacen que las sustancias gustativas no interaccionen con los
botones: microorganismos, moco, líquido, trastornos en la producción de
saliva…
x Destrucción de células sensitivas por enfermedades degenerativas de la cavidad
oral o sección de los nervios sensitivos.
x Inhibición en la tasa de renovación de los botones por metales pesados o por
ciertos fármacos.
Glándulas linguales
Cerca de la punta tenemos unas glándulas mixtas llamadas de NUHN o de BLANDIN
que forman una saliva que se vierte a la cara ventral.
Otras son las glándulas de von EBNER que son serosas y se abren al surco (al fondo)
que rodea a las papilas caliciformes en la musculatura. Se encuentran en la parte
posterior de la lengua
129
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Las glándulas mucosas son abundantes en la parte posterior y son las que se abrían en
las criptas de las amígdalas linguales en la raíz.
FARINGE
La faringe constituye el cruce de las vías respiratoria y digestiva. Comunica las
cavidades oral y nasal con la laringe y esófago. Su función es actuar como cámara de
resonancia de la fonación. Aunque posee tres tramos (rinofaringe, orofaringe y
laringofaringe) al estar hablando del aparato digestivo nos centraremos en la orofaringe.
Orofaringe
Está revestida por epitelio plano estratificado no queratinizado, bajo el cual hay lámina
propia muy fibrosa, con abundantes fibras elásticas longitudinales. La mucosa se asienta
sobre la musculatura longitudinal (no tiene muscularis mucosa), aunque algunos
miocitos son más horizontales. La adventicia es fibrosa. Distalmente, aparece una
submucosa con glándulas mucosas.
Y tenemos después los músculos constrictores y luego la capa fibroelástica con
colágeno y fibras elásticas.
Amígdalas
Acumulaciones masivas de nódulos linfáticos que se sitúa alrededor del istmo de
las fauces, en la orofaringe y en la rinofaringe. Forma el anillo de Waldeyer o
amigdalino, un anillo protector situado en la región anatómica inicial de los aparatos
digestivo y respiratorio. Está formado por:
x Amígdala lingual: cara dorsal de la raíz lingual.
x Amígdalas palatinas: a cada lado del istmo de las fauces.
x Amígdalas tubáricas: en las paredes laterales de la rinofaringe, tras la
desembocadura de las trompas de Eustaquio.
x Amígdala faríngea o adenoides: techo de la rinofaringe.
Estructura histológica
Son zonas con una superficie irregular y hendida por la presencia de criptas profundas y
abombamientos. El epitelio es difícil de diferenciar porque está invadido por linfocitos y
leucocitos. La luz de las criptas está taponada por moco y células epiteliales y
leucocitos.
Posee una cápsula de tejido conjuntivo que las delimita, haciendo fácil su extirpación.
Hay dos regiones: B (folículos) y T (parafolicular). Tienen VEA y linfáticos eferentes
que conducen la linfa hasta los ganglio linfáticos.
Amígdala lingual
Se encuentra en la mucosa de la cara dorsal de la raíz lingual. Es un epitelio
plano estratificado no queratinizado invadido por linfocitos, con criptas relativamente
cortas, poco ramificadas y separadas. Las glándulas salivales mucosas desembocan en
las criptas. Por debajo encontramos el músculo lingual.
130
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Cada cripta está asociada a tejido linfoide difuso y nódulos linfoides, esto es un
folículo lingual. Entre los folículos linguales quedan bandas anchas de tejido conjuntivo
en conexión con la cápsula.
Amígdalas palatinas
Son pares, y están a izquierda y derecha de la entrada de la faringe. Es un
epitelio plano estratificado no queratinizado y grueso (1cm) con criptas profundas,
ramificadas y próximas; los linfocitos invaden tanto el epitelio como las criptas.
Su cápsula es gruesa y emite tabiques que lobulan. Las glándulas salivales
mucosas desembocan en la superficie, fuera de la cápsula. Asociado se encuentra el
músculo faríngeo.
Amígdala faríngea
Impar, en la mucosa del techo de la rinofaringe, adherida al periostio. Es un
epitelio respiratorio con parches de malpighiano invadido por linfocitos.
No hay criptas, pero sí pliegues y recesos irregulares. La cápsula presenta
abundantes fibras elásticas. Las glándulas seromucosas desembocan en la superficie.
Se inflama con facilidad, dificultando la respiración. Son extirpables, son las
denominadas vegetaciones.
TUBO DIGESTIVO
Estructura
x Mucosa
o Epitelio
o Lámina propia
o Muscularis
mucosa
x Submucosa
x Muscular
x Adventicia
Mucosa
Es el componente más variable de la pared. El medio luminal equivale al medio
esterno. Sus funciones son:
x Protección
o Protección frente a patógenos y Ag (GALT, IgA, céllas de Paneth...)
o Protección frente a abrasión física (epitelio estratificado en esófago) y
química (moco)
o Protección frente a permeabilidad (zonula occludens) para separar
medios externo e interno
x Absorción selectiva: agua, electrolitos y nutrientes
Existen mecanismos de mejora funcional para aumentar el área de superficie
131
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
o Pliegues circulares, válvulas conniventes o de Kerkring (intestino
delgado). Son pliegues de la submucosa con orientación circunferencial.
o Vellosidades o plicae (mucosa del intestino delgado), tapizado por
epitelio estratificado cúbico. En el sentido opuesto a ellas están las
criptas de Lieberkühn.
o Microvellosidades (epitelio del intestino delgado): solo se ven en forma
de chapa estriada si son muchas e irregulares
o Glucocálix (MEC del intestino delgado)
Tras la absorción los productos difunden hacia capilares sanguíneos fenestrados y
linfáticos de la lámina propia
x Secreción
o Moco protector; lubricante y tamponamiento
o Enzimas, CIH: digestión
o Ac
Hay glándulas en
o La lámina propia (esófago, estómago, conducto anal)
o La submucosa (esófago, duodeno)
o Fuera de la pared del tubo (anejos)
El epitelio es de origen endodérmico. Forma el epitelio de revestimiento y las glándulas.
La lámina propia posee fibras colágenas, elásticas y reticulares. Presenta
GALUs, como segunda barrera defensiva frente a gérmenes, aunque disminuye en
zonas de glándulas. La vascularización es abundante, sanguíneo y linfática, y está
inervada.
En la muscularis mucosae hay músculo liso en dos estratos (circunferencial interno y
longitudinal externo) asociado a conjuntivo con fibras elásticas. Su contracción permite
movimientos de la mucosa, provocando pliegues que facilitan la absorción y la
secreción.
132
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Submucosa
Tejido conjuntivo laxo con fibras elásticas para dar elasticidad. Abundante
GALU, rica vascularización sanguínea y linfática. Aquí se sitúa el plexo de Meissner
con fibras preganglionares procedentes del vago y fibras postganglionares del plexo
mesentérico superior.
Muscular
Miocitos lisos, excepto en el 1/3 superior del esófago que son esqueléticos. Hay
2 estratos, y en algunas zonas 3. Provoca contracción: segmentarias por el estrato
circular interno, y peristálticas en el externo. Posee engrosamientos musculares, los
esfínteres:
x Faringo-esofágico: impide la entrada de aire
x Esófago-gástrico: impide el reflujo gástrico
x Pilórico: controla la liberación del quilo
x Ileocecal (distinto a la válvula íleocecal, que retrasa paso al ciego y controla
reflujo del colon)
x Anal interno: retiene heces
Las contracciones de segmentación producen la agitación del contenido luminal,
mientras que las ondas peristálticas provocan el avance del contenido luminal hacia el
ano mediante movimientos espontáneos y rítmicos. Hay un tono muscular con
contracción basal continuo que repercute en el tamaño de la luz.
Aquí está el plexo mientérico o de Auerbach, entre los dos estratos, que recibe
fibras preganglionales procedentes del vago (parasimpáticas) y fibras postganglionares
procedentes de los ganglios prevertebrales (simpáticas).
Adventicia
Es un conjuntivo laxo rico en adipocitos. Puede estar recubierto por mesotelio de
la cavidad peritoneal, la capa serosa, y su vascularización linfática y sanguínea es rica.
Vascularización
Sanguínea
Entran por la adventicia y pasa por las capas muscular y submucosa hasta la
mucosa. Aportan vasos para los músculos, las criptas y las glándulas mediante capilares
fenestrados.
El drenaje venoso es en sentido contrario y a nivel de la capa muscular empiezan a
presentar válvulas.
En el estómago la microcirculación es esencial para mantener la integridad de la
mucosa, que permite la salida del bicarbonato. En la enfermedad ulcerosa péptica se
produce la rotura de la microcirculación gástrica permite la acción destructiva del HCl,
la pepsina y las infecciones bacterianas
133
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
La alta vascularización también puede llevar a hemorragias intensas en úlceras
de estrés por erosiones superficiales de la mucosa tras traumatismo, consumo
prolongado de aspirina o corticoides…
Linfática
Es muy importante, están más profundos que los sanguíneos, pero en igual
posición.
En el estómago hay un sistema extenso de capilares entre las glándulas, más
profundo que los sanguíneos. En la submucosa están provistos de válvulas. La linfa
drena hacia la mucosa.
En el intestino delgado tiene lugar la absorción de grasa mediante los quilíferos,
en posición axial y ciego. Su endotelio es fino y contienen miocitos lisos. Tras la
digestión, la linfa es blanca y lechosa y se la conoce como quilo. En la base de la
vellosidad se anastomosa con los capilares perivasculares y forman plexo en los
músculos. Salen por la serosa.
Inervación
Aferente
Fibras aferentes viscerales generales (hambre, dolor...)
Eferentes
x Extrínsecas: simpáticas, procedentes del ganglio celíaco, y parasimpáticas,
procedentes del vago.
x Intrínsecas: SNEntérico forma plexos desde el esófago hasta el esfínter anal,
independientes del control simpático y parasimpático.
SNE conecta con SNA. Recibe axones preGg del SNPS y axones postGg del SNS.
Responde a estímulos locales y a señales procedentes de nervios del SNA
o Plexo submucoso o de Meissner
o Plexo mientérico o de Auerbach (IC y EL)
Se pueden encontrar:
o Ganglios con glía, neuronas de asociación y motoras, éstas de axón muy
corto. Sin conjuntivo dentro. Cápsula mínima e incompleta.
o Haces nerviosos, células intersticiales de Cajal.
Los SNE y SNA se integran y regulan
x El peristaltismo de la mucosa
x Los movimientos de músculos
x La secreción de glándulas de la mucosa y la submucosa
ESÓFAGO
El esófago es un tubo de naturaleza muscular que tiene aproximadamente unos 25 cm de
longitud y que se extiende entre la faringe y el estómago.
134
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
En concreto el esófago se inicia a la altura del cartílago cricoides y va a terminar 2-4cm
por debajo del diafragma.
La función principal del esófago es el transporte de los alimentos fragmentados (bolo
alimenticio), de manera que este transporte no interrumpe ni la respiración ni la
circulación cardiopulmonar.
Si se observa un esófago, o una traza de esófago al microscopio observamos que su luz
es muy irregular, que esta habitualmente colapsada. Esto se debe a una contracción
tónica sostenida de la pared del esófago.
Hay que decir que determinados excitantes como la cafeína, la teína o la cola y drogas
como el tabaco o un exceso de grasa en la dieta contribuyen a modificar esta
contracción tónica, concretamente a disminuirla; de tal forma que las personas que
consumen este tipo de productos se constata una luz más amplia en situación de no
deglución.
A luz colapsada se ensancha cuando tenemos que deglutir. Los pliegues longitudinales
desaparecen de manera temporal y vuelven a aparecer cuando ha pasado el bolo
alimenticio.
La mucosa del esófago tiene una tonalidad blanquecina que tiende a hacerse más rosada
a medida que nos acercamos a la zona de unión escamo-columnar, es decir, la transición
hacia el estómago. Esta línea rosada se le conoce con el nombre de línea Z.
En la imagen se observan tres secciones a microscopía óptica del esófago. Hay tres
secciones ya que es un tubo de bastante longitud y por lo tanto podemos apreciar
cambios a nivel histológico según vamos avanzando.
135
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Mucosa
Epitelio
El epitelio es plano estratificado no queratinizado aunque la naturaleza de este epitelio
se puede modificar con la dieta como se observa en otras especies animales. Tiene un
grosor bastante considerable, desde las 500 micras hasta más de 800.
En las preparaciones histológicas se aprecia que las células epiteliales tienen una escasa
tinción puesto que tienden a almacenar glucógeno. Como en cualquier epitelio
estratificado y con tinciones específicas podemos apreciar la presencia de células de
Langerhans, es decir, APC.
Este epitelio es una barrera firme a la entrada gérmenes y, sin embargo, tiende a hacerse
menos barrera, menos resistente y firme a medida que nos acercamos al estómago. La
transición del esófago hacia el estómago es brusca. Se aprecia la importancia de la
túnica muscular.
136
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Cuando en un individuo el esfínter esófago-gástrico falla, es frecuente que aparezcan
zonas en este epitelio que no respondan al patrón esofágico. Esto parches con un
epitelio diferente ocasionan el esófago de Barren. La naturaleza de esos parches puede
ser muy diferente. En unas personas podemos observar epitelio gástrico, en otras,
epitelio intestinal, es decir, no es un cambio fijo. La consecuencia de este esófago es que
estas islas o parches donde el epitelio es diferente (no es esofágico) tienden a ulcerarse e
inflamarse con facilidad, produciendo esofagitis por reflujo y disfagia.
En estas personas es característico un ardor intenso debido al reflujo gástrico (se
denomina pirosis). Las consecuencias a más largo plazo son todavía peores, pues en
estas personas se suelen apreciar, al cabo de un cierto tiempo, metaplasias a nivel
intestinal. Es decir, los parches a nivel esofágico acaban siendo malignos.
Lámina propia
Si pasamos a la segundad capa de la mucosa, apreciamos que es un tejido conjuntivo de
naturaleza elástica, en el que suelen abundar las fibras elásticas. Es característica del
tubo digestivo la presencia de GALUs y en este caso ya las podemos observar a nivel
del esófago.
Por lo tanto no nos extrañará el encontrar en las preparaciones linfocitos, eosinófilos,
plasmáticas o mastocitos.
A nivel de la lámina propia es característico encontrar las glándulas esofágicas
cardiales. Se sitúan en el tercio más distal, es decir, en la zona más terminal del esófago.
Si bien resulta curioso que en una población variada de personas, podamos localizar
glándulas de este tipo en el tercio esofágico. No hay una explicación de por qué en parte
de la población estas glándulas tienen esta localización.
Se trata de glándulas en número y desarrollo muy variables, de naturaleza mucosa, cuya
secreción es de pH neutro y lo que tiene es a protegernos del reflujo gástrico de
naturaleza ácida.
Muscular
La tercera capa del estrato mucoso corresponde a la muscularis mucosa. Está formada
por miocitos lisos que inicialmente forman un estrato: estrato longitudinal.
Más adelante aparece un segundo estrato de disposición circular. Tanto en uno como en
otro se observa que los miocitos están asociados al endomisio rico en fibras elásticas.
Esta capa muscular, contribuye decisivamente al aplanamiento de la mucosa en el
momento de la deglución.
Submucosa
Pasamos a la segunda capa del tubo digestivo. Es un tejido conjuntivo muy laxo, muy
rico en fibras elásticas y donde es muy frecuente detectar GALUs.
Aquí también podemos localizar otro tipo de glándulas: glándulas esofágicas. Se trata
de glándulas mucosas, de naturaleza tubuloacinar y compuestas.
137
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
La secreción que van a elaborar es una secreción que va a actuar como lubricante. Es
decir, protege al epitelio de la abrasión física que supone el roce del bolo alimenticio.
Esta secreción es muy viscosa y no es extraño encontrar que los conductos excretores
puedan enquistarse.
A este nivel encontramos un plexo nervioso: el plexo de Meissner.
Vascularización de la capa submucosa
La vascularización es muy rica, tanto sanguíneo como linfático.
En algunas ocasiones las venas pueden dilatarse. A esto se le denomina varices
esofágicas y es frecuente que ocurra como consecuencia de otro tipo de enfermedades,
como en las personas que padecen cirrosis hepática.
Por lo tanto, estas varices son secundarias a una hipertensión mortal. Debido a esta
enfermedad, u otras, o que simplemente la mucosa se ulcere, es posible que los vasos a
nivel de la submucosa se rompan. Esto lleva a una hemorragia que tiende a llevar la
sangre al estómago. El paciente va a sufrir vómitos frecuentes con sangre
(hematemesis).
Muscular
Esta tercera capa, la muscular o muscular externa, está formada por dos estratos, que en
el tubo digestivo corren siempre de manera que el estrato interno es circular y el externo
es longitudinal.
A lo largo del tubo digestivo, la mayor parte de las ocasiones vamos observar músculo
liso, no así en el esófago.
El tercio superior el esófago muestra miocitos exclusivamente esqueléticos. En la zona
media se incorporan miocitos lisos, de forma que en el tercio distal la musculatura del
esófago es exclusivamente lisa.
El tercio superior de esta capa muscular esta inervado por neuronas motoras de tipo
somático, que localizamos en el núcleo ambiguo. Las neuronas viscerales que inervan la
musculatura lisa se encuentran en el núcleo motor dorsal.
Entre ambos estratos se sitúa el plexo mientérico o plexo de Auerbach. Podemos
también mencionar la presencia de alteraciones neuromusculares que va a provocar en
los pacientes la aparición de espasmos musculares, un dolor en la zona subesternal y
una gran dificultad para la deglución.
Existe también una enfermedad, la enfermedad de Chagax-Mazza que está causada por
el parásito Trypanosoma cruzi, que provoca entre otros daños, una lesión en los plexos
nerviosos, concretamente a nivel del esófago y esto va a provocar la aparición de un
esófago extremadamente grande muy desarrollado
En esta capa muscular hay que hablar de la presencia de dos esfínteres:
x Esfínter esofágico superior: También llamado cricofaríngeo porque se sitúa a
nivel de este músculo. Este esfínter controla el inicio de la deglución.
138
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Esfínter esofágico inferior: También llamado gastroesofágico que es un esfínter
exclusivamente fisiológico, por tanto no tiene sustrato histológico ninguno.
Funciona creado unas diferencias de presiones que van a controlar el reflujo dele
estómago hacia el esófago. Luego lo normal es que estas diferencias de
presiones vayan a funcionar impidiendo el reflujo.
De forma habitual estos esfínteres están cerrados por contracción tónica. Pero es
también relativamente frecuente que el hiato esofágico inferior no cierre todo lo bien
que debería y por lo tanto el estómago pueda deslizarse en dirección proximal. Esto
supone la aparición de una hernia de hiato por deslizamiento.
Estos esfínteres están controlados por una inervación tanto parasimpática como del
sistema nervioso entérico. Tenemos así unas fibras que van a excitar, estimular la
contracción.
Son fibras en donde los mediadores encontrados son la sustancia P o la acetilcolina.
Pero también tenemos otras fibras que inhiben la contracción. En este caso los
mediadores detectados son el óxido nítrico y el péptido vasointestinal (VIP).
Tenemos también una enfermedad denominada Acalasia en la cual el esfínter esofágico
inferior no se distiende. Es decir, está más cerrado de lo que debería, dificultando
enormemente la deglución. Esta acalasia se produce por un defecto durante el desarrollo
embrionario, de tal manera que las neuronas que deberán inhibir esta contracción
mediante los péptidos mencionados (NO, VIP) no existen, se pierden y por lo tanto la
falta de neuronas inhibidoras lleva a una contracción permanente.
Adventicia
Es la última de las capas del tubo digestivo. Es de nuevo un conjuntivo laxo y puede
adherirse, fijarse a suturas contiguas. Esto puede ocurrir a nivel del esófago, en varias
zonas sobre todo en la porción distal. De manera que en estas zonas en vez de hablar de
adventicia hablamos de serosa (pues está cubierta por peritoneo visceral).
ESTÓMAGO
Región dilatada del tubo digestivo que se encuentra bajo el diafragma (1,5L).
Hay abundantes pliegues longitudinales (arrugas), y elevaciones y surcos (mamelones).
Sus funciones son:
x Digestión (homogeneizar)
x Segregar encimas y ClH
x Segregar factor para la absorción de la
vitamina B12
x Almacena el quimo
Según su motilidad se divide en dos regiones, una desplaza en sentido oral y otra en
sentido caudal. La región oral y el EEI se relajan en deglución. El tono muscular se
adapta al volumen sin alterar la presión interna.
139
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
La contracción caudal mezcla y propulsan el contenido a la unión
gastroduodenal. El contenido sólido es retropropulsado para facilitar la mezcla y la
digestión mecánica.
Según el tipo de glándulas
x Cardias: zona próxima al orificio esofágico. Glándulas cardiales
x Antro o región pilórica: parte proximal respecto al píloro. Glándulas pilóricas o
antrales.
x Región fúndica: Glándulas fúndicas o gástricas.
Estructura
x Arrugas gástricas con ejes de submucosa, profundas en región pilórica y fúndica.
Al distenderse desaparecen. No son un mecanismo de extensión de la superficie.
x Elevaciones o surcos que aumentan algo el área.
x Fovéolas, fositas, fosetas o criptas gástricas: orificios en la mucosa. Son
invaginaciones profundas del epitelio en continuidad con glándulas. Son menos
profundas en fundus y muy profundas en antro
Mucosa
Epitelio de revestimiento de la mucosa
La superficie y las fovéolas tienen epitelio prismático simple, con células
mucosas (gránulos mucígenos, PAS+). Segregan bicarbonato procedente de células
oxínticas.
El mucígeno es viscoso y turbio, cuya función es protectora frente a la abrasión del
quimo y al ácido. Posee zonula occludens y adherens. Favorece la absorción de aguas,
sales y componentes liposolubles (alcohol y fármacos).
Lámina propia
Conjuntivo laxo muy escaso por la abundancia de glándulas. Es rico en fibras
colágenas y reticulares, sin elásticas. Las células linfoides son abundantes, y posee
inclusiones de Rusell en sus plasmocitos por acumulación de Igs. En la zona pilórica
hay cordones de miocitos.
140
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Las glándulas se abren al surco de las fóveolas. Tipos
x Cardiales: drenan al fondo de las fositas del cardias
x Antrales o pilóricas: drenan al fondo de las fositas del antro o región pilórica,
menos profundas
x Gástricas o fúndicas: drenan al fondo de las fositas del fundus histológico,
también de menor profundidad.
Glándulas
Como ya se ha dicho, las glándulas son un importante componente de la lámina propia
gástrica. Cada una posee:
x Base o fondo: es la zona de mayor amplitud y suele dividirse en ramas dos o tres
veces
x Cuello: es estrecho y bastante largo
141
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Istmo: sirve de conexión entre el cuello glandular y la fovéola en cuyo fondo
desemboca la glándula. Contiene células madre, que al dividirse se diferencian
en células mucosas, las cuales ascienden para renovar el epitelio (que no sólo es
la superficie luminal de revestimiento, sino también sus invaginaciones, las
fositas), o en células secretoras, que descienden para renovar la glándula. En
condiciones normales, el número de células madre es bajo, pero si se produce
algún tipo de daño de forma continua aumenta su cantidad y actividad con fines
de reparación.
Glándulas cardiales
Las glándulas cardiales son muy largas, ocupan casi la mitad del espesor de la mucosa,
y tienen una luz muy estrecha. Se definen como glándulas tubulares ramificadas
enrolladas, y en ellas podemos encontrar fundamentalmente dos tipos celulares:
x Células mucosas
x Células endocrinas
Glándulas antrales
Las glándulas antrales tienen una longitud menor, ocupando tan solo un cuarto de la
mucosa, pero su luz es más amplia. Al igual que las cardiales, son glándulas tubulares
ramificadas enrolladas. A los dos tipos celulares que poseen las glándulas cardiales, se
les añade un tercero en este caso:
x Células mucosas
x Células endocrinas
x Células oxínticas
Glándulas gástricas o fúndicas
Las glándulas fúndicas son las de mayor longitud, ocupan prácticamente todo el espesor
de la capa mucosa, y poseen una luz muy estrecha. A diferencia de las anteriores, que
eran tubulares enrolladas ramificadas, este tipo de glándulas son tubulares rectas. A los
tipos celulares de los que disponen las glándulas antrales se les añade uno más en este
caso: las células principales.
142
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Células mucosas del cuello
Están situadas en el cuello de la glándula fúndicas. Son mucho más cortas que las
células mucosas superficiales, esto es, las del epitelio de revestimiento, y contienen
menos mucígeno en el citoplasma. Además, los gránulos de mucígeno, que como en el
caso de los de las células de revestimiento son PAS+, no coalescen, por lo que el polo
de la célula está cerrado. El citoplasma es pálido y el núcleo tiende a ser esferoidal y a
situarse en posición basal.
Las células mucosas del cuello secretan un moco soluble y líquido, a diferencia del
moco insoluble y turbio producido por las células mucosas superficiales. La liberación
de los gránulos de mucígeno es inducida por la estimulación vagal, en consecuencia, la
secreción desde estas células no ocurre en el estómago en reposo.
Células endocrinas
Se encuentran en todos los niveles de la glándula, aunque tienden a acumularse más en
la base. Como es lógico, se encuentran dispersas entre todos los demás tipos celulares.
Se trata de células que forman parte del sistema APUD.
La nomenclatura de las células enteroendocrinas en la bibliografía antigua hacía
referencia a su capacidad para ser teñidas con sales de plata o cromo: se llamaban
también cromafines, argentafines y argirófilas.
En el polo basal de estas células encontramos gránulos de secreción, que permiten
segregar hormonas que estimulan las funciones gástricas:
x Serotonina: estimula peristalsis
x Gastrina: estimula peristalsis y secreción de HCl. Activa la CCK.
x Histamina: estimula la secreción de HCl
x Somatostatina: inhibe la secreción de HCl y la motilidad gástrica.
x Grelina: estimula la secreción de GH y los centros del apetito
Por supuesto, la regulación corre a cargo de muchas más hormonas, sólo hemos
mencionado las más relevantes.
Células oxínticas
Son células que se sitúan en el cuello de la glándula, entre las células mucosas del
cuello, y en la parte más profunda de la glándula. Por tanto, están en toda la glándula,
aunque tienen tendencia a ser más abundantes en los segmentos superior y medio del
cuello. Tienen forma prismática, pero este prisma forma una prominencia o
abultamiento en la parte de la base, que está apoyada en la lámina basal.
Su núcleo es redondo y central, y su citoplasma eosinófilo. No presentan gránulos y en
general poseen pocos orgánulos, a excepción de las mitocondrias, que ocupan un 40%
del volumen citoplasmático, de ahí que el consumo de O2 de este tipo de células
quintuplique al de una normal.
La función principal de las células oxínticas es la secreción de HCl y factor intrínseco.
Sistema tubulovesicular y canalículos intracelulares
143
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Al examinarlas con el microscopio electrónico de transmisión, vemos que estas células
presentan un extenso sistema de canalículos intracelulares en contacto con la luz de la
glándula, en el polo apical. Este sistema se puede visualizar también mediante
impregnaciones argénticas, adquiere forma de garra si utilizamos este tipo de tinción.
Los canalículos son en realidad invaginaciones de la membrana plasmática apical, que
deja una red tortuosa de canalículos profundos que llegan hasta la membrana basal
aumentando la superficie de la membrana plasmática. La membrana invaginada es muy
rica en ATPasas Na/K (80%), que se encarga de sacar protones a la luz de la glándula y
meter K en el citplasma. También hallamos en ella canales para Cl. Desde la superficie
de los canalículos se proyecta una gran cantidad de microvellosidades, y en el
citoplasma contiguo a ellos hay un complejo sistema de membranas tubulovesicular.
El sistema tubulovesicular constituye un reservorio de membrana plasmática apical, y
ambos se modifican según la actividad: ante una estimulación de la secreción de HCl, el
sistema tubulovesicular disminuye y el número de microvellosidades aumenta,
consiguiendo que la superficie se quintuplique, y las crestas mitocondriales también
experimentan cambios de actividad.
Histofisiología del sistema de canalículos intracelulares
El CO2 proveniente del plasma sanguíneo entra en
contacto con la membrana plasmática basal. Esta
membrana es muy rica en anhidrasa carbónica, que
se encarga de catalizar la reacción: H2O + CO2 Æ
HCO3-, el cual se autodisocia dando iones
bicarbonato y H+. La ATPasa presente en la
membrana de los canalículos se encarga de sacar
estos protones e incorporar a la célula K+. A la par,
los canales de Cl- toman iones cloro también de la
membrana basal y vuelven a sacarlos al medio a
través de la membrana apical. En la luz de los
canalículos intracelulares, Cl- se une a H+ y se forma
HCl. El omeoprazol es un fármaco empleado para
tratar úlceras gástricas, que impide la formación de
clorhídrico porque bloquea la ATPasa Na/K.
El HCO3- formado como resultado de la disociación tras la reacción de la anhidrasa
carbónica sale por la membrana plasmática basal y llega a los capilares fenestrados
presentes en la lámina propia de la mucosa gástrica. Desde ahí, fluye hacia zonas más
superficiales y difunde hacia las células mucosas y hacia la capa de moco, aunque no
sea secretado por ellos. Ejerce un efecto tamponador importante, ya que se encarga de
aumentar el pH.
La regulación de todo este proceso corre a cargo de:
1. Gastrina: es una hormona peptídica gastrointestinal que constituye el principal
mecanismo de estimulación de la célula parietal. Por ello, si se forma un tumor
de células endocrinas productoras de gastrina vamos a tener una hipersecreción
144
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
de HCl, lo que va causar al individuo úlceras gástricas y duodenales crónicas:
este es el mecanismo del Síndrome de Zollinger-Ellison
2. Ach: es un neurotransmisor liberado por las fibras nerviosas del SNA que
también estimula a la célula parietal
3. Histamina: estimula del mismo modo
En los seres humanos, el factor intrínseco es secretado por las células parietales (En
otras especies, las encargadas de esta tarea son las células principales). Su secreción se
desencadena por la estimulación de los mismos receptores que activan la del
clorhídrico, a saber: acetilcolínicos, histamínicos y, en especial, de gastrina. El factor
intrínseco es una glucoproteína que forma un complejo con la vitamina B12 en el
estómago y en el duodeno, un paso necesario para la ulterior absorción de la vitamina en
el íleon. Por este motivo, tanto la aclorhidia (enfermedad autoinmune caracterizada por
la ausencia de células oxínticas) como la enfermedad ulcerosa péptica (caracterizada por
la destrucción del epitelio) conducen a una anemia perniciosa.
Células principales
Las células principales son células secretoras de proteínas típicas. Se trata de células de
forma prismática, situadas en el fondo de las glándulas. Son difíciles de preservar de
cara a su observación en una tinción histológica.
Poseen un núcleo, RER y Golgi basales, lo cual les da en esta zona un aspecto basófilo
característico. En la porción apical encontramos los gránulos de secreción, que
contienen pepsinógeno y lipasa, así como abundantes microvellosidades.
Al contacto con el jugo gástrico ácido, el pepsinógeno se convierte en pepsina, una
enzima proteolítica. La regulación de la secreción de pepsinógeno y lipasa se lleva a
cabo por medio de la secretina.
Jugo gástrico
El jugo gástrico, por tanto, y reuniendo lo ya estudiado, consta de dos secreciones:
x Constitutiva: siempre está presente. Es la capa de moco, que tiene función
protectora, junto con el HCO3-. Entre ambos mantienen un pH neutro y separan
el quimo de la mucosa.
El Helycobacter Pylori se halla en un 20% de los individuos a los 20 años, y se encarga
de la producción de amoniaco y proteasas citotóxicas. Aunque muchos individuos son
asintomáticos, algunos presentan úlceras pépticas (melenas, hematemesis y dolores
súbitos y persistentes se asocian frecuentemente a la presencia en el estómago de esta
bacteria).
x Facultativa: no está siempre presente. Está constituida por la pepsina y el HCl, y
es mucho más agresiva:
o HCl: proporciona un pH ácido, y tiene una gran importancia en la
digestión de proteínas, tanto por sí solo como por su acción sobre el
pepsinógeno, convirtiéndolo en pepsina. Tiene también función
bacteriostática.
o Pepsina: hidroliza proteínas a pequeños péptidos que posteriormente son
convertidos en aminoácidos
145
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Además, recordemos que también encontramos agua e iones Na+, K+ y Cl-. Así, los
pacientes con vómitos crónicos requieren un aporte extra de NaCl, dextrosa y K+ para
evitar una acidosis metabólica hipopotasémica. También encontramos factor intrínseco
unido a B12.
Recordemos que aún no hemos salido de la mucosa, ya que todas estas glándulas
pertenecen a la lámina propia
Muscularis mucosae
Se divide en dos estratos, un estrato circular interno y uno longitudinal externo, aunque
hay zonas en las que puede aparecer un tercer estrato. Su contracción comprime la
mucosa facilitando la expulsión del jugo gástrico de las fovéolas.
Submucosa
Se trata de un tejido conjuntivo laxo rico en adipocitos. NO PRESENTA
GLÁNDULAS.
Se encuentra ampliamente vascularizado, disponiendo de un plexo arterial, venoso y
linfático. Además, es a este nivel donde se encuentra el plexo nervioso de Meissner o
submucoso. También hallamos GALT.
Muscular
Se trata de tejido muscular liso, en el que podemos diferenciar tres estratos incompletos:
x Interno oblicuo
x Circular medio: forma el esfínter pilórico, cuya relajación permite la salida del
quimo del estómago hacia el ID (duodeno)
x Longitudinal externo
Estos tres estratos se disponen en espiral, de manera que se posibilita la adaptación del
estómago a los cambios de volumen sin que se produzcan cambios de presión en su
interior. Entre las capas musculares encontramos el plexo nervioso de Auerbach.
Serosa
Se trata de una adventicia cubierta por peritoneo.
INTESTINO DELGADO
Generalidades
Se trata de la porción del tubo digestivo situada entre el estómago y el intestino grueso.
En estado relajado mide unos 7-8 m, in vivo 5 metros. Su contenido es el alimento
semidigerido, conocido como el quilo. Tiene tres funciones básicas:
x Secreción de enzimas y hormonas
x Absorción
x Digestión
146
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Estructura anatómica
Anatómicamente, podemos dividir el intestino delgado en tres regiones principales:
x Duodeno: es la porción más corta, tiene unos 25 cm de longitud y es muy ancho.
Va formando una C desde el píloro hasta el ángulo duodeno-yeyunal alrededor
de la cabeza del páncreas. A él se abren los conductos pancreático y colédoco.
x Yeyuno: 2 m y medio
x Íleon: mide 3 m y medio y termina a la altura de la válvula iliocecal.
Tanto el yeyuno como el íleon penden por medio del mesenterio y son móviles.
Estructura histológica
Presenta varios mecanismos de aumento de la superficie:
1. Pliegues circulares, válvulas conniventes o válvulas de Kerkring: se trata de
pliegues permanentes con un eje de submucosa y forma semilunar. Son
especialmente abundantes en la porción distal del duodeno y en el yeyuno.
2. Vellosidades o plicas: se trata de proyecciones de la mucosa de 1,5 mm de
longitud, muy abundantes a la altura del duodeno y del yeyuno proximal. En
adultos tienen forma digitiforme, mientras que en el duodeno infantil tienen
forma foliácea. La altura máxima mencionada la alcanzan en el yeyuno
proximal.
3. Criptas
Criptas de Lieberkuhn
Recordemos que la secreción sale de la base de las vellosidades y que entre cripta y
cripta hay tejido conjuntivo y aparece un eje.
La lámina propia se encuentra ocupada por profundas criptas de Lieberkühn, en la que
encontramos enterocitos, células caliciformes, endocrinas, madre y de Paneth:
147
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Enterocitos
Son morfológicamente similares a los del epitelio de revestimiento aunque son algo más
pequeños. Su función principal es más secretora que absortiva: secretan agua y sales, de
manera que juegan un papel fundamental en que el contenido de la luz no se compacte
según avanza y se produce la absorción (mantiene el contenido en estado líquido). Para
esto la absorción debe estar equilibrada con la secreción de líquido de los enterocitos
secretores.
Células caliciformes
Son similares a las del epitelio de revestimiento.
Células endocrinas
Se encuentran en la base de las glándulas. Son capaces de migrar desde aquí, luego no
es extraño encontrarlas más arriba y pueden diseminarse por todo el epitelio glandular y
de revestimiento. Son argirófilas, argentafines y cromafines. Presentan gránulos de
imagen variable según el contenido, encontrándose sobre todo en la parte basal del
citoplasma. Forman hormonas y péptidos neuroendocrinos.
Secretan una gran variedad de hormonas:
x Colecistoquinina (CCK): se encuentra en duodeno y en yeyuno. Tiene efectos
diversos: promueve la salida de la bilis, inhibe el vaciado gástrico (inhibe a los
miocitos lisos, la musuclatura del estómago), estimula el crecimiento del
páncreas, estimula la capa muscular de la vesícula biliar para que se contraiga y
la secreción de jugo pancreático. A nivel del estómago aumenta la secreción del
bicarbonato, al estimular a las parietales. Además estimula el crecimiento del
páncreas y de enzimas pancreáticas.
x Secretina: se libera en duodeno. Estimula el crecimiento del páncreas, la
contracción de la vesícula biliar y la secreción de enzimas pancreáticas y de
HCO3-. Inhibe la secreción ácida a nivel del estómago
x Polipéptido inhibidor gástrico (GIP): se libera en duodeno y yeyuno. Estimula la
liberación de insulina (páncreas) e inhibe la secreción ácida (estómago). Actúa
sobre los miocitos del intestino y del estómago, estimulando el peristaltismo.
x Motilina: se libera en duodeno y yeyuno. Estimula la motilidad gástrica e
intestinal, actuando sobre la musculatura del tubo digestivo (intestino delgado y
estómago). Estimula la contracción.
Además de hormonas, las criptas secretan péptidos como:
x Péptido vasoactivo intestinal (VIP): hormona que actúa en el sistema de la
insulina. Estimula las secreciones intestinales y pancreáticas (a fin de liberar más
insulina), inhibe los esfínteres y la musculatura en general. Estimula la
secreción de las propias criptas y tiene un efecto paracrino.
x Encefalinas: estimulan la contracción muscular e inhiben las secreciones
intestinales.
Asimismo, las células endocrinas segregan una serie de hormonas paracrinas:
148
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Somatostatina: hormona inhibidora de la liberación de gastrina y otras hormonas
gastrointestinales, así como de la secreción ácida gástrica.
x Histamina: hormona estimulante de la secreción ácida gástrica.
Células madre
Permiten renovar el epitelio de revestimiento y glandular y están en la base de las
criptas. Se dividen activamente y migran (excepto las células de Paneth pues se
encuentran en la base). Tienen una tasa de renovación alta, que es máxima en el yeyuno.
Células de Paneth
Se sitúan en el fondo de las criptas y son más numerosas según avanzamos en el
intestino hacia distal. Son células altas y prismáticas de núcleo redondeado, con una
importante dotación de orgánulos, lo que hace que citoplasma basal sea basófilo (gran
desarrollo del RER). En la zona apical encontramos gránulos refringentes eosinófilos.
El citoplasma apical tiene orgánulos eosinófilos y birrefringentes.
En estos gránulos hay una mezcla de sustancias: proteínas ricas en arginina (que hacen
destruir las bacterias o su membrana plasmática), enzimas como la lisozima, TNF
(proinflamatoria en respuesta a lesiones)… Encontramos también: zinc y defensinas o
criptidinas, capaces de perforar las membranas de bacterias y parásitos, creando canales
que aumentan la permeabilidad de éstos y generando con ello una respuesta
inflamatoria. Además, son capaces de estimular el acercamiento de células dendríticas,
facilitando la reacción inmunitaria.
Las defensinas actúan perforando la membrana de la bacteria o parásito, lo que aumenta
la permeabilidad de la membrana del patógeno y se potencia la reacción contra ese
patógeno.
Las células de Paneth regulan el nivel de la flora bacteriana intestinal y eliminan
mediante fagocitosis, bacterias y protozoos. Presentan picos de secreción después de
una ingesta.
Hay personas con una tendencia a tener inflamaciones, relacionadas con la mutación en
el gen NOD2, que hace que haya una baja síntesis de defensinas.
Luego estas células secretan proteínas antimicrobianas
para limitar el contacto entre las bacterias y los
enterocitos. Las proteínas antimicrobianas son
retenidas dentro de la capa de moco intestinal. Por
eso, esta protege la superficie de los enterocitos
mediante dos mecanismos: creación de una barrera
que limita el acceso directo de las bacterias luminales
al epitelio y concentración de las proteínas
antimicrobianas cerca de la superficie de la membrana
plasmática del enterocito.
Clínica
x Enfermedad de Crohn: inflamación de la mucosa en
el intestino grueso e íleon.
149
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Colitis ulcerosa: inflamación de la mucosa en el intestino grueso.
En ambas situaciones, el intestino delgado las criptas muestran infiltración de
neutrófilos y las criptas acaban ocluidas, lo que conlleva la degeneración de las
glándulas. Los granulocitos llegan a la mucosa e invaden la muscular, de modo que el
intestino se obstruye por una sustitución progresiva en la que aparece conjuntivo. Se va
a atrofiar toda la mucosa y se forma granulosa en toda la capa muscular.
Esto da lugar a una reacción inflamatoria por linfocitos Th1 (enfermedad de Crohn), que
producen TNF alfa, INF gamma y IL12 o Th2 (colitis ulcerosa), que producen IL4 e
IL5. En ambas se secretan citocinas proinflamatorias. El tratamiento usado son
anticuerpos contra TNF alfa. Además IL10 parece actuar recuperando la tolerancia de
los linfocitos T a la flora bacteriana.
Muscularis Mucosa
Presenta un estrato interno, con los miocitos dispuestos circularmente; y otro externo,
con los miocitos dispuestos longitudinalmente. Permite mezclar mejor el contenido
intestinal gracias a la contracción y a la secreción de las gládulas salga a la luz del
intestino.
Submucosa
Es un conjuntivo laxo con adipocitos
frecuentes y fibras elásticas y
acúmulos linfoides. En el duodeno
aparece la glándula de Brunner.
La submucosa destaca por la gran
vascularización tanto de vasos
sanguíneos como linfáticos.
Es frecuente encontrar unidades
linfoides y el plexo de Meissner. Solo
hay
150
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
glándulas a nivel del duodeno: glándulas de Brunner.
Es una glándula tubular ramificada de naturaleza mucosa, cuya secreción es drenada al
fondo de las criptas de Lieberkuhn. Es una secreción de glicoproteínas que van a
permitir dar un pH neutro o básico al moco duodenal, combatiendo a su vez el ambiente
ácido. Esto es esencial para procesar el quimo y activar las enzimas pancreáticas.
Contribuyen a la capa de moco (que tiene bicarbonato). Se tienen que activar las
enzimas pancreáticas para lo cual necesitamos un ambiente ácido.
También se detecta en estas glándulas la urogastrona, un factor de crecimiento
epidérmico (EGF). La urogastrona es resistente al ambiente ácido y a enzimas
digestivas y actúa en diferentes órganos, puesto que hay receptores suyos en hepatocitos
y glándulas gástricas.
La urogastrona inhibe la secreción ácida gástrica. Estimula asimismo la proliferación de
las células madre, es decir, estimula la renovación y crecimiento de la mucosa digestiva.
Parece ser que interviene en situaciones de reparación o regeneración tras daño tisular.
Capa Muscular
Presenta un estrato interno, con los miocitos dispuestos circularmente; y otro externo,
con los miocitos dispuestos longitudinalmente. Permite mezclar mejor el contenido
intestinal. En ella, entre los dos estratos, encontramos el plexo de Auerbach.
Tenemos miocitos que se contraen, provocando dos tipos de contracciones:
x La capa interna forma contracciones de segmentación que movilizan el
contenido luminal, haciendo que se mezclen con los jugos que se vierten al
intestino, y aseguran que el jugo esté en contacto con la mucosa.
x La capa externa es responsable del peristaltismo, que permite el avance del
contenido a porciones distales. El avance depende de unas ondas que son ondas
migrantes motoras (ondas peristálticas).
Las ondas peristálticas se producen también en ayuno para que el intestino delgado se
vacíe y son estimuladas por la motilina. La motilina se segrega permanentemente
cuando no comemos y se inhibe su liberación cuando comemos. Si no se dan
movimientos peristálticos, se retiene alimento y crece la flora bacteriana, pues se ha
acumulado el contenido luminal y hay bacterias a mogollón.
Tenemos el esfínter iliocecal: se dispone entre el íleon y el ciego. Está parcialmente
contraído. Esta contracción tónica está impidiendo que el contenido avance hacia el
ciego. Si el ciego se distiende, entonces se produce, como reacción refleja, una
inhibición del peristaltismo del íleon (inhibición de la longitudinal) y una contracción
del esfínter (contracción capa circular). El íleon evita el peristaltismo hasta que se hayan
absorbido los nutrientes.
Tras una ingesta, lo que se produce es que el íleon sufre peristaltismo, la relajación del
esfínter y todo esto permite el avance del contenido y que este alcance el ciego.
También tenemos la válvula iliocecal. No hay que confundir el esfínter con la válvula
ileocecal: la válvula (válvula bicúspide) son pliegues de la mucosa orientados hacia el
ciego, de tal forma que una vez que el ciego se llena, los pliegues (la válvula) se cierran,
151
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
impidiendo que el contenido retroceda hacia el íleon. La válvula funciona por el cambio
de presiones, no por contracciones musculares.
Capa Adventicia
Sólo se encuentra en el duodeno, pues en el yeyuno y el íleon se encuentra asociada al
peritoneo, formando una serosa.
Características diferenciales
Duodeno
Encontramos vellosidades de tipo foliáceo, sobre todo en la infancia. Encontramos
glándulas de Brunner. En el duodeno se produce la llegada de jugo pancreático y la
bilis. No se encarga de la digestión final y permite la emulsión de las grasas.
Yeyuno
Es la zona donde se da un máximo desarrollo de las válvulas conniventes, vellosidades
y microvellosidades. Las vellosidades son digitiformes. Las células presentan una alta
tasa de división y es la zona donde se produce la máxima absorción de nutrientes.
Íleon
Tenemos un desarrollo máximo del tejido linfoide (GALT), y encontramos el esfínter y
la válvula ileocecales. Se produce la absorción específica de sales minerales y
reabsorción de sales biliares y vitamina B12, esta última sólo si está unida a la proteína.
INTESTINO GRUESO
El intestino grueso conforma la porción distal del tubo digestivo. Tiene un diámetro
mayor que el intestino delgado (5 cm.) y una longitud de 1.5 m. Anatómicamente se
divide en colon, recto y ano. El ciego es donde se sitúa además el apéndice vermiforme.
Entre las funciones del intestino grueso contamos las de absorción, secreción,
compactación de heces y almacenamiento temporal de las mismas.
Heces
Las heces se componen en un 75% de agua, y el 25% restante de grasa, proteínas no
digeridas, sustancias inorgánicas, células muertas y pigmento biliar. La composición y
el olor de las depende de la dieta y la flora bacteriana intestinal de cada persona.
La flora bacteriana es anaeróbica, y como consecuencia de ello se dan reacciones de
putrefacción y fermentación que conllevan la producción de gas (CO2, metano y H2). En
las heces encontramos vitaminas K y B12, ácido sulfhídrico, mercaptanos e índoles.
Estructura histológica
Al igual que todo el tubo digestivo, el intestino grueso presenta una mucosa con
epitelio, lámina propia y muscularis mucosa; una submucosa, una capa muscular y una
adventicia.
152
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
En la cara interna del intestino grueso encontramos los pliegues semilunares o válvulas
colónicas. Estos pliegues están formados por mucosa que se contrae por acción de la
muscularis mucosa. No encontramos válvulas conniventes ni vellosidades.
En la cara externa del intestino grueso encontramos las haustras y los apéndices
epiploicos.
Mucosa
Formada por epitelio. Este epitelio es similar al del intestino delgado, pues es prismático
simple con células calicifomres. Lo que le diferencian es que el número de caliciformes
es mucho mayor.
El epitelio presenta escasas vellosidades, alguna en la parte más proximal. Encontramos
abundantes criptas de Lieberkühn, más largas que en el intestino delgado. En el epitelio
encontramos diversos tipos celulares: enterocitos, células caliciformes, células madre y
células endocrinas.
Enterocitos
Son las células principales del intestino grueso, sobre todo en el colon (proporción 4:1
con caliciformes). La cantidad de estas células va disminuyendo, hasta que en el recto se
encuentran en el mismo número que las células caliciformes (proporción 1:1). Y al final
es mayor el número de células caliciformes.
Estos enterocitos son muy similares a los del intestino delgado, aunque son más altos.
Su membrana apical sigue teniendo microvellosidades, pero no son tan numerosas y
regulares como en el intestino delgado, por lo que no se observa chapa estriada al
óptico. Estos enterocitos presentan una mayor proporción de mitocondrias. El espacio
intercelular está muy dilatado.
153
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Esta región acoge agua (1L/diario), electrolitos, vitaminas y minerales que los
enterocitos absorben. Se recoge más agua ya que como ya hemos absorbido todos los
nutrientes ahora lo que queremos es conseguir agua. Es por eso que en el intestino
grueso, el transporte activo (y masivo) de Na+ arrastra mucha agua, lo que es
fundamental para que las heces sean cada vez más semisólidas. Los enterocitos
absorben asimismo determinados fármacos.
Los enterocitos segregan además K+ y HCO3-, secreción que presenta un control
endocrino por aldosterona.
Células caliciformes
Las células caliciformes son aquí más numerosas que en el intestino delgado,
encontrándose su máxima densidad en las criptas de Lieberkuhn.
Las células caliciformes segregan de manera constante un producto mucoso que juega
un papel fundamental para proteger (importante para la lubricación) la mucosa frente al
contenido luminal que se solidifica y erosiona la pared. El moco facilita asimismo el
avance de las heces y su compactación.
Células madre
Se sitúan en la base de las criptas de Lieberkühn y presentan un índice de renovación
similar a las del intestino delgado.
Las endocrinas son similares a las del ID pero algo menos abundantes. Se sitúan
preferentemente en la zona inferior de las criptas pero no es exclusivo, también pueden
estar dispersas en la zona superior. Multitud de péptidos y hormonas que están
segregadas en el Intestino Grueso.
Células endocrinas
Estas células son menos numerosas que en el intestino delgado, y se encuentran
principalmente en el colon, en la parte inferior de las criptas.
En el intestino grueso, las células endocrinas segregan glucagón, sustancia P,
cromogranina, somatostatina, encefalinas, histamina, péptido Y, VIP, GLP-1 (similar a
glucagón 1)…
La mayoría actúan sobre el estómago y la mayoría actúan inhibiendo la secreción
gástrica y la liberación de alimento. La somatostatina actúa estimulando la síntesis de
gastrina y la secreción ácida gástrica. GLP-1 inhibe la secreción acida y el vaciado
gástrico y estimula la liberación de insulina. El péptido Y inhibe el vaciado gástrico y la
secreción ácida.
Lámina propia de la mucosa
Es un conjuntivo bastante laxo con un espacio de conjuntivo, la meseta colágena. Esta
meseta tiene un grosor de 5 μm, formada por colágeno y proteoglucanos sulfatados y
está situada entre la lámina basal subeptielial (donde se apoya el epitelio) y las láminas
basales de los capilares fenestrados. Es una zona que regula el transporte de agua y sales
(electrolitos) que llegan hasta la sangre desde los enterocitos. La meseta colágena está
154
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
muy hipertrofiada en personas con pólipos en el colon (puede llegar a ser el triple de
grosor).
La lámina propia presenta una importante cantidad de fibroblastos alrededor de las
criptas formando una vaina. En estas células hay gran cantidad de mitosis. De aquí,
estos fibroblastos “hijos” migran en paralelo a las criptas, hacia la superficie. Lo curioso
es que estos procesos ocurren en sincronización con la división de las células madre de
las bases de las criptas: es por ello que se postula (aunque no se sabe con certeza) que
estos fibroblastos que forman la vaina dan lugar a los macrófagos de la lámina propia.
La lámina propia presenta un desarrollo muy marcado del GALT, en continuidad con
las placas de Peyer del íleon. No encontramos vasos linfáticos, lo cual se relaciona con
el hecho de que en buena parte del colon hay un bajo índice de metástasis cuando se da
un cáncer colónico.
Muscularis Mucosae
Presenta un estrato interno, con los miocitos dispuestos circularmente; y otro externo,
con los miocitos dispuestos longitudinalmente. Desaparece bruscamente al llegar al
conducto anal. Sí encontramos asociados a los miocitos una red de linfáticos (aquí sí
hay vasos linfáticos).
Submucosa
Es un conjuntivo laxo muy vascularizado, con una alta presencia de adipocitos. En esta
zona no encontramos glándulas (solo hay glándulas en el conducto anal).
Los miocitos longitudinales de las tenias van de trecho en techo emitiéndose en la capa
circular interna. Cuando los miocitos se contraen, también se contraen a lo largo y la
porción que se ha metido se encuentra en la circular. De manera que el intestino grueso
al contraerse forme sacos o hasuras. En el intestino grueso, con la propia contracción de
la musculatura se producen esos sacos, lo que indica es que en esas porciones hay zona
contraída y zona no contraída.
Muscular
El estrato circular interno es similar al del intestino delgado. En el conducto anal se
engrosa y forma el esfínter anal interno.
El estrato longitudinal externo solo existe en parte del intestino grueso: en el recto, el
conducto anal y el apéndice (esta capa es similar a la del intestino delgado). En el ciego
y el colon no hay capa longitudinal externa y los miocitos longitudinales se agrupan
formando tres bandas: las tenias del colon (aunque también hay en el ciego).
Los miocitos salen de la tenia y pasan al estrato circular interno de trecho en trecho,
haciendo que se observen segmentos (que se contraen de modo independiente) como
hundidos: los sacos o haustras. Estos sacos tienen unas contracciones locales,
independientes entre sí, de forma que se puede observar una zona contraída y una zona
no contraída.
La capa longitudinal provoca el peristaltismo, que permite el avance del contenido hacia
el ano, normalmente una vez al día.
155
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Capa Adventicia
En algunas zonas es una serosa (tejido conjuntivo laxo cubierto por mesotelio del
peritoneo en algunas zonas). Presenta unas proyecciones de la serosa asociadas con
adipocitos: los apéndices epiploicos.
Ciego
Es una expansión del intestino grueso cerrada por un extremo. Zona de máxima
riqueza glandular (criptas de Lieberkuhn) y de células caliciformes, con gran desarrollo
de GALT y tenias.
Hay una evanginación digitiforme, el apéndice, a unos 3 cm del íleon. La luz es
pequeña e irregular, con restos que producen oclusión. No hay vellosidades, aunque sí
posee chapa estriada. Las criptas son irregulares y escasas, pobres en caliciformes. El
GALT está muy desarrollado, haciendo que la pared esté engrosada. Tiene una capa
continua de nódulos con centros germinativos, se trata de un órgano linfoide secundario.
Se caracteriza, histológicamente, por:
x Muscularis mucosas mal desarrollada
x Submucosa laxa con adipocitos y vasos. Se hace más densa y fibrosa en adultos.
x La muscular es menor, con capa circular interna gruesa y longitudinal externa
delgada.
El apéndice sufre cambios inflamatorios crónicos, inexistentes en recién nacidos y
disminuyen en la infancia, quedando solo vestigios en el adulto. Esto puede llevar a una
peritonitis.
La apendicitis aguda consiste en una ulceración de mucosa conbstrucción de la luz.
Aumenta la presión
intraluminal,
156
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
produciéndose estasis y lesión de vasos y, finalmente, perforación y peritonitis.
Colon
Se divide en con
ascendente, transverso, descendente y sigmoide. Hay material semisólido en
descendente y sigma. Son característicos las tenias y los apéndices epiploicos. Aquí se
produce la absorción de agua y electrolitos, además de la captación de fármacos
(sedantes, analgésicos u esteroides) administrados como supositorios.
El tránsito retrasado por el colon produce estreñimiento grave. La enfermedad
de Hirschprung (megacolon congénito) se debe a la ausencia de SNE en un segmento
del colon distal (aganglinoides) por detención de la migración de precursores; hay hasta
tres genes implicados (RET, EDN3, EDNRB). La zona aganglionar está contraída de
forma permanente y no permite la entrada del contenido, hay dilatación por retención de
heces y, finalmente, perforación. Recordar que la ausencia de RET provoca que no haya
placa
s de
Peyer
Recto
157
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Va desde el colon sigmoide hasta el diafragma pélvico, siendo curvo y vacío. Tiene una
mucosa más gruesa, con criptas más largas (0,7mm). En su parte más superior se
observan pliegues rectales transversos, y en la inferior hay una zona dilatada, que forma
la ampolla rectal, con pliegues semilunares.
Conducto anal
Se inicia en el diafragma pélvico y llega al orificio anal (4cm). La parte superior
muestra columnas anales o de Morgagni (8-10 pliegues longitudinales). En la base de
las columnas hay unos pliegues de la mucosa, las válvulas anales. Existen unas
depresiones intermedias, los senos anales.
Los senos y las válvulas impiden fugas. Todos estos elementos (válvulas, senos
y columenas) se aplanan durante la distensión. La línea pectínea una la base de las
columnas, corresponde a la membrana anal fetal. Representa la línea divisoria entre la
parte ectodérmica y la parte endodérmica del canal anal. Hay un diferente drenaje
venoso y linfático y distinta inervación.
Según el epitelio se distinguen tres zonas:
x Tercio superior, colorrectal: epitelio prismático simple, similar al recto.
x Tercio medio, de transición: epitelio prismático estratificado.
x Tercio inferior, pavimentosa: epitelio plano estratificado, en continuidad con la
piel perianal.
La muscularis mucosa se inicia en el esófago y es continua hasta llegar aquí, donde
desaparece. Tampoco hay criptas de Lieberkuhn, desaparecen 2 cm antes del conducto
anal.
158
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Las glándulas anales son
tubulares rectas ramificadas,
y se encuentran en la
submucosa. Se extienden
por el esfínter anal interno.
Su secreción sale por un
conducto de epitelio
prismático estratificado que
abre a los senos. Son
glándulas mucosas, que
ayudan al vaciamiento del
canal anal.
Las glándulas pueden llevar
a tendencia a infecciones y formarse abscesos perianales y fístulas.
En la porción distal la piel contiene folículos pilosos, glándulas sebáceas y glándulas
perianales o circumanales. Entre la lámina propia y la submucosa hay ramas de la
arteria rectal superior y el plexo venoso rectal, los glomérulos vasculares. Hay un
cuerpo cavernoso rectal que ocluye la salida de las heces.
La estasis excesiva produce hemorroides internas, por ejemplo debido a hipertensión
portal. Las hemorroides externas se producen por estasis de venas cutáneas en el esfínter
rectal.
No hay tenias, pero la capa muscular interna puede engrosarse para formar el esfínter
anal interno, cuya contracción cierra el conducto anal. El esfínter anal externo es un
anillo de músculo esquelético del periné.
Cuando se distiende por material fecal:
x Columnas, senos y válvulas se aplanan.
x Secreción mucosa de glándulas anales.
x Salida de heces.
Carcinoma colorrectal
x Cáncer de colon hereditario poliposo (HNPCC): se debe a mutaciones en genes
implicados en la reparación de errores de ADEN
x Muchos cánceres de colon se originan a partir de un `´olipo, masa tumoral que
protruye hacia la luz. En los síndromes de poliposis familiares (poliposis
adenomatosa familiar, síndrome....
159
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
ANEJOS AL DIGESTIVO
GLÁNDULAS SALIVALES
Las glándulas se dividen en mayores y menores. Su estructura histológica está
formada por acinos y vías.
Glándulas menores
Son glándulas labiales, yugales, molares, linguales y palatinas. Se localizan en la
mucosa de la cavidad oral y abren a ella. Su secreción es continua y se encargan de
humedecer y lubrificar la cavidad oral.
Glándulas mayores
Son parótida, submaxilar y sublingual. Son grandes con largos conductos
excretores. Su secreción llega a distancia, incluso fuera de la cavidad oral, en respuesta
a estímulos mecánicos y químicos. Son tubuloaacinares compuesta.
Estructura histológica
Hay un estroma, mesenquimal, formado por cápsula y tabiques conjuntivos. El
parénquima es epitelial y forma sialonas, cada sialona está formada por un adenómero y
un sistema de conductos.
Los ácinos están formados por células secretoras se disponen alrededor de una
luz y están rodeadas por células mioepiteliales, Los acinos pueden ser serosos
(circulares, secreción de proteínas), mucosos (esferoidales, secreción de mucinas) o
mixtos (ambas). Las semilunas de Gianuzzi son artefactos debidos a fijaciones
convencionales que en preparaciones con criostato no aparecen; indican la presencia de
ácinos mixtos.
Los acinos segregan una
saliva primaria isotónica, con sIgA,
segregada por plasmáticas. Las
células mioepiteliales son estrelladas
y se encuentran entre las secretoras y
la lámina basal, impulsando gracias a
su capacidad contráctil la saliva hacia
las vías excretoras.
Las vías de salida son:
160
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Conducto intercalar o de Böll (intralobular): epitelio cúbico simple, sirve de
reservorio de células secretoras. Posee células mioepiteliales. Son largos en
glándulas serosas y mínimos en glándulas mucosas (poco identificables).
Conducen la saliva principal, pero secretan HCO3- y absorben Cl-, siendo
isotónica.
x Conducto estriado: epitelio prismático simple, con estriaciones basales
(invaginaciones de membrana) y abundantes mitocondrias. Es mínimo en
glándulas mucosas.
Hay una modificación de la saliva a hipotónica, regulada por la aldosterona:
o Secreción de lisozima, EGF, NGF,
o Secreción de calicreína (proteasa)
o Secreción de K+ y HCO3-
o Reabsorción de Na+ y Cl- (transporte activo)
o sIgA
161
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Conducto interlobulillar Epitelio pasa de cúbico a prismático y en las
x Conducto interlobular glándulas mayores se hace biestratificado.
x Conducto principal
La saliva es un líquido viscoso e incoloro, secretado principalmente por las
glándulas submandibulares (70%) y, en menor medida (25%), por la parótida.
Está formada por agua, α-amilasa, lisozima, peroxidasa, lactoferrina, proteínas
ricas en prol, cistatinas, histatinas, sIgA, IgG, IgM, glucoproteínas ricas en ác. siálico y
sulfato, glucosa, urea, ac. úrico, creatinina, colesterol, cAMP Los electrolitos, en
relación a la sangre, son: x7K, 1/10 Na y x3 HCO3.
Hay un control nervioso, de forma que el SNS estimula la saliva proteica y el
SNPS la saliva acuosa. El agua y el moco se van a encargar de;
x Humedecer la cavidad oral (desecación).
x Humedecer los alimentos secos (deglución).
x Lubrifica el bolo alimentario (deglución).
x Disolver alimentos (estimulación sensitiva).
La alpha-amilasa (ptialina) y la lipasa inician la digestión de azúcares y lípidos.
La lisozima (muramidasa, rompe pared bacterina), lactoferrina (quela, secuestra Fe),
sIgA e histatinas van a controlar la flora bacteriana y levaduras de la cavidad oral. Las
mucinas ricas en ácido siálico y sulfato forman una película adquirida sobre dientes, lo
que regula la unión de bacterias a la mucosa oral. El calcio y el fósforo desarrollan y
mantienen los dientes gracias a su mineralización tras la erupción y a la reparación de
manchas en el esmalte.
Los pacientes irradiados y tratados con fármacos anticolinérgicos tienen caries
generalizadas.
162
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Parótidas
Son pares, extendiéndose del arco cigomático al ángulo mandibular. Son
glándulas serosas, por lo que secretan proteínas, que son ricas en prolina, amilasa,
peroxidasa, lisozima, cistatinas e histatinas. Su estroma es rico con adipocitos
abundantes. Hay conductos intercalares abundantes y largos y estriados, relativamente
contos. El conducto de Sternsen se abre frente al 2º molar superior.
La parotiditis se produce por el virus de la rabia, orquitis, meningitis. Los
tumores benignos requieren cirugía con cuidado de no dañar al nervio facial, que
atraviesa la glándula.
Submaxilares
Pares, situadas entre la mandíbula y el suelo de la boca. Es mixta, con acinos
serosos y seromucosos, predominando el componente seroso. Secretan mucinas muy
glucosiladas ricas en ácido siálico y sulfato, que forman una película protectora sobre
tejidos duros. Van a modular la unión de bacterias con la mucosa oral.
Sus conductos intercalares son cortos, mientras que los estriados son abundantes
y largos. El conducto de Wharton se abre al lado del frenillo lingual.
163
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Sublinguales
Son pares y se sitúan en la mucosa del suelo de la boca cerca de la sínfisis
mandibular. Su estroma carece de cápsula completa y es de tipo mixto, con predominio
mucoso. Sus células mucosas segregan mucinas muy glucosiladas que forman una
película protectora.
Sus conductos intercalares y estriados son mínimos. Las grandes van a abrir al
conducto de Wharton, mientras que las menores lo hacen directamente en la plica
sublingual.
164
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
PÁNCREAS EXOCRINO
Generalidades
El páncreas es una glándula aneja al digestivo situada en la cavidad abdominal,
junto a la 2ª y 3ª vértebras lumbares. Pesa entre 65 y 160 g y mide unos 20-25 cm.
Anatómicamente, podemos dividirlo en cabeza (adherida a la parte media del duodeno),
cuello, cuerpo y cola, extendiéndose estos dos últimos hacia el bazo.
El páncreas es una glándula mixta, ya que combina la secreción endocrina y
exocrina:
x ExocrinaÆ secreción del jugo pancreático (enzimas digestivas).
x EndocrinasÆ secreción de hormonas, que básicamente tienen una función en la
regulación del nivel de glucemia.
Desarrollo embrionario
El páncreas solo se desarrolla en aves y mamíferos, en vertebrados inferiores su
función está desempeñada por células que no se reúnen en un único órgano. Procede de
dos divertículos del epitelio duodenal, cada uno con su propia irrigación, que van a ir
convergiendo hasta formar el páncreas.
Patología: páncreas dividido, los esbozos pancreáticos dorsal y ventral no llegan
a unirse, por lo que es estos individuos el páncreas está separado. El flujo principal
drena en este caso por el conducto accesorio de Santorini en vez de por el conducto de
Wirsung, que es lo normal.
Durante el desarrollo, los ácinos de la porción secretora se ahuecan, y también
posteriormente se forman las vías de excreción en la porción exocrina. En la porción
endocrina las células no se ahuecan, sino que forman cúmulos compactos que quedan
dentro de la porción exocrina, de manera que su conjunto representa tan solo el 1-2% de
la masa celular total.
165
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Estructura histológica
Como todos los órganos, el páncreas se divide en estroma y parénquima
Estroma
El páncreas no presenta cápsula,
aunque sí dispone de una envuelta
conjuntiva muy fina y laxa, cubierta por
peritoneo. La lobulación interior del
órgano es muy incompleta, porque el
estroma es mínimo. Es muy frecuente
encontrar corpúsculos de Pacini.
Parénquima
Es el divisible en porción
endocrina y porción exocrina (explicamos
solo la exocrina, los islotes de Langerhans
no son temario todavía).
Porción exocrina
Las células secretoras forman adenómeros
de secreción serosa. Estas células son piramidales
y tienen un núcleo central o basal no aplanado. Al
agruparse para constituir los ácinos, se unen
mediante uniones occludens y adherens,
delimitando espacios con luz estrecha. Estos
ácinos no presentan células mioepiteliales.
Las células secretoras tienen un citoplasma muy basófilo en su zona basal
debido a la presencia de RER, y el Golgi y los gránulos eosinófilos (gránulos de
cimógeno, más abundantes durante el ayuno y electrodensos) se sitúan en la porción
apical.
La porción exocrina segrega más o menos 1L/diario, como la cantidad de bilis
segregada por el hígado, de jugo pancreático, el cual no es más que el conjunto de unas
20 enzimas encargadas de la digestión de lípidos fundamentalmente, proteínas, ácidos
nucleicos y azúcares. Algunas de ellas son: tripsinógeno, quimotripsinógeno,
procarboxipeptidasa, proaminopeptidasa, elastasa, alfa amilasa, lipasas, colesterolasa,
fosfolipasa, desoxirribonucleasa, ribonucleasa… La concentración de los distintos
componentes enzimáticos varía según la dieta.
Buena parte de las enzimas mencionadas son proenzimas o enzimas inactivos,
las activas son la tripsina, la quimotripsina, carboxipeptidasas, etc. Las proenzimas son
activadas en el duodeno por acción de las enterocinasas situadas en su chapa estriada.
166
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Se calcula que existen en torno a 1’5-3 L diarios del jugo pancreático (esto no lo
entiendo, lo he revisado pero se contradice con lo de que solo hay un litro).
Vías excretoras
Células centroacinares
Las vías excretoras comienzan ya en el ácino, puesto que en él existen unas
células que forman parte del sistema de excreción: las células centroacinares. Estas
células son pálidas y aplanadas, apenas tienen orgánulos y se encuentran unidas entre sí
y con las células secretoras. Dan a la luz, forman un revestimiento interior continuo con
el conducto intercalar.
Las células centroacinares son muy importantes porque segregan el inhibidor de
la tripsina, fundamental para asegurarnos de que las enzimas proteolíticas no se activan
en la luz acinar, ya que si lo hiciesen esto sería muy perjudicial.
Conducto intercalar
Epitelio cúbico simple sin conjuntivo. Las células intercalares contribuyen a la
secreción del jugo pancreático, ya que secretan líquido rico en iones HCO3 - y Na+ capaz
de neutralizar el pH ácido del quimo, lo cual permite activar las enzimas del jugo
pancreático.
Conducto intralobulillar
Epitelio cúbico simple sin conjuntivo
Conducto interlobulillar
Epitelio prismático simple con caliciformes y endocrinas
Conductos principales
Formados por un epitelio prismático simple más alto en el que ya encontramos
tejido conjuntivo. Tenemos dos conductos principales:
167
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Conducto mayor o de Wirsung: se extiende de cabeza a cola del páncreas y va
recibiendo a lo largo de su longitud los conductos menores, de manera que su
aspecto recuerda al de una raspa de pescado. Finalmente se une al conducto
colédoco y desembocan en la carúncula mayor del duodeno. Es importante
recordar la presencia de la ampolla de Vater o hepatopancreática y del esfínter
de Oddi, que regulan el paso de la bilis y el jugo pancreático y por tanto impiden
un reflujo intestinal.
x Conducto accesorio de Santorini: recoge el jugo pancreático formado en la
cabeza y llega al duodeno abriéndose en la carúncula menor.
*Fijémonos en que en el páncreas no existe conducto estriado.
Patologías
Pancreatitis aguda hemorrágica
Propia de individuos que tienen ingestas excesivas o consumo prolongado de
etanol. Es frecuéntela activación prematura de las enzimas o una inactivación del
inhibidor dela tripsina, llevando en ambos casos a una autodigestión del páncreas por
los propios jugos pancreáticos.
Fibrosis quística
No afecta solo al páncreas, pero los quistes se forman en él. Supone la
obstrucción de las vías respiratorias, digestivas y genitales y una secreción anómala por
parte de las glándulas sudoríparas.
Pancreatitis crónica
Lleva a la pérdida de los acinos secretores, a la dilatación de las vías y a una
formación de quistes y una fibrosis. Por lo tanto estos individuos presentan una mala
absorción de grasas y proteínas, y sus heces serán voluminosas y grasas.
168
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
HÍGADO
Generalidades
El hígado es la mayor glándula del cuerpo humano, y la víscera más voluminosa
(1’5 kg). Presenta una superficie superior convexa que se adapta al diafragma y una
superficie inferior cóncava en contacto con el estómago, el riñón derecho y el ángulo
hepático del colon.
El hígado está dividido anatómicamente por surcos en dos lóbulos principales, el
derecho y el izquierdo, más grandes, y en otros dos más pequeños, denominados lóbulo
cuadrado y lóbulo caudado. Presenta como muchos órganos un hilio por el cual penetra
el pedículo hepático, constituido por la arteria hepática, la vena porta y el conducto
colédoco.
El hígado es una glándula mixta o anficrina, pero no presenta una parte
endocrina y otra exocrina claramente diferenciadas como el páncreas, sino que los
hepatocitos que lo componen llevan a cabo funciones tanto exocrinas como endocrinas.
Funciones
A grandes rasgos, sus funciones básicas son:
x Surge como órgano de reserva de moléculas energéticas
x Secreción de bilis en mamíferos
x Órgano central del metabolismo corporal
Vamos a comentar más en profundidad las funciones del hígado:
1. Captar, almacenar y distribuir sustancias nutritivas: hepatocitos
x Mantenimiento de la glucemia: los hepatocitos almacenan glucógeno (de ahí que
sean positivos a PAS), que es degradado por procesos de glucogenolisis en
situaciones de ayuno para formar glucosa, molécula que el hígado también
puede sintetizar a partir de aminoácidos por el proceso de gluconeogénesis.
x Captación de AG para su beta-oxidación y la formación de cuerpos cetónicosÆ
cetosis en diabetes y desnutrición
x Metabolismo lipídico: Síntesis de colesterol captado de la sangre y sales biliares,
parte esencial de la bilis, LDL, HDL, VLDL (en el caso del LDL, HDL y
VLDL, el hígado se encarga no solo de su síntesis sino también del control de
sus niveles circulantes).
x Síntesis urea y contribución por tanto a la eliminación de los iones amonio
generados por ejemplo como consecuencia del metabolismo de bacterias del
tubo digestivo. Los niveles elevados de amonio pueden inducir un coma
hepático.
x Síntesis de todos los aminoácidos esenciales, síntesis de aminoácidos no
esenciales y conversión de aminoácidos no esenciales en esenciales.
169
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
2. Almacenamiento de ciertas reservas de vitaminas para evitar déficits, sobre todo
vitamina A (10 meses), B12 (1 año) y D (en este punto hay cierta controversia,
porque muchos autores defienden que lo que se almacena es un precursor, 4
meses).
x Captación, almacenamiento y mantenimiento de niveles circulantes de vitamina
A: esta vitamina es un precursor del retinal, por lo que tiene una importancia
clave en la visión, motivo por el cual su déficit puede conducir a ceguera
nocturna además de a trastornos de la piel. Esta vitamina se almacena en las
células de Ito en forma de retinol unido a RBP.
x Captación, almacenamiento y mantenimiento de niveles circulantes de vitamina
K: la vitamina K participa a su vez en la síntesis de protrombina y factores de
coagulación, por lo que su déficit conduce a hipoprotrombinemia y trastornos
hemorrágicos. La vitamina circulante viaja unida o bien a quilomicrones o a
LDL.
x Metabolismo de vitamina D, Ca y P: estas tres moléculas son esenciales para la
osificación, por lo que su déficit desencadena raquitismo y trastornos de
mineralización. El hígado no almacena vitamina D, sólo la metaboliza,
realizando la conversión de vitamina D3 en 25 hidroxicolecalciferol.
3. Almacenamiento, metabolismo y homeostasis Fe: el hígado es esencial para
almacenar y metabolizar el hierro. En él se sintetizan proteínas que intervienen
en el transporte y el metabolismo férrico, como es el caso de las transferrinas,
haptoglobinas y hemopexinas. Además, el hígado actúa como almacén de Fe,
que se reserva en forma de ferritina o como gránulos de hemosiderina. Por tanto,
los niveles de hierro en sangre se van a hallar controlados por el hígado. En
personas que han sido trasfundidas repetidamente, el exceso de hierro circulante
puede dañar el hígado, situación que se conoce como hemocromatosis en el caso
de que el exceso de hierro se traduzca en un exceso de hemosiderina.
4. Síntesis de proteínas plasmáticas circulantes:
x Mantenimiento de la presión coloidosmótica y por tanto control del volumen de
plasma y del equilibrio hídrico: algunas de las proteínas que contribuyen a ello
son las globulinas no inmunes alfa y beta y, fundamentalmente, la albúmina.
x Transporte: las proteínas sintetizadas por el hígado se encargan de esta función.
Así, ya hemos dicho que a este nivel se produce la síntesis de lipoproteínas
como HDL, LDL y VLDL. También van a sintetizarse transferrinas,
haptoglobinas y hemopexinas para el transporte del Fe. Se va a formar asimismo
albúmina, que se encarga de transportar otras proteínas y AG.
x Cascada de la coagulación: síntesis de protrombina y fibrinógeno gracias a la
vitamina K
5. Inmunológica
x Síntesis de complejos Ig A- complejo secretor. Sabemos que esta Ig se genera en
su mayor parte en el tubo digestivo, y es capaz de retornar a la circulación
sanguínea para ir al hígado y volver a ser segregada por los hepatocitos, ya que
estos son capaces de sintetizar el complejo secretor, como parte de la bilis.
170
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Fagocítica: las células de Kuppfer presentan receptores fagocíticos y del
complemento. Gracias a ello, reconocen y eliminan a un 90% de las bacterias
que llegan al hígado a través de la porta, fagocitan células muertas y contribuyen
a la eliminación de eritrocitos
6. Degradación de proteínas
x Degradación de Hb: la bilirrubina se genera en el bazo a partir de la Hb
(destrucción eritrocitaria en bazo e hígado) y se une a albúmina, circula y llega
al hígado. Aquí, la bilirrubina es conjugada para que pueda ser eliminada por la
orina y en una pequeña proporción por las heces (bilis). Cuando esta función no
se realiza bien hay ictericia en piel y mucosas, signo de hepatitis, provocada por
altos niveles de bilirrubina libre o conjugada. Puede tener también una causa
hemolítica, infecciosa u obstructiva.
7. Degradación de fármacos, toxinas y proteínas extrañas: es una de las funciones
más importantes de los hepatocitos, que degradan moléculas no hidrosolubles y
por tanto no eliminables vía orina, que de no ser así se acabarían acumulando en
el organismo causando intoxicación. Se requiere una doble reacción:
1. Oxidación: en el REL, con participación de mitocondrias. Interviene el
citocromo P450.
2. Conjugación: en los peroxisomas. La molécula oxidada se une a ácido
glucurónico, taurina o glicina. Convierte la molécula en hidrosoluble y
eliminable.
La tolerancia a los fármacos se debe a que los hepatocitos se adaptan,
hipertrofiando su REL y aumentando la dotación de enzimas relacionadas con la
oxidación y la conjugación. El consumo sostenido de fármacos, como es el caso
de los barbitúricos, disminuye su eficacia y requiere dosis mayores para que se
generen los efectos que de ellos se esperan.
8. Secreción de bilis: es la secreción exocrina. Tiene función digestiva y de
eliminación de ciertos productos. La función digestiva está relacionada con las
sales biliares que contiene, sintetizadas por el hepatocito. Cuando no hay
suficientes, las grasas no se absorben y se eliminan en las hecesÆ heces grasas.
9. Funciones endocrinas: modifica la acción de hormonas
x Tirosina: convierte T4-tirosina en T3-tirosina, que es la forma activa
x Degrada insulina y glucagón
x Síntesis de 25 hidroxicolecalciferol a partir de vitamina D
x Síntesis de pequeña cantidad de GH
Estructura histológica
Origen embriológico
El hígado presenta un parénquima endodérmico y un estroma mesodérmico. En
el embrión, el hígado se desarrolla como una invaginación endodérmica del intestino
anterior, más concretamente de la zona que posteriormente dará lugar al duodeno, el
cual da lugar al divertículo hepático. El divertículo prolifera y da lugar a los hepatocitos,
171
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
que se organizan en trabéculas de Remak para formar el parénquima del hígado. El
pedículo original del divertículo hepático se convierte en el conducto biliar común. Un
brote de este conducto va a formar el divertículo cístico, que dará lugar al conducto
cístico y la vesícula biliar.
Parénquima
Tiene como célula principal al hepatocito. Los hepatocitos se organizan en
cordones llamados también trabéculas de Remak, que parten de manera radial desde una
vena central, que es una ramificación de la vena hepática. Los cordones constituyen una
red tridimensional, aunque nos sea difícil imaginarlo si no es en un solo plano, y son
paralelos a los sinusoides venosos. Entre la pared del sinusoide y la de los hepatocitos
de los cordones se delimita un espacio denominado espacio de Disse.
Estroma
Es un tejido conjuntivo que se continúa con la cápsula de Glisson. Esta cápsula
es de tejido conjuntivo denso, no es muy gruesa y se halla cubierta por mesotelio. Emite
trabéculas que permiten la formación de lóbulos y lobulillos (dentro de los lobulillos
solo encontramos una red de fibras reticulares, fuera de ellos también podemos ver
colágenas y unas pocas elásticas).
Formando parte del estroma están los espacios portales o de Kiernan, conjuntivo
laxo con colágeno I y reticulares, escasas células, vasos linfáticos y fibras amielínicas
simpáticas y parasimpáticas. Lo que caracteriza a estos espacios es la presencia de
tríadas portales, formadas por ramificaciones de:
x Conducto biliar
x Arteria hepática
x Vena porta
172
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
División del hígado
Lobulillos hepáticos clásicos
El primero en realizar una división del hígado fue Malpigio, que lo dividió en
lobulillos hepáticos: unidades estructurales de contorno poliédrico y forma hexagonal o
pentagonal. En el centro del lobulillo hepático tenemos la vena central, que desde ahí
emite ramificaciones que irrigan las trabéculas, pegadas a los sinusoides. Los espacios
portales se sitúan en la periferia del lobulillo. La limitación periférica del lobulillo la
constituye una placa limitante de hepatocitos solo perforada por las ramificaciones de la
arteria hepática y la vena porta y de los conductos biliares.
Dinámica del lobulillo clásico
La sangre arterial y venosa se mezclan en el sinusoide y circulan hacia la vena
central. La bilis circula en sentido opuesto, centrífugo. Según Malpigio, el lobulillo
hepático clásico es una unidad histológica y funcional, pero no todos los hepatocitos
funcionan ni son iguales. Se establece una zonación. Ello se demuestra cuando se hace
una ingesta, puesto que la glucosa se deposita antes en unas zonas que otras, de forma
concéntrica. Además, los fármacos tampoco afectan al mismo tiempo a todos los
lobulillos.
Como patologías asociadas a los lobulillos, destacamos:
x Necrosis farmacológica del lobulilloÆ toxinas, medicamentos, paracetamol
x Hepatitis crónica activa o persistenteÆ deterioro placa limitante
Lobulillo portal
Fue definido por Mall. En este caso, el espacio delimitado es más pequeño, de
contorno también poliédrico pero esta vez de sección triangular. Los límites periféricos
son las venas centrales, aunque sea contradictorio, y los espacios portales se encuentran
en el centro del lobulillo portal. No tienen una delimitación clara.
Mall entiende que el hígado es una glándula, y por tanto para él el lobulillo
portal es la unidad secretora. En este caso la bilis fluye hacia el centro, ya que el
conducto se sitúa ahí, dentro del espacio portal.
173
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Ácino hepático
El ideólogo de estos espacios fue Rappaport. Es una unidad poliédrica en forma
de romboide sin límites claros, en cuyo eje hay una serie de vasos y conductos, que son
los que van de un espacio de Kirnan a otro. Esto es una concepción más moderna que
permite entender mejor el funcionamiento del hígado con unidades de tipo metabólico
en las que los hepatocitos no funcionan igual tampoco, puesto que también hay
zonación (al igual que en los lobulillos clásicos), de dentro hacia afuera:
x Zona 1: mayor dotación de enzimas para la
gluconeogénesis y el metabolismo oxidativo.
Reciben por este motivo en primer lugar los
nutrientes y el oxígeno, y por ello son los
últimos en verse afectados en caso de una
isquemia. Sin embargo, se afectan en primer
lugar en caso de una intoxicación o estasis
biliar.
x Zona 2
x Zona 3: más carga de enzimas para el
metabolismo de lípidos, tóxicos y glucólisis.
Reaccionan más tarde a la presencia de toxinas,
pero son los primeros en verse afectados por
niveles bajos de O2. Es la más afectada por
insuficiencia cardiaca congestiva o cirrosis
cardiaca, esto es, una afectación del hígado cuyo
origen es una insuficiencia primaria del corazón.
Vascularización
En el hígado hay muchos sinusoides. En estos hay muchas fenestras, brechas y
con ello el plasma es capaz de salir a través de dichos sinusoides. Donde primero se
recoge dicho plasma es en el espacio de Disse.
El espacio de Disse queda entre la pared de las células endoteliales del sinusoide
y la membrana plasmática de los hepatocitos. Estos espacios son continuos y se
encuentran en la zona periférica del lobulillo clásico, es decir, en el espacio portal. El
plasma va de los espacios de Disse al espacio de Mall.
174
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
El espacio de Mall está en la zona periférica, fuera de la membrana limitante. Se
localiza entre el estroma del espacio portal y los hepatocitos más periféricos del
lobulillo.
Tanto en el espacio de Disse como en el de Mall encontramos revestimiento
propio del linfático.
Vascularización linfática
Los primeros capilares linfáticos se sitúan en el espacio portal, luego la linfa
fluye en la misma dirección que la bilis. Drenando al final al conducto torácico. En el
modelo clásico el plasma fluye hacia la periferia.
Los espacios de Disse están llenos de plasma que se extravasa a los espacios de
Mall (más amplios) y, además, en el espacio de Kiernan encontramos también capilares
linfáticos.
Vascularización sanguínea
Un 75% de la sangre que llega al hígado lo hace a través de la vena porta
hepática, con una sangre color rojo oscuro por su gran cantidad de nutrientes (aa,
azúcares, catabolitos de la hemoglobina del bazo, hormonas t.d/pancreáticas). Tiene este
color debido a que recoge los componentes endocrinos de la pared del tubo digestivo y
del páncreas, no sólo nutrientes, sino también hormonas. De esta vena porta hepática se
dan ramificaciones interlobares, conductoras, interlobulillares, hasta llegar a las venas
portas terminales, en los espacios portales.
El 25% de la sangre restante lo lleva la arteria hepática que aporta
fundamentalmente oxígeno (con sangre oxigenada), que también se ramifica también en
ramas interlobares e intralobulares, hasta las ramas terminales. Estas últimas se
localizan en los espacios portales.
Desde los espacios portales la sangre que ha entrado por la vena porta y por la
hepática, al llegar a los sinusoides se mezcla. Estos sinusoides están formados por
células endoteliales, células de Kupffer y células de Ito (llamadas también lipocitos o
células almacenadoras de grasa).
Una vez que la sangre se ha mezclado por los sinusoides, alcanza las venas
centrolobulillares, que convergen entre sí para formar venas sublobares o intercalares,
colectoras y hepáticas, hasta alcanzar la cava inferior.
Este sistema es independiente del sistema venoso portal.
175
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Células Endoteliales
El endotelio está formado por células epiteliales muy delgadas. Este endotelio
corresponde a capilares fenestrados con grandes brechas. Los capilares no se apoyan en
la lámina basal, de modo que es un conducto de pared muy fina por la que se sale la
sangre (tanto venosa como arterial) hacia los espacios de Disse; excepto los elementos
formes y los quilomicrones que quedan retenidos en los sinusoides.
Células de Kupffer
Son células de morfología
estrellada que provienen de macrófagos,
teniendo por tanto capacidad fagocítica.
Derivan de un precursor de la médula
ósea. Este precursor alcanza el hígado y
allí vamos a ver estas células en las
paredes de los sinusoides.
Las podemos encontrar tanto en la
luz como en la pared del sinusoide y no
presentan uniones entre ellas ni con las
células endoteliales. Además, son capaces de emitir pseudópodos y moverse y se
pueden dividir de forma activa (aunque siguen viniendo de la medula ellas pueden
proliferar). Luego no sólo son capaces de renovarse a partir de precursores, sino que
también ellas mismas se multiplican. Se detectan por sales de oro.
Las células de Kupffer tienen muchas funciones:
x Se encargan de degradar eritrocitos (aunque es fundamentalmente en el bazo el
lugar en el que ocurre esta degradación, en el hígado también se produce) por lo
que es fácil encontrar en estas células inclusiones de hierro. Esta función resulta
esencial en individuos que han pasado por una esplenectomía.
x Además, estas células tienen que ver con la función inmunológica del hígado:
segregan citocinas (TNF alfa, TGF beta, interleucina 6) ante la estimulación
producida por ciertos agentes (alcohol, infecciones), produciendo una reacción
inflamatoria, fundamentalmente ante una infección vírica, toxina bacteriana o
consumo tóxico (por ejemplo de etano).
Células de Ito o lpiocitos
Estas células se ven por tinción con sales de oro. Son células estrelladas (aunque
la morfología puede variar) de origen mesenquimal. Se caracterizan por tener
inclusiones con lípidos, entre ellos vitamina A, la cual almacenan y movilizan cuando es
necesaria. En su citoplasma encontramos muchas vacuolas lipídicas.
Se encargan, por tanto, del almacenamiento y control de retinoides. También, en
determinadas situaciones patológicas como cirrosis, las células de Ito se transforman en
miofibroblastos y van a comenzar a sintetizar de forma masiva colágeno
176
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
(fundamentalmente I y III) de manera que el hígado es cada vez más fibroso (fibrosis
hepática). Estas células están en el sinusoide y son contráctiles, se produce una
disminución en el diámetro de los sinusoides (habiendo una disminución en la cantidad
de fenestras de estos vasos). Por tanto, va a acabar causando hipertensión portal de
manera que hace que estas células de ito también puedan regular o controlar el flujo
sanguíneo que hay por los sinusoides.
En pacientes con citocinas más elevadas ocurre que el hígado se hace fibrótico.
Las células de Ito juegan un papel importante cuando, tras una lesión hepática,
comienza una regeneración. Durante esta reparación, las células de Ito se encargan de
restaurar/remodelar la matriz extracelular, no solo sintetizando colágeno I sino otro tipo
de moléculas que forman parte de ella.
Espacio de Disse
Es la primera zona donde encontramos la extravasación de plasma, es un espacio
intercelular que queda entre el endotelio sinusoidal y la membrana plasmática del
hepatocito.
En este espacio casi no hay elementos de matriz, algunas pocas fibras de
reticulina, fibras nerviosas perdidas y algún axón amielínico. Hay sobre todo plasma
que fluye hacia la zona periférica, hacia el espacio de Mall. En este espacio de Disse
encontramos las microvellosidades de las membranas de los hepatocitos.
177
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
El espacio de Disse es la zona en donde se produce la hematopoyesis durante la
etapa fetal. En adultos con anemias crónicas, se ha demostrado que las células
hematopoyéticas de esta zona se recuperan.
Hepatocito
Los hepatocitos son células de origen epitelial, luego están polarizadas. Esta
célula se divide pero no con mucha frecuencia y muy lentamente, pues tienen una vida
media bastante larga de 4-6 meses. El hepatocito tiene una alta capacidad regenerativa:
en situación de daño hepático, esta capacidad se pone de manifiesto y se recupera la
masa hepática rápidamente.
Los hepatocitos son células grandes, con núcleo grande (aunque es típico
encontrar hepatocitos poliploides /polinucleados) y en posición central. Tienen un
citoplasma eosinófilo con todo tipo de orgánulos.
Pueden aparecer inclusiones lipídicas, lo cual es normal, salvo que se acumule
demasiado, lo cual ocurre en el hígado graso (situación patológica no grave pero no
normal). Presenta asimismo inclusiones de glucógeno alfa y beta (PAS+). Esta cantidad
de glucógeno no es una cantidad fija, sino que presenta variación circadiana
(variaciones a lo largo de las 24 horas del día en función de la ingesta).
El hepatocito en su citoplasma presenta abundantes mitocondrias y un gran
desarrollo del citoesqueleto (lo que es normal en una célula epitelial), que forma una red
bastante más densa/abundante en algunas zonas, como por ejemplo en las cercanías de
la superficie canalicular. El hepatocito presenta una gran cantidad de RER (por la gran
cantidad proteínas que sintetizan), que se pone de manifiesto a microscopía óptica como
grumos de Berg. Su aparato de Golgi es muy grande, con hasta 50 dictiosomas (no solo
uno), relacionados con la secreción de bilis y VLDL.
El REL del hepatocito es fundamental en degradación de tóxicos, fármacos,
proteínas no hidrosolubles… (Conjugación). Asimismo, está relacionado con la síntesis
178
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
de colesterol y sales biliares, la degradación de glucógeno, la degradación de ácidos
grasos para formar triglicéridos y la detoxificación. Otra de sus funciones es la
eliminación de yodo y la metabolización del etanol (en el REL) para lo cual existen dos
vías alternativas:
x Con la ADH (enzima
que se posee cuando se
consume alcohol de manera
habitual). Esta actúa sobre el
etanol, obteniéndose protones
y acetaldehído dentro de las
mitocondrias. El acetaldehído
se transforma en acetato y en
esta vía aunque conseguimos
eliminar el etanol obtenemos
protones y acetaldehído que,
cuando se forman en grandes
cantidades, producen daño
hepático: dañan las
mitocondrias y el
citoesqueleto y además
generan una respuesta
autoinmune.
x La segunda alternativa es la vía MEOS (sistema microsomal de oxidación del
etanol). En esta vía, el etanol se metaboliza dando acetaldehído y radicales de
oxígeno, muy lesivos para la membrana plasmática del hepatocito. Como
consecuencia se produce acúmulo de sustancias lesivas y disminución de la
capacidad de desintoxicación. El hepatocito reacciona provocando una reacción
autoinmune y perdiendo su capacidad de resistir los residuos de etanol.
Así, por mucho que consigamos metabolizar el etanol, hay daño hepático.
Clínica
x Hígado graso (reversible)
x Esteatohepatitis: Además de mostrar acumulación de lípidos tenemos
reacción inflamatoria)
x Cirrosis (fibrosis hepática): acumulación de fibras.
x Hepatocarcinoma
Una persona que bebe alcohol crónicamente tiene un hígado graso, cuando en el
hígado ya hay un inicio de respuesta inflamatoria se da esteatohepatitis. Cuando ya hay
signos de que el hígado sea más fibroso da cirrosis y si todo continua y da a más se tiene
hepatocarcinoma.
El REL también tiene que ver con la metabolización del grupo hemo ya sea por
la bilirrubina del bazo como la del intestino. Esta bilirrubina viaja en sangre unida a la
albúmina. En niveles elevados es toxica para el cerebro.
179
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
En el bazo se degrada el grupo hemo de la hemoglobina. En el bazo, el grupo
hemo se transforma en biliverdina y después en bilirrubina. Si esto no ocurre
correctamente, aparece ictericia. La bilirrubina viaja en sangre unida a la albúmina, pero
es tóxica para el cerebro.
En los fetos cuya sangre es incompatible con la de la madre esto puede dar
problemas si se da eritroblastosis fetal, ya que la destrucción de los eritrocitos produce
una hiperbilirrubinemia que puede desembocar en kernícterus, una intoxicación del
SNC por bilirrubina en el feto (daño irreversible en el desarrollo del SNC).
El hepatocito capta la bilirrubina (el complejo albúmina-bilirrubina), que entra al
hepatocito sin albúmina (se queda en el plasma). La bilirrubina entra unida a la
ligandina, viaja al REL y allí la bilirrubina ya libre se conjuga con el ácido glucurónico.
Esta bilirrubina conjugada es la que el hepatocito segrega al canalículo biliar (va a la
superficie canalicular).
La bilirrubina conjugada contenida en la bilis llega al duodeno y, más adelante,
el ácido glucurónico se escinde, teniendo otra vez bilirrubina libre. Parte de esta es
metabolizada por bacterias, formando urobilinógeno, que es excretado. Un 20% de la
bilirrubina es reabsorbida, fundamentalmente en el íleon y el colon, y vuelve al hígado.
Este mecanismo es lo que se conoce como circulación entero-hepática de la bilis.
Una pequeña cantidad de bilirrubina se excreta con la orina como urobilina.
Algunas de las patologías relacionadas con el metabolismo del grupo hemo son:
x Hiperbilirrubinemia
o Enfermedad de Crigler-Najjar: el hepatocito no conjuga la bilirrubina,
provocando encefalopatías porque tenemos niveles elevados de
bilirrubina que altera al SNC y al hígado.
o Enfermedad de Dubin-Johnson: deficiencia en el transporte de la
bilirrubina conjugada, que no es segregada correctamente, el individuo es
incapaz de expulsarla por el canalículo.
Encontramos en el hepatocito una importante cantidad de pigmentos biliares y
lisosomas en los que se acumula Fe, Cu y lipofucsina. La cantidad de estos lisosomas
aumenta en ciertas patologías: anemia, hepatitis, estasis biliar…
Algunas patologías relacionadas con esta función del hígado son:
x Tesaurismosis: exceso de lisosomas con Fe en hepatocitos.
x Hemocromatosis hereditaria: es genética, y consiste en un exceso de Fe en
hepatocitos, que provoca cirrosis y carcinoma. Se acumula hemosiderina.
x Degeneración hepatolenticular o enfermedad de Wilson: es una acumulación
de Cu, que en el hígado provoca daño hepático crónico, que a su vez provoca
cirrosis.
180
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
En el hepatocito también hay peroxisomas. Los peroxisomas son orgánulos
pequeños con funciones fundamentales en el hígado, siendo muy acumulables en él.
Metabolizan ciertos radicales (llevan a la formación del peróxido de hidrógeno que es
muy lesivo) y tóxicos, de modo que son fundamentales en la función de detoxificación,
junto con el REL. El 50% del etanol es detoxificado en los peroxisomas.
Intervienen con su gran cantidad de enzimas en la beta-oxidación, en
gluconeogénesis y en el metabolismo de las purinas.
Existen algunas enfermedades relacionadas con los peroxisomas:
x Enfermedad de Zellweger: consiste en una ausencia de las enzimas de los
peroxisomas, que están vacíos. Es mortal.
x Adenoleucodistrofia: es una enfermedad ligada al cromosoma X, y provoca
la degeneración de la mielina de los nervios.
Superficie Sinusoidal, Intercelular y Canalicular
En la superficie del hepatocito hay diferentes polos.
La zona que da a los sinusoides es el polo basal. Es la zona mayoritaria y donde
se da el intercambio de moléculas con la sangre. Hay microvellosidades, pero no hay
lámina basal.
En el hígado encontramos los hepatocitos adosados en forma de cordones. La
mayor parte de la membrana plasmática de los hepatocitos da al espacio de Disse,
formando la superficie sinusoidal hepática (70%). Esta membrana tiene abundantes
microvellosidades, lo que permite aumentar la superficie de intercambio con la sangre.
Necesitamos mucha superficie porque el hepatocito está continuamente cogiendo
productos de la sangre y aportando otros (redistribuyendo).
181
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
El resto de la membrana plasmática se divide entre la superficie intercelular
(15%), adosada a otro hepatocito, regular y continua, con uniones de comunicación (no
son las únicas uniones pero son las más llamativas); y la superficie canalicular (15%),
que es un espacio delimitando los canalículos biliares que se abre entre los hepatocitos
(es la membrana por la que se excreta la bilis que está junto a un espacio que se abre
entre hepatocitos denominado canalículo de 0,5 μm – 1,5 μm).
El canalículo es un hueco que se abre entre dos o tres hepatocitos y a donde se
secreta la bilis. Estos canalículos tienen una anchura de 1μm y muestran
microvellosidades que no son tan abundantes.
Encontramos abundantes fosfatasas alcalinas que se encargan de intercambiar
productos y excretar la bilis (consume mucha energía), puesto que el canalículo es la
primera vía de la secreción biliar. Esta membrana plasmática tiene una serie de
transportadores: son cuatro de la familia ABC (que consumen energía), encargados del
transporte específico de moléculas. MDR1 transporta colesterol; MDR2, fosfolípidos;
MOAT, bilirrubina conjugada; y BAT, sales biliares.
Es fácil encontrar mutaciones en los genes de estos transportadores, afectando a
la secreción de la bilis (la bilis no tiene el componente que transporta ese transportador)
y además cursaría con necrosis que no sería generalizada sino focal (en puntos donde
los transportadores no funcionan). Estas necrosis focales provocan la proliferación de
los conductillos biliares e inflamación.
En los hepatocitos se crea un gradiente osmótico de ácidos biliares, por lo que el
hepatocito tiende a expulsar agua al canalículo biliar, lo que permite el paso de
bicarbonato y cloro al mismo (hay canales para el cloro y para el bicarbonato) y en la
luz del canalículo, asociada a la membrana plasmática existen unas enzimas
denominadas ectoenzimas.
182
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Además, en cualquier secreción necesitamos que el canalículo esté sellado (si no
la bilis fluiría hacia la superficie intracelular) por lo que hay mayor trama de
citoesqueleto y uniones ocluddens y adherens (a las que van filamentos de actina
pudiendo haber una contracción del citoesquelto causando que el diámetro del
canalículo cambie, aumentando y disminuyendo, 0.5-1μm). Así, si conseguimos
disminuir el diámetro influimos en el avance de la bilis
La Amanita phalloides inhibe este sistema contráctil inactivando los
microfilamentos.
Bilis
Compuesta por agua, Na+, K+, Ca2+, Mg2+, Cl-, HCO3-, fosfolípidos, colesterol,
ácidos biliares primarios y secundarios, pigmentos biliares y bilirrubina. Funciones:
x Excreción de colesterol, fosfolípidos, sales biliares, bilirrubina conjugada, y
electrolitos.
x Excreción de catabolitos de metales pesados y fármacos procesados.
x Facilita la absorción de las grasas al emulsionarlas
x Transporte de IgA a la mucosa intestinal (circulación enterohepática)
x Los ácidos biliares conjugados impiden el crecimiento bacteriano en el intestino.
Árbol biliar
Vías de calibre creciente. Se dirigen de los hepatocitos a la vesícula biliar y
finalmente al intestino.
Vías intrahepáticas
x Canalículos o capilares biliares: Espacios
entre hepatocitos adyacentes, con
microvellosidades de la membrana del
hepatocito. Poseen fosfatasa alcalina (puede
causar obstrucción de las vías biliares) y
zonula occludens, que forma un anillo
completo. El flujo es centrífugo, cerca del
espacio portal se reúnen (colangiolos).
x Conductillos biliares, colangiolos o conductos de Hering: Dentro del lobulillo
hepático hasta el límite, perforando la placa limitante. Son cortos, de 1-1,5
micras. Su epitelio es cúbico simple, con lámina basal.
x Conductos biliares interlobullillares: Está en la triada portal, con un diámetro de
15-40 micras. El epitelio evoluciona de cúbico a prismático simple.
Encontramos microvellosidades, fibras elásticas en el conjuntivo denso y fibras
musculares cerca del hilio.
x Conductos hepáticos derecho e izquierdo
183
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Conducto hepático común: Mide 3 cm, con epitelio prismático simple bajo el
que se encuentran las mismas capas que en el tubo digestivo, a excepción de la
muscularis mucosa.
x Conducto cístico: Nace de la vesícula biliar, con las mismas capas. Su mucosa
está plegada en espiral formando la válvula espiral o de Heiser. Controla la
entrada o salida de bilis de la vesícula.
x Conducto colédoco: Mide 7 cm. Se une con el conducto de Wirsung, con el que
va a desembocar por la carnúcula mayor. Se una dilatación, la ampolla de Vater
y su excreción está controlada por el esfínter de Oddi, formado por:
o Esfínter de Boyden: engrosamiento de la muscular que retine la bilis en
la vesícula
o Esfínter pancreático: inhibe la expulsión del jugo pancreático e impide el
paso de la bilis al conducto pancréatico.
o Esfínter de la ampolla: impide que la bilis y el jugo pancreático se dirijan
al tubo digestivo.
o Fascículos longitudinales: entrada de la bilis y el jugo pancreático al tubo
digestivo.
x Ampolla de Vater
El flujo de bilis depende de la presión secretora generada por los hepatocitos y el
cierre del esfínter de Oddi. En el ayuno el esfínter está cerrado y la bilis permanece en la
vesícula. La digestión induce la salida de CCK, que estimula la contracción de la
vesícula y la relajación del esfínter de Oddi, lo que produce que la bilis se dirija al
duodeno.
Clínica
x Colestasis: alteraciones en la formación y excreción de la bilis. Puede ser
intrahepática (hepatocito) o extrahepática (vías). La causa puede ser estructural o
mecánica (colelitiasis).
x Colelitiasis: Concentraciones sólidas de bilis sobre las que se depositan sales de
calcio. El 70% se debe al colesterol. Presencia de sales de calcio, ácidos biliares,
pigmentos biliares, proteínas, ácidos grasos y fosfolípidos. Hay una calcificación
del pigmento: bilirrubinato de calcio.
Los factores de riesgo para su desarrollo son la obesidad, los trastornos
alimentarios, la pérdida rápida y exagerada de peso y los estrógenos.
x Colecistitis: Colelitiasis del conducto cístico. Afecta a la vesícula biliar
produciendo mayor contracción, desarrollo muscular, invaginaciones de mucosa
y riesgo de infección. Da lugar a dolor y fiebre. Da lugar a un cólico biliar.
x Ictericia obstructiva: Provoca retención biliar, paso de la bilis hacia los
sinusoides, cambios en capilares biliares, alteración de digestión de grasas y
heces pálidas.
x Infección de las vías:
o Tremátodos
o Inflamación, abscesos y dificultad en el flujo de la bilis.
184
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
VESÍCULA BILIAR
Es un órgano hueco y alargado de dimensiones variables, cercanas a 4x10cm,
con una capacidad de 50-70 mL. Se divide en fondo, cuerpo y cuello, el cual se continúa
con el conducto cístico. Se drenan 600-1000 mL/día de bilis.
Posee una evanginación espiral de la mucosa del cuello, la válvula espiral de
Heister, que evita la distensión o el colapso cuando hay cambios bruscos de presión.
Se encarga de:
x Almacenar la bilis
x Concentrar la bilis (x10)
x Liberación en respuesta de alimentos
x Regulación de la presión hidrostática del árbol biliar.
Estructura
Mucosa
Su borde es irregular, formada por pliegues o arrugas paralelos con muchas
prolongaciones. Está cubierta de un moco protector. Los pliegues son muy variables y
según la contracción de la mucosa son:
x Contraída: pliegues altos, numerosos y paralelos.
x Distendida: pliegues cortos, irregulares y separados.
También hay invaginaciones o divertículos, conocidos como senos de
Rokitansky-Aschoff. Están formadas por un epitelio similar al de revestimiento y se
sitúan en el cuello, penetrando hasta la muscular. No deben confundirse con glándulas.
Se ha discutido que si aumenta su número en situaciones patológicas. La colecistitis
crónica es una acumulación de bacterias que se sitúa en estos senos.
El epitelio es prismático simple formado por células principales y endocrinas,
sin caliciformes. Las células principales poseen abundantes microvellosidades (PAS+),
ATPasa Na+/K+, complejos de unión, su núcleo es ovalado y basal, con citoplasma
eosinófilo y rico en mitocondrias (basales y apicales). Sus funciones son:
185
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Absorbe agua, Na+, Cl- y HCO3- y los secreta por la membrana lateral.
Hay un gradiente osmótico desde la luz/citoplasma hasta el espacio intercelular.
El aumento de la presión hidrostática hace que la secreción llegue a la lámina
propia y finalmente hasta la sangre. Es similar a enterocitos del intestino delgado
y del colon.
x Vesículas de secreción (glucoproteínas), que van a formar una película
protectora.
La lámina propia es el conjuntivo, similar al del colon, con abundantes linfocitos
y plasmocitos. Su vascularización es rica, con capilares fenestrados, aunque con
ausencia de linfáticos. A nivel del cuello van a tener glándulas túbuloalveolares
mucosas, son particularmente abundantes durante la inflamación.
Muscular
Red irregular de miocitos lisos oblicuos, longitudinales y transversales con fibras
elásticas y colágenas. La disposición es helicoidal. Su contracción reduce el volumen
haciendo que se expulse la bilis, regulada por CCK y Ach.
Adventicia/serosa
Gran grosor, libre. Es una capa gruesa de conjuntivo denso con fibras colágenas
y elásticas y abundantes adipocitos. Está en continuidad con el conjuntivo interlobular
hepático.
Hay grandes vasos y un plexo linfático que recoge la linfa del hígado. Hay
nervios y glanglios parasimpático, incluso conductos biliares aberrantes (de Luschka).
Secreción biliar
El hígado segrega la bilis de forma continua, pero llega al intestino de forma
intermitente. El almacenamiento de la bilis en la vesícula va a depender de CCK (grasa
en el intestino) y de la inervación parasimpática (nervio vago).
La CCK va a provocar contracciones peristálticas en el músculo de la vesícula
biliar, y a su vez relaja el músculo intestinal y el esfínter de Oddi, haciendo que el flujo
de bilis sea intermitente. La liberación parasimpática produce un aumento de la
liberación de las enzimas y la bilis, pero no del volumen; se pueden administrar
fármacos colinérgicos.
186
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
APARATO URINARIO
FUNCIONES
x Producción de la orina
x Permite la evacuación del semen
x Control de la homeostasis del agua y las sales Filtración,
x Control del volumen y osmolaridad del líquido extracelular excreción y
x Control del equilibrio ácido-base absorción.
x Síntesis y secreción de hormonas y factores: eritropoyerina, renina, activación de
1,25 dihidroxicalciferol
x Control de la tensión arterial
x Participa en el metabolismo: eliminación de catabolitos, productos extraños y
formación de orina. Se utilizan para ello mecanismos de filtración y excreción.
RIÑÓN
Par, en el retroperitoneo. Forma de haba, con un hilio rico en grasa. Consumen
una gran cantidad de energía (20% del gasto cardiaco) para la filtración de la sangre
(125 mL/min). Se producen 189 mL al día de orina primaria (ultrafiltrado). En las
nefronas se reabsorbe el 99% de la orina, quedando 1,5 L.
Vascularización
La arteria renal se encuentra en el hilio. Se dividen en las arterias interlobulares,
que ascienden por columnas de Bertin. Las arterias arciformes se originan en la unión
cortico-medular en dirección horizontal y dan lugar a las arterias interlobulillares, que
ascienden por la corteza hasta la cápsula, siendo arterias terminales sin anastomosis, de
forma que su obstrucción produce necrosis renal. Esta sangre puede pasar a:
x Arteriola aferente: entra al corpúsculo renal, donde se capilariza formando el
glomérulo. Da ramas para túbulos corticales.
x Arteriola eferente: la sangre que sale pasa a una red capilar peritubular
fenestrado, descendente. Irriga los túbulos de las nefronas y conductos
colectores de corteza, por lo que es un vaso porta. Va a drenan en las venas
interlobulillares y vasos arciformes.
187
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
En la corteza la sangre circula por vasos rectos, que alcanzan la unión cortico-
medular, desde donde pasan a las venas arciformes que drenan a las venas
interlobulillares. Tras ello la sangre va a las venas interlobares y sale por la vena renal.
188
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Estructura
x Estroma: Cápsula e intersticio
x Parénquima: elementos epiteliales (corteza y médula).
Cápsula
Completa. Se introduce en el hilio, formando la cubierta del seno. En el exterior
hay fibras colágenas y fibroblastos y en el interior miofibroblastos. Esta estructura
permite adaptabilidad.
Corteza
Contiene el 90-95% de la sangre del riñón, por lo que es rojiza. Rodea por
completo a la médula. Contiene corpúsculos de Malpigio, de granulosa. Hay
estriaciones verticales, los rayos medulares de Ferrein, que pueden llegar a 500.
Médula
Pálida y estriada, sin corpúsculos y de doble espesor. Contiene 8-18 pirámides
de Malpigio separadas por tejido cortical (columnas de Bertin). En la base está cubierta
por corteza y forma una papila hacia el seno renal. Se puede dividir en médula externa
(con franja externa e interna) y médula interna.
Lóbulo y lobulillo
El lóbulo renal corresponde a una pirámide renal más la corteza por la que está
rodeada. En el conejo y la rata solo hay un lóbulo. El hombre es multilobular, siendo los
lóbulos solo visibles hasta el primer año.
Un lobulillo renal es de estructura cortical, definida de dos maneras:
x Parte de corteza flanqueada por dos arterias interlobulillares ascendentes.
x Parte de corteza con un conducto colector y las nefronas que drenan en él.
Cada lobulillo contiene porciones
rectas de las nefronas y el conducto
colector que forma un rayo medular, eje
del lobulillo. Las porciones adyacentes
contienen los laberintos corticales.
189
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Túbulo urinario
El túbulo urinario es la unidad renal, y está constituido por:
x Nefrona: de origen metanéfrico, hay unas 2000000 por riñón.
x Tubo colector: se desarrolla a partir del botón ureteral
Además, entre los túbulos urinarios vamos a encontrar un tejido intersticial
constituido por un conjuntivo laxo con vasos sanguíneos, vasos linfáticos y nervios.
Nefrona
Una nefrona es un tubo largo con
un extremo proximal ciego, dilatado e
invaginado denominado cápsula de
Bowman, en cuyo interior se sitúa un
ovillo capilar. Se continúa, de proximal
a distal, con:
x Túbulo proximal: tiene una
porción contorneada más
proximal y una recta más distal.
x Segmento delgado: más estrecho
que el túbulo proximal
x Túbulo distal: su primera porción
es recta y después se vuelve
contorneado. Posee una mácula
densa.
x Tubo colector.
Corpúsculos renales
La cápsula de Bowman junto con el ovillo capilar o
glomérulo renal forman el corpúsculo renal. Podemos dividir
el corpúsculo renal en dos polos:
x Polo vascular: constituye la zona de entrada de la
arteriola aferente y salida de a arteriola eferente para
constituir el ovillo capilar o glomérulo renal.
x Polo urinario: opuesto al arterial. Es el que se continúa
con la porción contorneada del túbulo proximal.
Tipos de corpúsculos renales:
x Corticales: están más próximos a la cápsula renal, y son los que se desarrollan
más tardíamente, pero los que permanecen después.
x Yuxtamedulares: están más próximos a la médula. Se desarrollan antes que los
corticales, pero van a sufrir un proceso de atresia fisiológica, con lo que al final
los que van a permanecer son los corticales.
190
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Además de en su localización, desarrollo y evolución, se diferencian en la
constitución de las nefronas, concretamente en el segmento conocido como asa de
Henle, que comprende el segmento delgado y los segmentos rectos de los túbulos
proximal y distal. Así:
x Nefronas corticales: su asa de Henle es más corta. Tienen un túbulo recto
proximal más largo y un segmento delgado corto que da paso al túbulo distal
justo en la zona de cambio de dirección. El túbulo recto distal también es largo.
x Nefronas yuxtamedulares: su asa de Henle es más larga. Tienen un túbulo
proximal corto, un segmento delgado largo que es el que forma el asa en sí, ya
que primero desciende y luego asciende, y un túbulo distal corto.
Al igual que veíamos que la secreción de las glándulas salivares se modifica en
el sistema ductal haciéndose hipoosmótica, en el riñón vamos a encontrar dos sistemas:
x Sistema filtrante: el glomérulo renal se encarga del filtrado, con lo que se forma
un ultrafiltrado de plasma
x Sistema tubular: constituido por el túbulo contorneado proximal, el asa de
Henle, el túbulo contorneado distal y el tubo colector. Se encarga de modificar el
ultrafiltrado glomerular.
Cápsula de Bowman
Presenta dos capas entre las cuales se sitúa el espacio urinario, encargado de
recoger el ultrafiltrado del sistema filtrante:
191
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Capa parietal: es la más externa, constituida por un epitelio plano simple
x Capa visceral: es la más interna, y por tanto recubre el glomérulo. Su epitelio es
muy característico y está constituido por podocitos. Los podocitos son un tipo
especial de células epiteliales estrelladas que se hallan adosadas a los capilares
del glomérulo renal, pero que presentan un rico citoesqueleto que les permite
emitir protrusiones hacia el espacio urinario. Estas protrusiones se conocen
como prolongaciones: las primeras se denominan primarias, su ramificación da
las secundarias y la ramificación de estas origina las terciarias. Las
ramificaciones terminales de los podocitos se denominan pedicelos
interdigitados.
Los podocitos sintetizan GEGF, que es el factor de crecimiento endotelial
glomerular, el cual estimula el crecimiento del endotelio glomerular y el
mantenimiento de las fenestraciones.
192
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Barrera de filtración glomerular
Denominamos así a la barrera que separa la sangre glomerular del espacio urinario
relleno de ultrafiltrado. Está constituida por:
x Endotelio capilar: al tratarse de capilares fenestrados, el endotelio capilar presenta
fenestraciones con diafragma (El cual puede perderse en las preparaciones
histológicas). Es muy plano y está revestido de glucoproteínas que le otorgan carga
negativa, siendo permeable al agua, urea, glucosa y pequeñas proteínas.
x Lámina basal glomerular: habitualmente fusionada con la lámina basal de los
podocitos, que también formaría parte de la barrera de filtración glomerular. Por
tanto, está formada por las propias células endoteliales y los podocitos, y
constituida por colágeno IV, fibronectina, laminina y heparán sulfato,
componentes que se encuentran en constante recambio. Es muy gruesa, de unos
300 nm, pero además en patologías como la diabetes se halla todavía más
engrosada. Podemos dividirla en tres capas:
1. Lámina rara interna
2. Lámina densa
3. Lámina rara externa
El colágeno IV interviene en tres enfermedades:
o Síndrome de Goodpasture: se trata de una enfermedad autoinmune
provocada por la formación de anticuerpos contra la cadena alfa 3 del
colágeno, que provoca una glomerulonefritis progresiva y hemorragia
pulmonar por destrucción de la lámina basal de la que el colágeno IV forma
parte.
o Síndrome de Alport: se trata de una enfermedad genética provocada por una
mutación del gen COL4A5 ligada al cromosoma X. En el glomérulo,
provoca el adelgazamiento, después engrosamiento y separación de la lámina
basal glomerular. Se da entonces una glomerulonefritis progresiva por salida
de proteínas y hematíes a través de la barrera de filtración glomerular no
funcional, lo que conduce a hematuria (Sangre en la orina) y finalmente lleva
a una insuficiencia renal. También se asocia con hipoacusia por defectos en
la función de la estría vascular y defectos oculares, especialmente en la
cápsula del cristalino.
o Hematuria benigna familiar: provocada por una mutación en el gen
COL4A4, no causa insuficiencia renal.
Existen otras patologías asociadas a la lámina basal, pero no específicamente al
colágeno IV, muchas de las cuales son glomerulopatías asociadas a mecanismos
inmunitarios. Las glomerulonefritis pueden deberse en estos casos a:
o Formación de Ac frente a componentes del glomérulo, como es el caso del
síndrome de Goodpasture
o Formación de complejos Ac-Ag circulantes, que quedan atrapados en la
barrera de filtración convirtiéndose en lugares de unión para proteínas del
complemento, tanto en la zona subepitelial como en la zona subendotelial.
Pueden inducir la proliferación de células epiteliales y mesangiales y atraer a
monocitos y neutrófilos.
193
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Pedicelos interdigitados de los podocitos y ranuras. Los pedicelos se anclan a la
lámina basal mediante integrinas de tipo α6B1, dejando entre ellos unos espacios
denominados ranuras de filtración, de unos 6 nm. Estas ranuras de filtración se
hallan cubiertas o atravesadas por un material membranoso, fundamentalmente
nefrina, que constituye el denominado diafragma de la ranura.
La nefrina es una molécula de adhesión capaz de unirse a otra nefrina de forma
homófila. El dímero se une a las moléculas transmembrana Neph1 y Neph2, así
como a la actina de los pedicelos mediante podocina, Z0-1 y CD2. Una
mutación en el gen para la nefrina causa el síndrome nefrótico congénito, mortal
al nacimiento, debido a una ausencia de diafragma de la ranura. Ello provoca
proteinuria, hipoalbuminemia, edema, hiperlipidemia y distensión abdominal.
En conjunto, la barrera de filtración glomerular retiene o deja pasar moléculas
según su tamaño, carga y forma. Quedan así retenidos los elementos formes de la sangre
y las grandes moléculas. Las moléculas sin carga inferiores a los 4 nm pueden pasar, las
superiores a los 8 quedan retenidas, y las que tienen un tamaño comprendido entre esos
dos pasarán o no en función de su carga.
194
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Orina primaria
Gracias a la barrera de filtración glomerular se permite la formación de un
ultrafiltrado de sangre capilar, que es el que constituye la orina primaria. La filtración se
produce gracias al gradiente de presión existente entre la sangre glomerular y el espacio
urinario, que es de unos 10 mm Hg. Se producen unos 125 mL/minuto de orina
primaria, pero de ellos se reabsorben 124 en el sistema tubular, con lo que se forma 1
mL/minuto (aproximadamente 150 mL diarios).
Túbulo proximal
Se halla en continuidad con la capa parietal de la cápsula de Bowman a través
del polo urinario, por lo tanto es un epitelio cúbico simple. Se trata de un tubo de unos
14 mm de longitud, en el que distinguimos como ya se ha dicho antes:
1. Parte contorneada o pars convoluta
2. Pars recta
Asa de Henle
El asa de Henle es la suma del segmento delgado y las dos porciones rectas (la
del túbulo proximal y la del distal). Hay nefronas que tienen un asa de Henle corta y
otras en las que es muy larga, lo que permite diferencias fisiológicas importantes. En las
cortas el segmento delgado descendente gira y da paso al túbulo recto distal. En las
largas, el segmento delgado es descendente y ascendente.
1. El túbulo proximal está en continuidad con la capa parietal de la cápsula de
Bowman por su polo urinario. Mide 14mm y está recubierto por epitelio cúbico
simple. Posee una pared contorneada/ convoluta y una pared recta. Podemos
observar túbulo contorneado proximal y túbulo recto proximal:
x El túbulo contorneado proximal está exclusivamente a nivel de la
corteza. El epitelio es cúbico simple con un núcleo redondeado con
tendencia a situarse en el tercio basal aunque generalmente está en el
centro y citoplasma eosinófilo. La membrana plasmática apical tiene un
número considerable de microvellosidades muy desarrolladas, lo que
permite visualizar al óptico el ribete en cepillo con glucocálix PAS+.
Aparecen gran número de vesículas de endocitosis, lisosomas, cuerpos
de Rhodin, RER, REL y Golgi, y abundantes zónula ocludens aunque no
son totalmente impermeables. Las células se interdigitan lateralmente. Al
óptico se ven estriaciones en la zona basal del citoplasma que son
invaginaciones de la membrana basal (bastoncitos Haiden-Hain). Son
paralelas entre sí, perpendiculares a la lámina basal y muy profundas.
Entre ellas hay mitocondrias con grandes crestas.
x El túbulo recto proximal se inicia en la corteza pero luego se mete en la
franja externa de la médula externa. El epitelio es cúbico simple con
tendencia a ser algo más bajo. Las células tienen microvellosidades
menos abundantes (ribete en cepillo menos desarrollado) y hay menor
195
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
desarrollo de los pliegues basales y de las interdigitaciones laterales, así
como menor número de mitocondrias.
2. El segmento delgado es la porción de la nefrona de menor diámetro. Está
tapizado por epitelio plano simple. Siempre está en médula (tanto su franja
externa como la interna es médula externa) y su longitud es muy variable,
dependiendo de la nefrona.
En las nefronas corticales es exclusivamente descendente y corta y después de
girar da paso al túbulo recto distal, mientras que en las yuxtamedulares
desciende, gira y asciende, alcanzando la médula interna y con ello siendo más
largo que en las nefronas corticales. Su diámetro es de 15μm y es importante no
confundirlo con capilares, aunque este error es fácil cometerlo. Al lado de estos
segmentos hay siempre capilares.
En este segmento delgado se definen histológicamente y de forma clásica cuatro
tipos celulares diferentes en función del grado de su grosor y del desarrollo de
los orgánulos:
Las células tipo IV son mucho más planas y finas mientras que las tipo II son
algo menos planas, más altas. Todas tienen desarrollo escaso de orgánulos, que
destacan algo más en las II y III. Las tipo I están sólo en el segmento delgado de
las nefronas corticales de asa corta (corticales) y poseen pocos orgánulos,
mientras que los otros tres tipos están en determinadas regiones del segmento
delgado de las nefronas yuxtamedulares (las II por todo el segmento delgado, las
196
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
de tipo III en la zona de médula interna y las IV en la curvatura y segmento
ascendente). Las de tipo II y III tienen muchos orgánulos.
La orina que penetra en este segmento delgado es isosmótica en comparación
con el plasma. Así es como penetra en la rama descendente, pero cuando sale
por la rama ascendete pasa a ser hiposmótica con respecto al plasma. Tras el
segmento delgado podemos observar el túbulo distal.
3. El túbulo distal es ascendente y se vuelve a meter en la corteza. El de cada
nefrona se aproxima a su corpúsculo de origen. La luz acaba siendo tan (o
incluso más) amplia como la del túbulo proximal y el epitelio no pasa
bruscamente de plano simple a cúbico simple, sino que hay un tramo de
transición donde hay células que van siendo progresivamente más altas y el tubo
va aumentando de diámetro, obteniendo al final el tubo distal. El túbulo distal
tiene:
x Parte recta que sería el túbulo recto distal
x Parte contorneada que sería el túbulo contorneado distal.
x Zona entre medias que es la pars maculata (mácula densa).
El túbulo recto distal nos lleva de nuevo a la corteza. Sus células sufren una
transición gradual a cúbicas y tienen escasas microvellosidades, por lo que no se
observa ribete en cepillo. Tampoco hay vesículas endocíticas. El núcleo redondo
tiende a estar en el tercio apical de la célula y el citoplasma sigue siendo
eosinófilo. Tiene desarrollo poco llamativo de orgánulos (escasos RER y Golgi)
pero tienen abundantes zónula occludens e interdigitaciones laterales. También
hay estriaciones basales y abundan los pliegues basales con mitocondrias
verticales dispuestas entre ellos.
A lo largo del asa de Henle se da una reabsorción de sales, Na+,K+, Cl- y en el
túbulo proximal se retira el 75% del agua. Cuando se retira toda la cantidad, hay una
disminución del volumen de la orina. Los elementos que se reabsorben vuelven a la
sangre.
Hay elementos que se reabsorben por transporte pasivo (acuaporinas), otros por
activo (bombas proteasas) y otros por endocitosis mediada por receptor, por eso en el
tubo proximal hay muchas vesículas endocíticas.
197
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Como se reabsorbe mucho Na+ se instala un gradiente osmótico que hace que el
agua entre directamente por ósmosis y por ello se retira mucho volumen de agua.
*En el síndrome de Fanconi el paciente no puede reabsorber determinados aminoácidos
o glucosa, de manera que los elementos se pierden por la orina. Luego hablamos de una
nefropatía de carácter hereditario o adquirida en la que hay una menor actividad de la
bomba ATPasa Na+/K+, con depósitos de Mg2+.
En el asa de Henle, el asa descendente
(túbulo recto proximal + segmento delgado), se
comporta de manera que es muy permeable al
agua y altamente impermeable al NaCl y a la
urea principalmente. El brazo ascendente (túbulo
distal + porción ascendente), es completamente
opuesto al descendente. Así tenemos un asa de
Henle con mitades fisiológicamente distintas.
El 10% de agua filtrada se reabsorbe en
este asa y el 20% de Na+ filtrado es reabsorbido.
Fármacos como los diuréticos inhiben la ATPasa
Na+/K+, de manera que no se reabsorbe el NaCl
y aumenta la excreción de NaCl y agua.
Como ya se ha mencionado, la orina entra isosmótica, sale hiposmótica y todas
las sales van al intersticio. Luego este intersticio es hiperosmótico. Este mecanismo,
este asa de Henle consigue una orina hipertónica (muy concentrada). Se concentra más
cuanto más largo sea el segmento delgado del asa de Henle.
Luego se establece un gradiente de hipertonicidad entre la unión cortico medular
y la papila. Así se observa que no hay el mismo grado de concentración a lo largo de
toda la médula renal.
Si hay distintas nefronas y estas se diferencien en la longitud, no todas las
nefronas contribuyen a concentrar la orina.
Mácula densa y túbulo contorneado distal
La mácula densa es una porción corta del túbulo distal. Está junto al corpúsculo
de origen, entre la arteriola aferente, a la que está pegada, que entra en el glomérulo del
corpúsculo y la eferente. Muestra un epitelio prismático con 20-30 células bastante
estrechas con un gran número de núcleos muy juntos que parecen superpuestos (están
apiñados), por lo que se tiñen más y dan sensación más basófila. Además, su membrana
plasmática presenta algunas microvellosidades y un cinocilio. No hay mitocondrias
verticales, luego la membrana es rectilínea.
La polaridad de estas células epiteliales está invertida (núcleo apical y Golgi
basal). Por otro lado, carecen de los pliegues basales típicos del túbulo recto pero sí
198
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
tienen prolongaciones basales que van a atravesar la lámina basal. Tampoco tienen
invaginaciones.
La mácula densa parece funcionar percibiendo cambios en la concentración de
sodio (NaCl) de la orina del túbulo distal. El cinocilio es un sensor que permite percibir
cambios en la concentración del Na+, cambios en la orina que llega a esta zona, entre
otros.
El túbulo contorneado distal está siempre en la corteza y es la parte final de la
nefrona. Es relativamente corto con una luz amplia y drena al conducto colector.
Muestra epitelio cúbico simple con células sin ribete en cepillo con alguna
microvellosidad corta. El núcleo es apical y el citoplasma menos acidófilo. Hay
abundantes lisosomas. Tienen pliegues basales e interdigitaciones laterales con
mitocondrias verticales. Esta porción es parecida al túbulo recto distal, pero las
invaginaciones de membrana son menos profundas.
Actúa como tubo impermeable al agua, excepto cuando tenemos niveles
elevados de ADH. Es impermeable a cloro y bicarbonato. Se produce secreción de iones
K+ y H+, y con ello aumenta la acidez, y se reabsorbe Na+. Es aquí donde la orina
comienza a ser claramente ácida. En estas células el amoniaco pasa a ser ion amonio.
La orina va al túbulo colector, pero antes está el túbulo reuniens entre el
conducto colector y el túbulo contorneado distal. Es corto y se encuentra tapizado por
epitelio plano simple. No forma parte de la nefrona, pero es una conexión entre el
túbulo distal y el colector. La orina llega al conducto colector y este recorre a manera de
eje un rayo medular.
Conducto colector
Inicialmente (en la zona cortical) muestra epitelio plano simple, para luego
hacerse cúbico. En la médula llega a ser de cúbico a prismático simple. Cada conducto
colector recoge la orina de unas 10 nefronas. Los conductos colectores convergen y se
forman otros de mayor calibre: Conductos Papilares de Bellini (con una luz mucho más
amplia). El conducto colector tiene dos tipos de células:
x Células Principales: Más pálidas. Presentan
microvellosidades gruesas y cortas y tienen un
cinocilio, sensible y que mide el flujo y
concentración de la orina. La membrana plasmática
de este cincocilio tiene policistina 1 y 2. Son
proteínas muy diferentes, la 1 tiene que ver con la
adhesión intercelular y celular con MEC, mientras
que la 2 funciona como canal para el calcio.
Estas dos proteínas son importantes y cuando sus
genes tienen mutaciones, se origina la poliquistosis
renal. Esta enfermedad es cromosómica dominante.
199
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
De manera que en el riñón se forman quistes renales debido a conductos
colectores dilatados que provocan que la orina no pueda atravesar, habiendo una
obstrucción. Se puede sobrevivir con diálisis o trasplante.
La membrana basal también forma pliegues y en ella hay distintas acuaporinas:
3 y 4. El resto de la membrana muestra otras proteínas muy importantes para su
funcionamiento, regulados por distintas hormonas:
o La apical tiene muchos canales, unos para el Na+ y otros para el K+.
Luego tenemos una bomba ATPasa Na+/K+ que está en la zona
basolateral. Esta célula controla la cantidad de Na que se va a eliminar
por la orina. Absorbe Na y segrega K.
o Aldosterona: Controla ambos sistemas. Así según la aldosterona la orina
segregada tendrá más o menos concentración de Na+.
o PNA: Este péptido inhibe los canales de Na y cuando se segrega al ano,
se absorbe menos Na y aumenta la cantidad de Na que se elimina por la
orina. Las células principales son las que controlan la cantidad de sodio
que se reabsorbe y la cantidad de potasio que se elimina.
o ADH: Actúa en las acuaporinas tipo II. LA ADH es una hormona
controlada, cuando la ADH esta elevada, las células se hacen permeables
al agua y se reabsorbe agua. Se elimina poco volumen de agua en la
orina, saliendo ésta muy concentrada.
Estas células también controlan el volumen de agua que se va a eliminar. Puede
ser de poco a mucho volumen (de sobrehidratación a deshidratación).
Este mecanismo por el cual las células principales reabsorben agua es
independiente de la reabsorción de Na+. El 20-30% del agua se reabsorbe en los
conductos colectores.
*Diabetes insípidas: puede ser la propiamente dicha en la que hay un déficit de
ADH (problema hipofisario) y los conductos colectores no reabsorberán agua y
se pierde mucha agua por la orina, llegando a estar en situación de peligro de
deshidratación. Otra posibilidad en esa situación se da no porque el túbulo
colector no responde a la ADH (diabetes insípida nefrógena).
200
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
La ingesta excesiva de agua es también perjudicial pues puede inhibir la
secreción de ADH lo que conduciría a la formación de grandes volúmenes de
orina hipoosmótica.
x Células Intercalares: Son más basófilas, oscuras. No se distribuyen por igual,
sino que están principalmente en la porción cortical y el número va a ir
disminuyendo conforme avanzamos hacia la médula.
Estas células presentan algunas microvellosidades, no presentan pliegues en las
membranas basales. Tienen muchas mitocondrias, micropliegues e
interdigitaciones. Estas células juegan un papel fundamental en la reabsorción
del K, en la secreción de más bicarbonato e hidrogeniones y regulan el equilibrio
ácido-base, importante para determinar el pH de la orina.
Los conductos colectores de Bellini se abren en la punta de la papila. Esa zona
donde hay muchos puntos que se abren es el área cribosa. Estos conductos tienen
epitelio prismático simple y abundantes zonulae occludens.
Tejido Intersticial
Se trata de un tejido conjuntivo laxo entre túbulos y corpúsculos. Abundantes
fibras colágenas y reticulares, abundancia de proteoglicanos formando el estroma a
modo de red de sostén. La cantidad de tejido intersticial no es uniforme, es menor en
corteza y va a aumentar según avanzamos a la médula; siendo 20% en la médula interna
(con células intersticiales cargadas de lípidos) y 7% en corteza (fibroblastos y APC).
El tipo de células que hay en el intersticio a nivel cortical son los fibroblastos,
células presentadoras de antígeno que sintetizan y renuevan intersticio además de
producir EPO. En la zona medular predominan las células intersticiales cargadas de
lípidos. Las células intersticiales van paralelas a los túbulos
Estas células contribuirían a compactar este tejido aunque no se sabe con
certeza. Lo que sí sabemos que estas células emiten prolongaciones que contactan con
las células de los túbulos, yendo también hacia la lámina basal.
Los núcleos de estas células aparecen en posición transversal, como si fueran los
peldaños de una escalera.
Tienen un buen desarrollo de todos sus orgánulos (RER y AG sobre todo),
tienen capacidad contráctil y debido a eso se piensa que tienen capacidad de contraerse
y regular el avance sanguíneo, pues están al lado de túbulos y capilares (bajo control
hormonal).
Tienen inclusiones lipídicas y estas inclusiones tienen Prostaglandina tipo E2.
Los lípidos están implicados en la síntesis de medulipina I. La medulipina I es un
precursor de la medulipina II. La II es un vasodilatador, contribuyendo a bajar la presión
arterial. Se ha demostrado que los lípidos de estas células contribuyen a la eliminación
de Na+.
201
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Clínica
Una insuficiencia renal crónica causa anemia secundaria. Estos fibroblastos
actúan como células que sintetizan eritropoyetina. Luego si una persona padece
insuficiencia renal es normal que padezca a su vez una anemia de tipo secundario.
La nefritis intersticial es causada por fibroblastos activados, macrófagos y
linfocitos. Su origen puede ser inmunológico o deberse a fármacos nefrotóxicos
(metales pesados). La nefritis de origen intersticial se debe a que los fibroblastos del
intersticio se activan y aparecen en gran cantidad macrófagos y linfocitos, acabando en
una inflamación del riñón con origen intersticial.
Mesangio
Es una estructura intraglomerular entre capilares, formado por células
mesangiales y MEC mesangiales. Es laxa, PAS+, con colágeno tipo IV, V y VI,
fibronectina, laminina y perlecán. Es más fibrilar que la lámina basal.
Además, las células mesangiales se agregan fuera del glomérulo (células
mesangiales extraglomerulares), en un espacio limitado por la mácula densa y las
arteriolas aferentes y eferentes glomerulares. Las células mesangiales intraglomerulares
pueden ser continuas con las células mesangiales extraglomerulares.
Las células mesangiales son estrelladas, con núcleos grandes y oscuros,
lisosomas y lipofucsina. Tienen filamentos de actina y miosina. Sus prolongaciones se
introducen entre los capilares, atravesando la lámina basal y las fenestraciones. Tienen
movilidad y son pericitos modificados. Son contráctiles, fagocíticas y secretoras.
Sus funciones son:
x Sostén: Actúa de esqueleto intraglomerular
x Síntesis de ME mesangial
x Fagocíticas: renovación de la lámina basal. Control de la barrera de filtración
glomerular.
x Síntesis de PG, IL1, PDGF
x Síntesis de endotelinas: constricción de vasos
x Contracción
x Rc para angiotensina II
x Regulación del flujo sanguíneo glomerular
202
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
La glomerulonefritis: membranoproliferativa, membranosa y nefropatía por IgA
o enfermedad de Berger. Se produce por acumulación de Igs contra componentes de
mesangio o complejos Ac-complemento en la ME mesangioal. Proporcionan puntos de
unión para el complemento, afectando a los capilares, lo que ocasiona una reacción
inflamatoria que lleva a la oclusión de los vasos.
Aparato yuxtamedular
Es un órgano endocrino formado por la mácula densa, células mesangiales
extraglomerulares, del lacis o de Goormaghtigh y células yuxtaglomerulares o
mioepiteliales de Ritter.
Está limitado por la mácula densa y las arteriolas aferentes y eferentes. Es un
mecanismo de retroalimentación túbuloglomerular que permite la autorregulación del
flujo sanguíneo y de la filtración glomerular.
Mácula densa
Posee lámina basal discontinua con prolongaciones que alcanzan células
yuxtaglomerulares. Sus células son más estrechas y más altas. Su función es percibir
cambios en NaCl en la orina del túbulo distal.
203
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Células mesangiales extraglomerulares
Están en continuidad con las células mesangiales, con nexos entre estas y ellas
mismas. Su función es percibir cambios en NaCl en la orina del túbulo distal.
Células yuxtaglomerulares
Pertenecen a la pared de la arteriola aferente o eferente y son miocitos
modificados de la túnica media. Son células epitelioides, redondeadas y claras, ricas en
RER, aparato de Golgi y gránulos (I y II).
Se encargan de la síntesis, almacenamiento y secreción de renina, encargada de
formar angiotensina I. Escinde angiotensinógeno en angiotensina I, la cual es convertida
en angiotensina II por la ECA en el pulmón. Este sistema se va a encargar de controlar
la homeostasis de Na+ y la hemodinámica renal.
La angiotensia II es un potente vasoconstrictor, que aumenta el flujo renal y con
ello la filtración y las presiones sistólica y diastólica. Además, aumenta la síntesis de
aldosterona en el túbulo colector y la rama ascendente del asa de Henle, lo que lleva a
un aumento de la reabsorción de Na+ y agua por túbulo contorneado proximal.
La reabsorción de NaCl producida en el túbulo contorneado distal se ve
estimulada, como la secreción de ADH (aumenta la reabsorción de NaCl y agua).
En definitiva, son barorreceptores que reaccionan ante cambios en la presión
arterial. Si la presión aumenta se inhibe la liberación de renina. La mácula densa va a
percibir cambios en la concentración de NaCl. Si hay una disminución de Na+, o por
efecto de SNS, dopamina y noreprinefrina, se estimula la secreción de renina.
204
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Funciones renales de equilibrio hídrico
Se puede perder agua por orina, por evaporación (piel, espiración) y por heces,
aunque la excreción renal es independiente de otras sustancias. La orina puede ser
hiperosmótica o hiposmótica.
El túbulo contorneado proximal tiene orina isoosmótica. La rama descendente
del asa de Henle es muy permeable al agua y poco al NaCl y el intersticio es
hiperosmótico, lo que hace que la orina sea hiperosmótica en el giro del asa. La rama
ascendente va a ser impermeable al agua y permeable al NaCl, que es reabsorbido.
Finalmente, la orina es hipoosmótica.
En el túbulo contorneado distal y en el conducto colector cortical se reabsorbe
NaCl. Si ADH aumenta se estimula la reabsorción de agua, lo que continúa el proceso
de concentración de orina y la convierte en hiperosmótica. Sin ADH estas zonas van a
ser impermeables al agua.
205
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
El proceso es multiplicador e intercambiador de corriente, pues hay dos tubos
paralelos con dirección de flujo opuesta (los túbulos rectos del asa de Henle), estando
los capilares (asas capilares de vasos rectos) próximos a ellos y con un flujo también
opuesto a su correspondiente tubo, lo que permite el intercambio de calor o sustancias.
Se produce transporte activo de Na+, K+ y Cl-.
Todo este proceso genera un intersticio hiperosmótico, aumentando el NaCl y el
gradiente con la profundidad de la médula. Los vasos rectos se van a encargar de
eliminar agua y solutos.
Este proceso permite la formación de un orina hipertónica, con pH 5,5 y que
permite la expulsión de urea, ácido úrico, amonio y creatinina.
206
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Regulación de la absorción de agua y NaCl
x La angiotensina II estimula la reabsorción de NaCl y agua. En situación de
hipovolemia se activa la secreción de renina.
x La aldosterona provoca la reabsorción de NaCl en la rama ascendente del asa de
Henle, en el túbulo contorneado distal y en el tubo colector. El aumento de los
niveles de angiotensina II/K+ estimula la secreción de aldosterona.
x El factor natriurético auricular estimula la excreción de agua y NaCl e inhibe la
liberación de ADH.
x La urodilatina, secretada por el epitelio del túbulo contorneado distal y el tubo
colector, va a estimular la reabsorción de agua y NaCL en el tubo colector
medular.
x La ADH (núcleo supraóptico y paraventricular del hipotálamo) va a tener
distintos efectos:
o Hipovolemia: aumenta la permeabilidad al agua en el tubo colector,
estimulando la reabsorción.
o Sin efectos sobre la secreción de urinaria de NaCl.
VÍAS URINARIAS
x Cálices menores
x Cálices mayores Seno renal
x Pelvis renal
x Uréter
x Vejiga
x Uretra
Cálices
4 cálices menores desembocan en uno mayor. Posee urotelio,
lámina propia (densa en la superficie y laxa en profundidad, con fibras
elásticas y nódulos linfoides), capa muscular (longitudinal interna y
circular externa) y adventicia.
Uréter
Mide 24-34 cm. Está incluido en el tejido adiposo
retroperitoneal, con la porción proximal dilatada (pelvis renal). La
porción distal se introduce en la pared vesical.
Se puede producir nefrolitiasis, cálculos renales por sales
cálcicas, ácido úrico o acetato de Mg-amonio por cristalización. Si
obstruye el uréter la contracción de la muscular provoca dolor
207
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Vejiga
Receptáculo distensible situado en la pelvis, tras la sínfisis pubiana. Se distingue
un trígono formado por los orificios ureterales y por el orificio uretral interno. Su origen
embrionario distinto, deriva de los conductos mesonéfricos, y el resto de la cloaca. Es
liso y de grosor constante, no se puede distender a diferencia del resto de la vejiga.
Mucosa
Muy plegada en estado contraído, y lisa en estado dilatado. Posee urotelio y
lámina propia (igual que anterior).
El urotelio es estratificado con un número de
capas diferente según la zona, desde 2-3 hasta 7-8. Es
la distensión se aplana y es muy poco permeable
(zónula occludens). Las células basales son cúbicas-
prismáticas, aplanándose en zonas superiores, siendo
más redondeadas, claras y binucleadas. El citoesqueleto
es rico y tira de la membrana cuando no es necesaria
tanta superficie, internalizándose en forma de vesícula,
y formando una superficie en cúpula en la distensión.
La membrana plasmática muestra placas rígidas y gruesas, que se unen por
filamentos de actina.
Muscular
Hay una capa interna longitudinal y otra externa circular. La parte distal de los
uréteres y la vejiga tienen un tercer estrato longitudinal. Es un esfínter funcional
formadao por 3 capas: las capas no son independientes, sino que hay una estructura
continua espiralada (músculo detrusor). Las ondas peristálticas que genera va a inducir
la expulsión de la orina.
208
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Al distenderse la vejiga se comprimen los orificios ureterales, impidiendo el
reflujo. Cuando la vejiga se contrae los orificios siguen cerrados por la contracción del
músculo vesical. Si la orina pudiera volver a ascender se producirían infecciones
superiores.
La inervación es simpática y parasimpática en uréteres y vejiga, formando el
plexo vesical. La simpática inerva los vasos, y la parasimpática la musculatura, y dan
lugar al reflejo de micción, estimulado por fibras sensitivas aferentes. Si se extirpa no se
modifica el peristaltismo.
Adventicia
Parte de los uréteres y la vejiga están revestidos de peritoneo.
Uretra
Es un tubo fibromuscular desde la vejiga al exterior. En el varón va a ser larga,
formando también parte de las vías espermáticas. En la mujer es más corta.
Varón
Su longitud alcanzo los 20 cm y tiene tres segmentos:
x Uretra prostática: 3-4cm. Recibe los conductos eyaculadores y de glándulas
prostáticas. Tiene urotelio y lámina propia laxa y muy vascularizada.
x Uretra membranosa: 1cm. Atraviesa el diafragma urogenital de la pelvis y entra
en el periné. Es un músculo esquelético que actúa de esfínter. Su epitelio es
prismático estratificado.
x Uretra peneana: 15 cm. Se abre en el glande, en la fosita navicular. Recibe las
glándulas de Cowper y de Littré. Su epitelio es prismático estratificado o
pseudoestratificado, convirtiéndose en epitelio plano estratificado en el glande.
La mucosa está rodeada por el cuerpo esponjoso del pene y carece de capa
muscular.
Mujer
La uretra es corta, de unos 3-4cm, terminando en el vestíbulo vaginal, justo
detrás del clítoris. Su mucosa forma pliegues L y es rodea por el esfínter ureteral
externo al perforar el diafragma urogenital.
Inicialmente posee urotelio, pero finalmente se transforma en epitelio plano
estratificado. Bajo el epitelio hay una lámina propia de fibras elásticas. Hay un plexo
venoso similar al cuerpo esponjoso penenano. La musculatura interna es lisa, mientras
que la externa es esquelética. Consta de distintas glándulas:
x Glándulas uretrales: Son pequeñas y se encuentran en la pared proximal.
x Glándulas parauretrales: Homólogas a la glándula prostática de los hombres.
Vierten su secreción, alcalina, a ambos lados del orificio uretral externo.
209
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
APARATO GENITAL MASCULINO
CÉLULAS GERMINALES PRIMORDIALES
Las células germinales primordiales extraembionarias (espermatogonias) se detectan en
la cuarta semana en la pared del saco vitelino. Son células grandes, con prolongaciones,
núcleo redondeado en que destacan uno o dos nucléolos, citoplasma con glucógeno,
mitocondrias con crestas tubulares, polisomas y RER. Presentan fosfatasa alcalina.
A partir de la cuarta semana hasta la sexta migran, a la vez que proliferan, a lo largo del
mesenterio dorsal del intestino posterior mediante movimientos ameboides.
Encontramos entre 2500 y 5000 células que pueblan el blastema gonadal (el
mesénquima) y que inducen la proliferación del epitelio celómico.
Este proceso migratorio es posible gracias a la interacción del factor de célula madre
con el receptor c-Kit. Si el ligando o el receptor faltan, tenemos un déficit de células
germinales primordiales, lo cual afecta a la hematopoyesis, desarrollo de melanocitos y
mastocitos.
Es decir, que a partir del 2º mes, las células germinales se encuentran a la altura de la
décima vértebra dorsal, donde inducen la proliferación de células del mesonefros y el
epitelio celómico (segregando factores que inducen a ello). Las células del mesonefros
revestidas por celoma formarán el blastema gonadal, que es una estructura
indiferenciada (cordones que vienen del mesonefros revestidos de epitelio). Estas
células son resistentes a la radioterapia y quimioteramia (así, el tratamiento antitumoral
va a afectar a las espermatogonias que están experimentando mitosis y divisiones, pero
las células madre repueblan el epitelio cuando se interrumpe el tratamiento).
Finalmente, las células primordiales o madre germinales invaden el blastema en la sexta
semana. Los cordones ahora serán cordones sexuales primordiales o
primitivos. También se forman crestas gonadales.
Bajo ellas, encontramos una lámina basal y por fuera un estroma gonadal
indiferenciado. Así, las células que habían invadido la gónada permiten la formación de
los cordones testiculares y la gónada masculina se formará a partir de la gónada central
indiferenciada. Si el individuo es XY el gen SRY del cromosoma Y codificará para
TDF (factor determinante del testículo) y MIS (inhibidora mülleriana). Entonces los
cordones testiculares medulares que terminan de madurar y forman los túbulos
seminíferos. A partir de la séptima semana de vida, esta gónada se desarrolla hacia la
vía masculina (testículos).
Después, en zona del hilio de la gónada en diferenciación, los cordones se disgregan y
forman zona laberíntica que será la red de Haller o Rete Testis.
210
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Hablamos entonces de gonocitos, no de células germinales, y los cordones dejan de ser
sexuales para pasar a ser cordones testiculares o medulares que estarían formados por
células de Sertoli/de sostén y gonocitos (células dentro del cordón). Entre los cordones
que hay en la gónada hay un mesénquima que formará tejido intersticial en el cual
existen células de Leydig. Los cordones testiculares constituirán los túbulos
seminíferos.
Inicialmente, estas células germinales y gonocitos, se sitúan en zona central de cordón.
Son células grandes, con un solo núcleo con nucléolo marcado, mitocondrias con
crestas tubulares, elevados niveles de fosfatasa alcalina y el protooncogen c-kit. Estas
células se siguen dividiendo dentro de los cordones (sexuales primitivos), es decir, que
la mitosis no es exclusiva del periodo migratorio y a partir de semana 12 se desplazan
hacia la base del cordón (apoyándose en la lámina basal, algo que no ocurría cuando
estaban en el centro del cordón). Así, aquí finalmente tendremos cordones formados por
células de Sertoli, gonocitos y espermatogonias fetales (células hijas de las anteriores)
Los gonocitos persisten en todo el periodo incluso hasta 6 meses después del
nacimiento. Sin embargo, vamos a tener una bajada muy marcada entre la semana 16 y
la 22.
Parte de las células que originan los gonocitos (sus células hijas) son espermatogonias
fetales. Son células más pequeñas que los gonocitos, su cromatina está mucho más
condensada y tienen una gran reserva de glucógeno (son PAS+). Las espermatogonias
tienen mitocondrias con crestas no tubulares, unidas entre sí mediante un elemento
electrodenso, RNA+.
Además, hay que destacar que estas células también se dividen, persistiendo en todo el
periodo embrionario y más allá del nacimiento (apareciendo hasta los 2 y 3 años de vida
del niño)
Todas las células que se forman por división a partir de un gonocito, se mantienen
unidas por puentes intercelulares, lo que es muy llamativo porque son el primer tipo
celular que los muestran (la citocinesis no es completa). Por tanto estas células no son
independientes sino que están dentro de un cordón unidas.
Hacia la semana 16, 17, 20 y 21, comienza la degeneración celular en las gónadas. Dura
estas 4 semanas y afecta a cualquier tipo celular (especialmente a los gonocitos). De
esta manera, tras la semana 22 los cordones celulares tienen como célula principal la
espermatogonia fetal.
La mitosis de estas espermatogonias fetales son reguladas por un factor promotor de la
mitosis MAS y otro inhibidor MIS. Predomina el inhibidor.
Durante el resto de la gestación no hay cambios en los cordones testiculares. Al nacer,
los túbulos seminíferos son cordones macizos, que tienen sobre todo células de Sertoli.
El epitelio seminífero está rodeado de una lámina basal de células peritubulares
(aplanadas) con células de Sertoli, gonocitos, espermatogonias fetales y un nuevo tipo
211
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
celular, las espermatogonias A (aparecen muy al final de la gestación). Todas estas
células se asientan sobre la lámina basal, bajo la cual hay miofibroblastos.
Existen ciertos autores que describen otros tipos celulares que son transición entre la
espermatogonia fetal y el tipo A.
Los gonocitos se siguen detectando hasta los 6 meses. Las espermatogonias fetales
duran hasta 4 años, de modo que encontramos túbulos seminíferos que contienen sobre
todo espermatogonias A. Las mitosis que experimentan estas células duran unos pocos
meses y son dependientes de gonadotropinas (FSH y LH). Estas gonadotropinas
disminuyen su nivel hasta que cesan sus mitosis en el sexto mes. Después se produce un
reposo de 2 o 3 años, apareciendo una oleada de mitosis de origen desconocido (no
dependen de gonadotropinas) que da lugar a las espermatogonias B.
Antes de la pubertad, el testículo crece y madura. Se alargan y ensanchan los túbulos
seminíferos y se ahuecan hacia los 9 años. Al final de esta etapa, hay un gran
crecimiento de células germinales regulado por gonadotropinas, cuyos niveles aumentan
irregularmente. Hay una tercera oleada proliferativa, cuando se estabilizan los niveles
de gonadotropinas y se consigue ya espermatogénesis completa (13-14 años).
Espermatogénesis: tipos celulares
Es el proceso por el cual las espermatogonias se van a dividir y llevar a cabo una serie
de procesos de diferenciación para formar los espermatozoides. Esto en el caso del
varón, ocurre de forma constante y continua, por lo que necesitamos que se generen las
espermatogonias B a partir de las A de forma constante (renovación constante). Por otro
lado necesitamos una temperatura 2 o 3 grados menor que la corporal para poder
llevarlo a cabo.
Espermatogonias
Son células diploides. Cuando comienzan a dividirse, las células que resultan están
unidas por puentes. Tanto las unas como las otras se consideran células madre. Las A se
dividen activamente y forman las de tipo B que aseguran la renovación de las células
del tubo seminífero.
x Espermatogonia A: Las encontramos en la base del cordón, sobre la lámina
basal. Con núcleos más ovalados y claros pegado a la envuelta nuclear,
nucléolos periféricos, se dividen en densas y claras. Las densas tienen núcleo
heterocromático, y las claras, eucromático. Además tienen el nucléolo adosado a
la envuelta y, al igual que las fetales están unidas por puentes citoplasmáticos.
Encontramos 4 tipos según el núcleo que tengan:
o Ad: núcleo esférico y de contorno más irregular. Vacuola nuclear sin
membrana. Mayoritarias al principio.
o Ap: núcleo esférico pálido y regularo Al: citoplasma y núcleo alargado
cromatina más oscura, densa, con más heterocromatina y vacuolas
nucleares.
212
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
o Ac: núcleo esférico con zonas claras y oscuras intermedio entre ap y ad.
Tienen muchos polisomas, algo de RER y REL y AG. Las mitocondrias
siguen unidas por RNA+, tienen crestas no tubulares y se agrupan
alrededor del núcleo. Aparecen en el citoplasma dos estructuras:
Nuage: una nube de material denso fibrilar que esta libre en el
citoplasma.
Cristal de Lubarsch: filamentos paralelos de unos 15 nm de
diámetro entre los que hayespacios de unos 20 nm ocupados por
gránulos que forman hileras.
Estas espermatogonias aparecen en el nacimiento y se dividen durante los primeros
meses de vida respondiendo a niveles elevados de FSH y LH (que no se sabe por qué
aumentan en esta etapa) pero de repente entran en una fase de reposo o quiescencia que
de entre los 6 meses hasta dos años y medio/ tres. Pasado este tiempo aparece una nueva
ola de divisiones que afecta a todas las espermatogonias. A partir de estos 2 años y
medio aparecen espermatogonias A y también un nuevo tipo, las espermatogonias
B (casino hay gonocitos y están desapareciendo por completo las espermatogonias
fetales). Excepcionalmente podemos encontrar algunos testículos la proliferación de
espermatocitos y espermátidas.
x Espermatogonia B: Núcleo redondo, con cromatina densa y aspecto granular con
uno o dos nucléolos no asociados a la envuelta que no se encuentran en zonas
tan basales (los núcleos) pero siguen teniendo contacto con la lámina basal. El
citoplasma se parece al de las A pero con más polisomas y sin cristales de
Lubarsch, pero las mitocondrias sí se unen por el material RNA+ y se suelen
agrupar cerca del núcleo.
DESARROLLO SEXUAL MASCULINO
Sexo
Tenemos que distinguir varios conceptos antes de empezar:
x Sexo genético: determinado en la fecundación por la presencia o ausencia del
cromosoma Y (sexo masculino y femenino respectivamente).
x Sexo gonadal: determinado por el segmento SRY, situado en el brazo largo del
cromosoma Y, y que codifica para el TRF o factor determinante testicular.
x Sexo hormonal: determinado por la aparición de testosterona,
dihidrotestosterona y sustancia antimülleriana (MIS) en el embrión masculino.
La gónada está indiferenciada hasta la 7ª semana del desarrollo. En ese momento, el gen
SRY controla la diferenciación de la gónada hacia testículo. La parte conocida como
corteza gonadal va a involucionar, mientras que la médula va a constituir el testículo en
sí.
213
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Las células de Sertolli se encargan de la secrción
de la sustancia antimülleriana, a partir de ahora
MIS, que conduce a la regresión del conducto de
Müller y con ello impide el desarrollo de las
estructuras derivadas de él. Si las células de
Sertolli no secretan MIS, lo que va a suceder es
que el conducto de Müller se va a desarrollar y va
a originar oviductos, útero, cérvix uterino y el
tercio superior de la vagina, que es lo que sucede
por ejemplo en pacientes con disgenesia gonadal
de tipo XY.
Clínica: disgenesia gonadal XY
Se trata de pacientes con estructuras de origen
mulleriano (oviductos, útero, cérvix uterino y
tercio superior de la vagina) por ausencia de MIS,
que presentan poco desarrollo gonadal.
Las células de Leydig se encargan desde la 8ª
semana del desarrollo de la producción de
testosterona, por estimulación de Hcg. La
testosterona induce la diferenciación del conducto
de Wolff, que dará lugar al epidídimo, conducto
deferente, vesículas seminales y conductos
eyaculadores.
En la mujer, debido a la ausencia de andrógenos, el conducto de Wolff sufre regresión.
En mujeres con elevados niveles de testosterona y andrógenos por tanto va a persistir el
conducto de Wolff, pero también el de Müller.
Clínica: síndrome de insensibilidad androgénica
Este síndrome está provocado por un defecto genético que determina la infraexpresión o
ausencia de expresión del receptor de andrógenos. Con ello, aunque el individuo
presente unas células de Leydig productoras de testosterona que le permitan tener
niveles normales e incluso elevados de esta hormona, no va a tener ninguna de las
estructuras derivadas del conducto de Wolff (epidídimo, conductos deferentes,
conductos eyaculadores y vesículas seminales). Sin embargo, dado que la funcionalidad
de las células de Sertolli es perfecta, sí que va a producirse la regresión del conducto de
Müller. Como resultado, vamos a tener un individuo que no tiene genitales internos
funcionales, y cuyos testículos no han descendido, presentando genitales externos
femeninos.
Por acción de la 5alfa-reductasa, la testosterona se transforma en dihidrotestosterona
(recordemos que es la única sustancia que interviene en el sexo hormonal que nos falta
214
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
por comentar). La presencia de dihidrotestosterona o DHT determina la formación del
tubérculo, el pliegue, la tumefacción y el seno genitales, que darán lugar al pene, el
escroto y la próstata.
Los individuos que no tienen 5alfa-reductasa o presentan un déficit en ella no van a
desarrollar genitales externos masculinos, puesto que ello está mediado por la DHT, con
lo que son frecuentemente confundidos con mujeres en el momento del nacimiento,
puesto que además si no existe DHT se van a desarrollar los labios mayores y menores,
el clítoris y los dos tercios inferiores de la vagina, esto es, genitales externos femeninos.
215
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
CÉLULAS DE SERTOLI
Son las más abundantes en la infancia y en la vejez. En la etapa de adulto son
minoritarias y proceden de divisiones. Son muy altas, estrelladas, con un núcleo claro y
se encuentran en el centro del epitelio con un nucléolo grande. Presentan todos los
orgánulos (muy desarrollados). También tienen un buen citoesqueleto como buena
célula epitelial.
Hay Cristales de Charcot que son como los de Luschard. Las células de Sertoli crean
avientes, espacios, donde abrazan y entran en contacto con las otras células: nichos y
criptas. Establecen contactos directos con otras células. Son fundamentales estos
contactos para que las células puedan dividirse y diferenciarse. Establecen uniones
estrechas, uniones de comunicación y de adhesión. Se originan 2 compartimentos
diferentes:
x En la zona basal: el que queda entre la célula de Sertoli y/o sus prolongaciones y
la lámina basal. De manera que en ese espacio lo que encontramos son
espermatogonias. También los espermatocitos de primer orden en sus etapas
iniciales. Los espermatocitos primarios van a sufrir meiois y antes del estado de
leptotene están en el compartimento basal.
x Espacio adluminal: todo lo que queda por encima de la basal.
Son espacios que tienen microambientes completamente distintos y están
herméticamente separados. Estos dos espacios se separan por los complejos de unión los
cuales sellan los dos espacios, principalmente el espacio basal.
Los espermatocitos primarios tienen que subir, para ello rompen las uniones, suben y
rápidamente se vuelven a formar las uniones en el mismo sitio. La regulación consiste
en que las uniones se formen y vuelvan a ser completamente herméticas una vez que los
espermatocitos han pasado.
Necesitamos dos compartimentos distintos para formar la barrera hematotesticular. Esto
sirve para proteger el compartimento adluminal. Este compartimento debe estar
separado para proteger frente a todas las demás células que no son de la estirpe
germinal. Ya que las de germinal son genéticamente diferentes. Además, se podrían
producir reacciones contra estas células, tienen que evitar el contacto con la sangre
porque por la sangre pueden llegar factores patógenos. Si la barrera no está bien aislada,
se produce autoinmunidad.
Responden a dos controles: FSH y testosterona. Son necesarios para que se dé la
espermatogénesis.
Son células de sostén que forman la barrera hematotesticular. Progtegen y nutren a las
células espermatogénicas. Además, inducen la espermatogénesis y facilitan la migración
de la zona basal a la apical de las células espermatogénicas.
Van a fagocitar cuerpos residuales y facilitan la espermiación (liberación de
espermátidas maduras por contracción).
216
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Además, tienen funciones endocrinas: producen estrógenos, excretan líquido testicular
(iones y proteínas) y sintetizan ABP (proteína transportadora de andrógenos), encargada
de fijar la testosterona. Van a influir en la FSH mediante:
x Inhibina: inhibe la síntesis de FSH
x Subunidad activina: estimula la síntesis de FSH.
Se encarga de la síntesis de MIS, que va a degenerar el conducto de Müller.
ESPERMATOGÉNESIS
La espermatogénesis es un conjunto de divisiones y cambios por el que las
espermatogonias originan espermátidas. Es una producción continua que requiere una
Tª de 2-3ºC menor a la corporal. Son diploides y proceden de células unidas por puentes
de unión. La espermiogénesis, por su parte, produce la formación de espermatozoides.
Cada gonia de lugar a cuatro espermátidas, y la duración media es de 64 días.
Tiene lugar en el epitelio germinativo, de modo que la maduración de este permite la
formación de gametos masculinos. Este epitelio está compuesto por células
espermatogénicas. Las más pegadas a la membrana basal son las células madre
espermatogénicas y las espermatogonias Ay B, que se dividen por mitosis hasta que
reciben señales hormonales. Muchas de ellas están conectadas por medio de puentes
citoplasmáticos.
Cuando las espermatogonias comienzan la primera división meiótica, pasan a llamarse
espermatocitos primarios, que aún no han perdido los puentes citoplásmicos, como
tampoco lo hacen los secundarios, obtenidos cuando los primarios finalizan la primera
división meiótica. Cuando los espermatocitos secundarios atraviesan la segunda
división meiótica se convierten en espermátidas inmaduras, que por un proceso de
maduración conocido como espermiogénesis se convierten en espermatozoides.
La función de las células de Sertoli es separar el compartimento adluminal del basal,
puesto que en la superficie de los espermatozoides hay una serie de proteínas que
podrían ser reconocidas como extrañas si pasasen al torrente sanguíneo, de manera que
se producirían problemas de
infertilidad.
Espermatogonias
Las espermatogonias son células
espermatogénicas diploides que se
encuentran en contacto directo con
la lámina basal en el compartimento
basal. Se localizan por debajo de las
uniones oclusivas entre las células
de Sertoli y están, por tanto, fuera de
217
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
la barrera hematotesticular. Las espermatogonias proceden de células madre
espermatogónicas y sufren sucesivas divisiones mitóticas, que empiezan en la pubertad.
Se pueden observar dos tipos de espermatogonias a nivel morfológico:
x Las espermatogonias de tipo A, que muestran un núcleo oval eucromático
paralelo al eje mayor y un nucléolo unido a la envoltura nuclear. Representan la
población de células madre que mantiene mediante mitosis un número adecuado
de espermatogonias a lo largo de toda la vida.
x Las espermatogonias de tipo B, que tienen un núcleo redondo, masas de
heterocromatina unidas a la envoltura nuclear y un nucléolo central. Destinadas
a entrar en meiosis, se originan a partir de las de tipo A.
Tipo A
Tipo B
Con la pubertad adquieren movilidad. Sus descendientes van a quedar unidos mediante
puentes citoplamáticos.
Espermatocitos I
Los descendientes de las espermatogonias de tipo B, que han entrado en la primera
división meiótica, son los espermatocitos primarios. Estos se localizan justo por debajo
de la capa de espermatogonias pero aún inmersos en el citoplasma de las células de
Sertoli.
Están separados de las espermatogonias por prolongaciones laterales de células de
Sertoli. Son descendientes conectados por puentes citoplásmios y tienen forma
redondeada.
Los espermatocitos I son diploides, 4C, y mayores. Van a atravesar la barrera
hematotesticular para alcanzar el compartimento adluminal. Bajo ellos se establecen
nuevas uniones, siendo las previas degradadas.
218
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Van a entrar en la primera división meiótica, donde hay selección aleatoria de
cromosomas paternos y maternos, con una profase muy lenta. La meiosis I tiene lugar
en 22 días:
x Profase
o Leptotene: translocación a compartimento adluminal
o Zigotene: el mRNA no traducido da lugar a la síntesis de protaminas, que
sustituyen a las histonas nucleares y permiten el grado de compactación
de la cromatina nuclear necesario durante las fases finales de la
formación de los espermatozoides.
o Paquitene
o Diplotene
x Metafase
x Anafase
x Telofase: 2 espermaticitos II o secundarios.
Espermatocitos II
Los espermatocitos I originan espermatocitos II que mantienen los puentes
citoplasmáticos. Son células haploides, 2C, que no sintetizan ADN. Poseen un núcleo
eucromático sin nucléolo, característica importante para la diferenciación, por ejemplo
respecto a las espermátidas.
Su presencia es muy breve porque
rápidamente entran en la segunda división
meiótica, muy rápida y donde tiene lugar
la separación de las cromátidas hermanas.
Tras una corta interfase, con una duración
de minutos, sufren la meiosis II, completa
en 8h, y da lugar a dos espermátidas
haploides.
Espermátidas
Las cuatro espermátidas obtenidas a partir
del mismo espermatocito primario
permanecen conectadas entre ellas y
también aproximadamente a otras 100
espermátidas. Se localizan también cerca
de la luz de los tubos y son pequeñas y
alargadas. Su núcleo es claro y con un
nucléolo prominente. No se dividen, sino
que sufren un proceso conocido como
espermiogénesis.
La presencia de puentes va a permitir que la maduración sea paralela. Estas
espermátidas son el producto final de la espermatogénesis, haploides e inmóviles. Van a
219
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
ocupar posiciones centrales, con abundante REL, AG y RER. Las mitocondrias son
abundantes y tiene cuerpos RNA+ parecida al nuage. Son redondas y pequeñas, situadas
en nichos de células de Sertoli, para luego alargarse y situarse en las criptas de las
células de Sertoli.
Una vez maduras se liberan por espermiación, gracias al citoesqueleto contráctil apical
de células Sertoli.
Espermiogénesis
La espermiogénesis es la última fase de la espermatogénesis. Las espermátidas
maduras son liberadas hacia la luz del túbulo seminífero en un proceso llamado
espermiación (batido de los flagelos para que se desprendan provocado por las células
de Sertoli).
Se produce una reducción progresiva del tamaño del núcleo y una tremenda
condensación del material cromosómico asociada a la sustitución de las histonas por las
protaminas, conjuntamente a un aumento de la expresión génica y de la síntesis
proteica.
Fase de Golgi
Se inicia por la salida de vesículas densas del AG: gránulos preacrosómicos (PAS+).
Estas vesículas van a migrar vía microfilamentos y microtúbulos. Se va a producir
entonces la fusión con la vesícula acrosómica, situada cerca del núcleo, haciendo que
aumento su tamaño. Su posición determina el polo anterior del núcleo.
Los centriolos se desplazan al polo posterior. El distal y el perpendicular al núcleo
empiezan a formar el flagelo (axonema 9+2).
Fase de capuchón
El gránulo acrosómico crece y se extiende sobre el núcleo porque el AG sigue formando
gránulos.
Las histonas son sustituidas por protaminas ricas en Arg y Lis. La cromatina se
condensa y los gránulos son cada vez mayores. Van a desaparecer los nucléolos y los
poros de la envuelta. El núcleo se alarga.
El axonema sigue creciendo, cubierto por la membrana plasmática.
Fase de acrosoma
La espermátida se alarga y hay una clara poralidad celular. En esta etapa se va a
producir la reorientación: el polo anterior queda hundido en la cripta y el polo posterior
con el flagelo en crecimiento hacia la luz. La espermátida está formada en la cabeza por
el acrosoma y el núcleo, en el cuello por los centriolos y en la cola por el flagelo.
220
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Los centriolos retornan junto al núcleo, hundiéndose el centriolo proximal al
surco del núcleo. El cuello queda formado por fibras densas externas que conforman 9
columnas longitudinales alrededor del axonema.
La membrana se invagina donde está el acromosoma y rodea al núcleo (anillo nuclear o
de Jensen). Varios haces de microtúbulos rodean al núcleo y alcanzan los centriolos,
formando la vaina caudal o manguito. La mayor parte del citoplasma se desplaza hacia
el polo posterior y se elonga el núcleo.
La membrana vuelve a invaginarse en el origen del flagelo formando un anillo
membranoso: el anillo del flagelo. Las mitocondrias van a adquirir una disposición
helicoidal en la parte proximal del flagelo (zona media) y acaban en un anillo, que
delimita el límite de los segmentos medio y principal.
Fase de maduración
Las espermátidas tienen una constitución muy alargada y aún están unidas. Van
a sufrir una reducción del volumen citoplasmático.
Su núcleo es alargado y la cromatina alcanza su condensación máxima (x6). El
sistema de reparación del ADN es eliminado y las células son insensibles a agentes
mutágenos.
221
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
222
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
ESPERMATOZOIDE
Desaparecen fibras 3 y 8: 7+2
Es una célula haploide, de unas 60 micras. Se enuentra libre y es inmóvil hasta la
maduración. Tras esto, su movilidad es de 1-3 mm/min. Se distinguen cabeza y cola.
La cabeza se divide en segmentos apical, principal y caudal y contiene enzimas
hidrolíticas para la fecundación. La membrana acrosómica y la membrana nuclear se
encuentran unidas por el acroplaxomsa, formado por filamentos de actina F. Además,
existen vacuolas nucleares.
223
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
ORGANIZACIÓN CELULAR
En el túbulo seminífero hay diferentes estadios en desarrollo, no hay distribución
al azar. Hay agrupaciones debido a que las células derivadas de una gonia están unidas
por puentes y las células sincronizadas pasan por tiempos específicos para cada etapa
de maduración.
La asociación celular se refiere a una
organización fija y definida de tipos celulares
en determinados estadios evolutivos. En el
hombre hay 6 asociaciones (nombradas en
números romanos). Cada asociación celular
representa un estadio o etapa de un proceso
cíclico.
Un ciclo es una serie completa de asociaciones que se van sucediendo en el tiempo, que
se continúan unas con otras en un corte transversal de un túbulo seminífero. Implica
cambios en el tiempo en un punto. También se define como el tiempo necesario que una
secuencia de asociaciones celulares tarda en cambiar un punto concreto.
Las asociaciones celulares se suceden a lo largo del túbulo seminífero. Para que
se genere una espermátida madura se precisan varios ciclos. En el hombre hay 4-6
ciclos de 16 días (74 días).
Una onda es una sucesión de asociaciones celulares en la distancia. En la rata se
describen 12 ondas lineales en cada túbulo seminífero; en un corte transversal solo se
observa un estadio. En el hombre no se observan ondas porque son helicoidales; en un
corte transversal varios estadios.
224
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
TEJIDO INTERSTICIAL
Entre los túbulos seminíferos, contiene las células de Leydig y los macrófagos,
junto a vasos sanguíneos y linfáticos.
Células de Leydig
Aparecen en grupos, próximas a vasos. Son grandes, poligonales, con núcleo
redondo, claro y excéntrico y un citoplasma eosinófilo. Encontramos REL, mitocondrias
con crestas tubulares, lípidos, cristales de Reinke e interdigitaciones con macrófagos.
Aparecen en la 8ª-10ª semana a partir de la diferenciación de los fibroblastos en
respuesta a la hCG (placenta). Sintetizan y secretan andrógenos en el feto (semana 8-
18), neonato (2-3 meses) y con la pubertad (95% de testosterona). Tras el nacimiento
hay una regresión y parecen fibroblastos.
Van a responder a LH, que estimula la producción de testosterona. La prolactina
induce la expresión del Rc de LH. En caso de hiperprolactinemia se inhibe la función
reproductora masculina al reducir la secreción de gonadotropina; disminución de la
síntesis de testosterona, disminución de la espermatogénesis, disfunción eréctil con
infertilidad.
Se van a encargar de la síntesis y secreción de andrógenos,
x Testosterona: El 95% de la testosterona está en suero unida a la SHGB
(globulina transportadora de hormonas sexuales). El 5% se forma en la corteza
suprarrenal. La testosterona difunde al líquido testicular. Se fija a ABP y se
logran elevados niveles junto a células espermatogénicas en desarrollo.
x Dihidropiandrosterona (DHEA)
225
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Androstendionas
Los andrógenos van a tener distintas funciones a lo largo de la vida del varón:
x Feto:
o Regulación de la diferenciación de los genitales internos y externos de
tipo masculino.
o Estimulación del crecimiento, desarrollo y función de los genitales
internos y externos masculinos.
o Diferenciación del SNC.
x Pubertad:
o Estimulación de los caracteres sexuales secundarios: vello sexual, voz
grave, barba…
o Estimulación de la espermatogénesis.
o Estimulación de la secreción de las glándulas sebáceas.
o Unión a proteínas transportadoras de andrógenos (por células de Sertoli
tras FSH).
x Adulto:
o Mantenimiento de la espermatogénesis.
o Mantenimiento de los caracteres sexuales secundarios.
o Mantenimiento de las vías espermáticas y genitales externos.
o Mantenimiento de la función secretora de las glándulas accesorias.
o Conducta, incluido la líbido.
o Mtc gral (?), crecimiento del esqueleto, desarrollo muscular esquelético,
distribución del tejido adiposo subcutáneo, función renal.
226
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
227
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
VÍAS ESPERMÁTICAS
Origen embriológico
La
testosterona estimula el desarrollo del conducto mesonéfrico de Wolf. La parte proximal
al esbozo gonadal va a formar un conducto muy contorneado, el conducto epididimario.
Los conductos eferentes van a originarse debido al contacto de los túbulos excretores
mesonéfricos con los cordones testiculares. La porción distal va a originar el conducto
deferente, y el extremo más distal formará el conducto eyaculador y la vesícula seminal.
Vias intratesticulares
Túbulos rectos
Cerca del mediastino testicular los túbulos seminíferos van a dar paso a los túbulos
rectos, quedando entre ambos una porción formada solo por células de Sertoli.
Los túbulos rectos son muy cortos y tienen una luz muy estrecha. Están formados
exclusivamente por células similares a las de Sertoli que constituyen un epitelio simple
prismático bajo o cúbico, que presentan en su dominio apical (veamos que antes era en
el dominio basolateral) uniones occludens, microvellosidades y cilios ocasionales.
Rete testis
228
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Es una red laberíntica de canales anastomosados que se encuentra en el mediastino
testicular. Hallamos en su luz altas concentraciones de testosterona porque hay ABP,
que es una proteína ligadora de andrógenos,
esenciales para la función de la cabeza del
epidídimo. La concentración intraluminal de
andrógenos va a ser de hecho mayor a la
sanguínea.
La pared está formada por un epitelio cúbico
simple con algunas microvellosidades y un solo
cilio, similar al de los túbulos rectos. Por debajo
encontramos un conjuntivo con células mioides.
Destaca la abundancia de agregados de células de
Leydig y de canales linfáticos y vasos
sanguíneos.
Vías extratesticulares
Conductos eferentes
Se trata de 10-20 túbulos que conectan los conductos de la red testicular o de Haller en
el borde superior del mediastino testicular con la
porción proximal del conducto epididimario.
Conforme los conductillos eferentes abandonan el
testículo atravesando la túnica albugínea sufren un
enrrollamiento pronunciado formando de 6 a 10 masas
cónicas, los conos eferentes, cuyas bases son parte de
la cabeza del epidídimo y desembocan en el conducto
epididimario.
Los conductillos eferentes están tapizados por un
epitelio pseudoestratificado cilíndrico en el que hay
cúmulos de células altas y bajas, lo cual le da a la
superficie apical un aspecto festoneado o en diente de
sierra. Presenta varios tipos de células:
1. Células cilíndricas altas: son ciliadas. Los cilios baten hacia el epidídimo.
2. Células basales: actúan como células madre
3. Células principales del epitelio bajo: se trata de células no ciliadas que presentan
una gran cantidad de microvellosidades e invaginaciones canaliculares de la
superficie apical. Poseen una gran cantidad de vesículas pinocíticas y cuerpos
densos delimitados por membrana, lisosomas y otras estructuras citoplasmáticas,
relacionadas con la actividad endocítica.
La mayor parte del líquido secretado en los túbulos seminíferos se reabsorbe en los
conductillos eferentes.
229
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Por debajo del epitelio tenemos una capa de tejido conjuntivo con células mioides. Bajo
ella, aparece por primera vez en la vía espermática una capa de varias células de espesor
de músculo liso, que se dispone en forma de vaina circular en la pared del conductillo.
Entremezcladas con las células musculares hay fibras elásticas.
Los espermatozoides son inmóviles, por lo que son arrastrados por el líquido producido
en secciones anteriores. Sin embargo, se postula que contribuyen a ello la acción ciliar y
las células mioides.
Epidídimo
El epidídimo es un órgano de unos 7’5 cm de longitud que contiene tanto los conductos
eferentes como el conducto del epidídimo. Se trata de una estructura con forma
semilunar apoyada en la superficie superoposterior del testículo. Además de las
estructuras ya mencionadas, encontramos en él vasos sanguíneos, músculo liso y una
cubierta de tejido conjuntivo.
El conducto del epidídimo es un tubo que se origina por el conducto de Wolf gracias a
la acción de genes como OXA 10 y OXA 11. Resulta clave para el transporte,
maduración y almacén de los espermatozoides. Se encuentra muy enrollado, tiene una
longitud de entre 4 y 6 m, y en él se describe:
1. Cabeza: ocupada por los conductos eferentes
2. Cuerpo: ocupada por el conducto epididimario
3. Cola: ocupada por el conducto epididimario
230
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
La longitud de las distintas secciones varía enormemente En el epitelio del conducto
epididimario encontramos:
1. Células principales: su altura es mayor en la cabeza del epidídimo y disminuye
hacia la cola. Desde su superficie apical se extienden microvellosidades
modificadas largas e irregulares que reciben el nombre de estereocilios, cuya
altura sigue igual evolución de cabeza a cola. Presentan muchas vesículas
endocíticas y lisosomas, en general buen desarrollo de orgánulos, y su
membrana basal y basolateral se encuentra invaginada.
2. Células basales: son células madre redondeadas y apoyadas en la lámina basal
3. Linfocitos intraepiteliales, que reciben el nombre de células del halo.
Funciones del conducto epididimario
1. Absortiva: la mayor parte del líquido testicular (90%) que no es absorbido en los
conductos eferentes se absorbe en la porción proximal o cabeza del epidídimo.
2. Fagocítica: fagocitan espermatozoides con defectos o en degeneración o cuerpos
residuales que han escapado de las células de Sertolli.
3. Secreción de ácido siálico y carnitina, que inmovilizan a los espermatozoides
para permitir su maduración, y glucoproteínas y glicerofosforilcolina, las cuales
contribuyen a la maduración propiamente dicha, que ocurre en zona media o
cuerpo del conducto epididimario con la intervención fundamental de la
testosterona. Gracias al proceso de maduración el espermatozoide logra una
movilidad anterógrada y se estabiliza su cromatina. También se producen
cambios en las cargas y en los receptores de la membrana encaminados a que el
espermatozoide se pueda unir a la zona pelúcida del óvulo en una eventual
fecundación.
4. Almacén: Los espermatozoides ya maduros se almacenan en la cola. La
capacidad de almacenaje es limitada: si no se produce la eyaculación van a ir
degenerando.
5. Transporte: En la cabeza y la mayor parte del cuerpo del epidídimo hay una sola
capa delgada de músculo liso similar a la de los conductillos eferentes, pero en
la cola se añade una capa longitudinal interna y otra externa que se van a
continuar con las del conducto deferente. Ello sirve para explicar las diferencias
funcionales: en la cabeza y el cuerpo las contracciones peristálticas rítmicas
espontáneas sirven para mover los espermatozoides a lo largo del conducto,
mientras que en la cola, donde se produce el almacén, ocurren pocas
contracciones peristálticas mucho más potentes que tienen como fin empujar los
espermatozoides hacia el conducto deferente, y que ocurren durante la
estimulación sexual. La inervación de esta zona es adrenérgica, y resulta
fundamental la coordinación con la musculatura del conducto deferente.
Mutaciones de los genes que codifican para proteínas morfogenéticas óseas BMP4, 7 y
8 causan alteraciones en la formación y diferenciación de segmentos específicos del
conducto epididimario.
231
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Conducto deferente
Es el segmento más largo de la vía espermática, y presenta un diámetro pequeño debido
a su gruesa capa muscular. Asciende y se introduce en el abdomen como componente
del cordón espermático, y tras abandonarlo desciende en la pelvis hasta la altura de la
vejiga, donde su extremo distal se dilata para formar la ampolla, que recibe el conducto
de la vesícula seminal y continúa hasta la uretra a través de la próstata con el nombre de
conducto eyaculador.
El conducto deferente está revestido por un epitelio pseudoestratificado cilíndrico muy
parecido al del epidídimo, pero en el que la luz no es lisa y regular, sino que la mucosa
parece tener pliegues longitudinales profundos en la mayor parte de su longitud,
consecuencia fundamentalmente de la contracción de la capa muscular. En la parte
proximal hallamos estereocilios, que desaparecen después.
Presenta los mismos tres estratos musculares que la porción muscular del epidídimo,
aunque más desarrollados, especialmente el circular medio. Los miocitos adoptan una
disposición espirilada que permite la expulsión de los espermatozoides, dependiente
fundamentalmente de la capa muscular y su fuerte inervación adrenérgica.
232
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Antes de la eyaculación
por tanto tenemos que la
parte distal (deferente)
del conducto eyaculador
está cerrada, mientras
que su parte proximl
está abierta, con lo que
si no hay estimulación
se impide la salida.
Transporte de
espermatozoides
La eyaculación depende de movimientos rápidos y repetidos de aspirado-eyección, y es
posible gracias a la importante inervación de la capa muscular del conducto deferente.
Antes de la eyaculación, los conductos eyaculadores están cerrados. La contracción de
las capas longitudinales externa e interna del conducto deferente provoca la dilatación y
acortamiento del mismo, lo cual conduce a una disminución de la presión interna en el
conducto deferente que lleva al aspirado de los espermatozoides situados en el almacén
de la cola del epidídimo.
La eyaculación se inicia por tanto con la contracción de toda la muscular del conducto
deferente, coordinada con la contracción de la musculatura del epidídimo y la dilatación
del conducto eyaculador. Se produce entonces una elongación y estrechamiento del
conducto deferente que hace que aumente su presión interna y los espermatozoides sean
propulsados a la uretra prostática en un fenómeno que se conoce como emisión.
Conducto eyaculador
Es un tubo delgado de solo 2 cm situado dentro
de la próstata, que desemboca por medio del
colículo seminal en la uretra prostática. Recibe
tanto la secreción de las vesículas seminales
como los espermatozoides procedentes del
conducto deferente.
Su mucosa es delgada, y está constituida por un
epitelio prismático simple y una lámina propia
rica en fibras elásticas. El conducto eyaculador
solo presenta capa muscular en su porción
proximal.
Uretra
233
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Se trata de un conducto de 20 cm que podemos dividir en:
x Prostática: recibe la secreción de la próstata y del conducto eyaculador.
x Membranosa
x Peneana: recibe la secreción de las glándulas de Cowper.
En la uretra prostática encontramos urotelio, mientras que en la membranosa y en la
peneana o esponjosa hay un epitelio prismático estratificado. En ambos casos hay
glándulas mucosas intraepiteliales que se llaman glándulas de Littré, y se encargan de la
secreción de un líquido que protege frente a la orina.
A la altura de la uretra membranosa hay dos esfínteres, uno de músculo liso y otro
esquelético, encargados de regular tanto la micción como la eyaculación.
234
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
GLÁNDULAS ANEJAS
En el aparato genital masculino encontramos tres: próstata, glándulas de Cowper y
vesículas seminales.
Vesículas Seminales
Se forman durante el
periodo embrionario a
partir de la ampolla del
conducto deferente
(derivado del conducto de
Wolff) y recibieron ese
nombre porque se pensaba
que allí se almacenaban los
espermatozoides aunque no
es así.
Órganos pares, pequeños, siendo un conducto muy retorcido (15 cm de longitud cuando
está desenrollado) y que recuerda bastante al deferente aunque mucho menos en su
conjunto. El desarrollo de la glándula depende de la testosterona y su actividad secretora
es a partir de la pubertad (el epitelio es secretor).
En un corte apreciamos múltiples luces porque su mucosa tiene muchos pliegues que se
ramifican. El epitelio está formado por células prismáticas y basales y es ligeramente
variable, siendo prismático simple o pseudoestratificado y depende de la testosterona
para mantenerse (habiendo aplanamiento de sus células prismáticas por ejemplo en la
vejez). Consta de muchos orgánulos (RER, AG, inclusiones de lipofucsina…) y tiene
función secretora por lo que aparecen muchos gránulos eosinófilos apicales. La imagen
típica de estas células es de apocrinia (a partir de la pubertad).
Se apoya sobre una lámina propia laxa y delgada con una importante capa muscular de
dos estratos: uno circular interno y otro longitudinal externo (aunque a veces
encontramos tres). Finalmente encontramos una adventicia.
Las vesículas seminales forman una secreción alcalina muy espesa y eosinófila que
forma la mayor parte del esperma (60-80%). Dentro del eyaculado esta secreción sale al
final (es decir, el eyaculado va por tramos y esta es la parte final) y contiene fructosa,
prostaglandinas, aminoácidos, potasio y flavinas (útiles en medicina legal porque son
fluorescentes)
La función principal de esa secreción es la nutrición de los espermatozoides y de la
coagulación del semen (por parte de algunas proteínas únicamente).
La secreción se drena por conductos cortos, entre el deferente y los eyaculadores, que
atraviesan la cápsula de la próstata metiéndose dentro para llegar al conducto
eyaculador. Cuando se produce la eyaculación además de la contracción del deferente y
del epididimario ocurre también la de los miocitos de la pared de la vesícula lo que
permite expulsar la secreción de las vesículas a la parte final del eyaculado.
235
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Próstata
Es una glándula impar con forma de castaña. Es la mayor de las glándulas accesorias.
Se caracteriza por tener un estroma en cuyo TC hay una gran cantidad de miocitos,
siendo entonces palpable la glándula por su gran consistencia. En su interior circula la
uretra prostática y los conductos eyaculadores.
El estroma no solo está dentro sino que tiene una importante cápsula fuerte, firme y
consistente, no especialmente gruesa de la cual parten tabieques que van al interior. En
el parénquima tenemos unas 30-50 glándulas tubuloalveolares que drenan por unos 15-
20 conductos excretores independientes al seno prostático uretral (al colículo seminal).
Hay algunas que se abren directamente a la uretra (por eso encontramos más glándulas
que conductos).
236
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Anatómicamente, la próstata tiene 4 regiones más o menos concéntricas entre sí:
x Región periférica: es de mayor tamaño (70%) o volumen y es importante desde
el punto de vista clínico porque y es la que se inflama y maligniza con mayor
facilidad/frecuencia (formación de carcinomas).
x Zona central: es más resistente a las reacciones inflamatorias y malignización
(prácticamente nunca). Es donde se incluyen los conductos eyaculadores, y tiene
un origen embriológico diferente.
x Transicional.
x Periuretral.
La periférica y la central son las más grandes. La zona transicional y periuretral también
tienen importancia clínica porque con la edad las células proliferan en exceso
produciéndose la Hiperplasia Benigna Prostática (HBP). Se forman nódulos que
comprimen la uretra y dificultan la micción (retención de orina, cistitis, pielonefritis…).
Esto se produce porque las células del estroma, no las del parénquima, contienen 5 alfa-
reductasa que forma dihidroxitestosterona a partir de la testosterona. La DHT estimula
la formación de factores de crecimiento mitogénicos para células estromales y
epiteliales. Esta patología se trata con inhibidores de la 5 alfa-reductasa.
Histológicamente, hablamos de 3 regiones concéntricas en la próstata, en función del
tipo de glándulas:
x Periférica (glándulas principales)
x Submucosa (glándulas submucosas)
x Mucosa (glándulas mucosas)
Tenemos tres zonas concéntricas próstata, y en el centro del todo vemos la uretra. Todas
las glándulas mucosas drenan a la uretra directamente mientras que las principales y
submucosas tienen conductos excretores más largos en los senos prostáticos junto a las
crestas uretrales. El epitelio se inicia siendo prismático simple pero en la uretra acaba
siendo de transición o urotelio.
La luz de la próstata presenta los cuerpos amiláceos (1-2mm de tamaño) que son
eosinófilos y visibles con estructura laminar concéntrica. Se forman cuando, alrededor
237
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
de algún resto, se precipitan elementos de la secreción prostática (glucoproteínas),
debido a las concentraciones de amiloide. En estas masas hay glicoproteínas y calcio
que precipitan alrededor de restos celulares son los cálculos o concreciones que
aumentan con la edad pero que no son nada patológico.
Las glándulas son túbulo-alveolares de trayecto contorneado, luz amplia y adenómero
variable (diminuto, estrecho, amplio). El epitelio de estos adenómeros es un epitelio
(liso, con pliegues…) de tipo pseudoestratificado que puede estar o muy replegado o
muy distendido. Encontramos células secretoras cuya altura depende de la actividad (a
más actividad más altura) la cual, a su vez, está regulada por la testosterona.
Antes de la pubertad solo tenemos conductos excretores (como en la mama) y las
unidades secretoras se elevan con la pubertad que estimula al epitelio. Por tanto, los
botones del epitelio crecen, forman los alveolos y hacen que el tamaño de la glándula
aumente. Así en la pubertad el tamaño de la glándula llega a duplicarse.
Se describen 3 tipos celulares en el epitelio:
x Principales o secretoras: presentan abundante RER y aparato de Golgi debido a
su función secretora. Tienen también lisosomas con fosfatasa ácida específica o
prostática FAP (se usa como marcador). Forman una secreción muy fluida,
alcalina y lechosa que forma parte de la primera porción del eyaculado (la
primera es muy fluida y la segunda no). Contiene altos niveles de FAP, PSA que
es un antígeno prostático específico, enzimas, fibrinolisina, poliaminas en
medicina legal, prostaglandinas y ácido cítrico (aunque recordemos que es una
secreción alcalina) proteasas, amilasas, inmunoglobulinas y zinc.
x Basales: encargadas de la renovación (son células madre).
x Endocrinas: sintetizan serotonina pero no está clara cuál sería la función de este
neurotransmisor (contráctil tal vez). El epitelio se asienta sobre una LP fina que
se asienta sobre el estroma (rico en conjuntivo y con miocitos)
238
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
La PSA es la molécula usada para la detección precoz del carcinoma de próstata. Sus
niveles circulantes son bajos y proceden del hígado y cuando se elevan dan indicios de
un posible carcinoma de próstata que es el más abundante (70% varones mayores de 70
años). Esos niveles se elevan por la hiperactividad de las glándulas prostáticas
principales. Supuso un enorme avance ya que el cáncer prostático era asintomático y se
detectaba muy tarde.
Esta secreción neutraliza el pH acido de la vagina, aporta nutrientes, estabiliza la
cromatina y aporta componentes bacteriostáticos. Hace que el semen más líquido. La
próstata como glándula solo es activa desde la pubertad. Hasta entonces está
indiferenciada: es más pequeña y apenas se observan conductos. El desarrollo de
epitelio y las glándulas es posible al elevarse los niveles de testosterona, momento en
que se desarrollan los adenómeros y las células secretoras.
Glándulas de Cowper
Son glándulas pares, pequeñas, situadas detrás de la uretra membranosa y tienen un
conducto excretor (relativamente largo para el tamaño de la glándula) que desemboca en
la uretra peneana en su porción más proximal, llegando directamente a la uretra del
cuerpo esponjoso. Son glándulas tubuloalveolares de secreción mucosa tapizadas por
epitelio prismático simple, con un conducto excretor tapizado por un epitelio prismático
simple (el mismo).
La secreción de estas glándulas es un líquido claro, que precede a la emisión del semen
y es mucosa. El pH alcalino de esta secreción neutraliza el pH ácido de la orina y actúa
como lubricante de la uretra esponjosa/ peneana. Contiene sobre todo galactosa y ácido
siálico, aunque también encontramos galactosamina, ácido galactourónico y
metilpentosa.
Semen
Es una secreción alcalina, lo que le permite neutralizar el pH ácido de la uretra y el del
medio vaginal. El semen contiene líquido testicular (que aunque hemos absorbido
mucho sigue quedando), secreciones del epidídimo, del conducto deferente, la próstata,
las vesículas seminales y las glándulas de Cowper (también había células de los
conductos que contribuían a estas secreciones). El 10% del semen corresponde a
espermatozoides que se encuentran en una concentración de 100 millones por mililitro.
Además en el semen encontramos también un número importante de células
descamadas. El análisis del semen se lleva a cabo mediante el espermograma.
Se considera que cuando el número de espermatozoides disminuye a 20 millones/ml, es
signo de infertilidad a causa de oligospermia. Aunque el volumen sea normal, no
significa que los espermatozoides sean sanos. En individuos sanos, el 20% de los
espermatozoides eyaculados son anormales (con algún tipo de deficiencia) y el 25% de
estos son inmóviles.
Además estos gametos pueden almacenarse en nitrógeno líquido a bajas temperaturas lo
que no afecta su capacidad de fertilización. Esto permite la criopreservación que
posibilita la inseminación artificial y la FIV (fecundación in vitro) con semen
congelado.
239
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
PENE
Es el órgano copulador masculino. Está recubierto por piel fina pigmentada, deslizable y
sensitiva. La dermis cuenta con músculo liso, interior longitudinal y exterior circular.
Estos miocitos se continúan con el dartos del escroto. El escroto no tiene ni vello ni
folículos pilosos.
En la piel de la raíz peneana sale vello pubiano. La piel del pene se asienta sobre un
conjuntivo muy fino y laxo con adipocitos. Su parte distal es el glande, cubierto por una
piel deslizable, el prepucio. En él hay unas glándulas sebáceas llamadas de glándulas de
Tyson que secretan esmegma.
Esta región tiene una alta inervación sensitiva y somática. Corre tanto a cargo de fibras
simpáticas como parasimpáticas y logra la erección y eyaculación. Está formado por 3
estructuras de forma cilíndrica:
x Cuerpos cavernosos: dos en la cara dorsal
x Cuerpo esponjoso: uno en la cara ventral.
Son cuerpos de tejido eréctil, que se encuentran rodeados por una cápsula de tejido
conjuntivo muy fibroso.
x Cuerpos cavernosos: Reciben ese nombre porque al observarlos hay muchos
espacios irregulares y anastomosados: cavernas. Las cavernas están revestidas de
endotelio, por lo que son espacios vasculares. Entre dos cavernas queda un
estroma con una importante cantidad de tejido conjuntivo y es característico que
en estas trabéculas de estroma haya muchos miocitos.
Los espacios (cavernas) más grandes tienden a situarse en la zona central. En la
periferia son más pequeñas y están en continuidad con un plexo venoso. Se encuentran
rodeados por la túnica albugínea, muy gruesa y fibrosa. En la zona media las túnicas
albugíneas que rodean los cuerpos se fusionan en un tabique. La sangre ha de atravesar
las túnicas albugíneas para entrar en las cavernas.
x Cuerpo esponjoso: Similar, pero la túnica es más delgada y más elástica.
También encontramos un plexo venoso en la zona periférica. Las cavernas
disminuyen en tamaño y en número en el extremo terminal (glande). Aquí se
sitúa la uretra peneana. Gran desarrollo de musculatura. Las cavernas están
llenas de sangre procedente de arterias helicinas, que describen una trayectoria
helicoidal.
En estado de flacidez, las cavernas están colapsadas en los cuerpos cavernosos pero sí
tienen sangre en el cuerpo esponjoso.
En estado de erección todas las cavernas se llenan de sangre. Cuando esto sucede, las
cavernas comprimen las venas de drenaje: llega sangre pero no puede ser drenada.
Como consecuencia, los cuerpos cavernosos aumentan mucho de tamaño. El esponjoso
también crece, aunque no tanto.
Las arterias helicinas son ramas de la arteria profunda medial de cada cuerpo y son las
que permiten o no el paso de sangre a los cuerpos cavernosos. Estas arterias tienen una
túnica media muy desarrollada, con miocitos de inervación parasimpática. No solo hay
miocitos en la media, sino que también se encuentran en la íntima en forma de
almohadillas (propulsión), dispuestos longitudinalmente. Los miocitos de la media
240
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
están muy regulados, y responden a dos controladores: PS y NO. Cuando están abiertas
se permite el paso de sangre a las cavernas.
El drenaje en los cuerpos cavernosos se produce por medio de vénulas postcavernosas
que dan paso a un plexo venoso periférico. Este plexo tiene paredes muy finas, y
atravesará la túnica albugínea para permitir el drenaje por la vena dorsal profunda. En el
cuerpo esponjoso es similar, pero drenado por la vena dorsal superficial. También hay
anastomosis arteriovenosas que permiten el paso directo de la sangre de arterias al
sistema venoso: de ahí que las cavernas están perfundidas o no.
Erección
El paso de la sangre a las cavernas es un proceso vascular que se inicia con estímulos
sensitivos. La erección es la respuesta del SNC ante un estímulo sexual.
Provoca que el SNPS actúe sobre los miocitos tanto de las arterias helicinas como los
del estroma entre las cavernas, relajándolos. Esto facilita que la sangre que no pasaba a
los cuerpos e iba por las arterias, llegue a las cavernas, se llene, y dé la erección. El SNS
controla la emisión de esperma y el cierre del cuello vesical.
El SNPS libera ACH que actúa estimulando células endoteliales. Éstas podían activarse,
con lo que sintetizan y segregan NO. Es un factor que actúa sobre miocitos, activando la
guanilato-ciclasa, que estimulada forma GMPc.
La estimulación de GMPc acaba por rebajar los niveles de calcio libre por el citoplasma.
La bajada de calcio es esencial para la relajación muscular, al no haber Calcio
disponible para la contracción. Los miocitos, tanto los trabeculares como los de las
arterias helicinas están unidos entre sí por uniones GAP que son esenciales para
propagar el efecto del NO sobre cada una de las células, ya que no llega a todas ellas.
Cuando la sangre llega al cuerpo cavernoso se estanca por compresión del drenaje. En el
esponjoso el aumento de tamaño no es marcado, ya que no se debe permitir la oclusión
de la uretra.
La erección acaba cuando la fosfodiesterasa elimina el GMPc en formación. Cuando sus
niveles bajan claramente, la musculatura comienza a contraerse y aumenta entonces el
tono en las arterias y en las trabéculas (debido a la liberación de Calcio). Esto supone
que la sangre pueda seguir entrando. Disminuye el paso de sangre a las cavernas, y
como se está contrayendo, se permite el drenaje sanguíneo.
241
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Clínica: Disfunción eréctil
Multicausal (puede ser por traumatismo, Parkinson, ictus, dabetes…). Provoca
incapacidad para mantener una erección que permita mantener un coito. Cuando no hay
afección del SNPS se puede tratar con (sildenafil) viagra. Éste actúa sobre la
fosfodiesterasa, que no elimina el GMPc, potenciando el efecto del NO y alargando la
erección (manteniendo así los niveles de GMPc elevados). Esto ocurre siempre y
cuando no haya habido una afectación del SNSP.
Los procesos de erección y orgasmo también tienen parte Simpática importante. Si ha
habido secciones medulares afectando al PS todavía se permite que tengan erecciones a
pesar de todo.
ESCROTO Y TESTÍCULOS
Estructura par. Bolsa de piel cuya cara interna está revestida por la túnica
vaginal. En su interior se aloja un testículo, gónada que ha descendido por el conducto
inguinal durante el desarrollo embrionario, y pesa 20 gr.
Los testículos se alojan fuera de la cavidad abdominal (extraabdominales). Su
cara posterior está adosada al epidídimo. Dentro del escroto, el testículo está sostenido
por el cordón espermático.
El descenso testicular se produce durante el periodo embrionario y tiene dos
fases: transabdominal e inguinoescrotal. Si no se produce el descenso adecuado ocurre
242
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
la criptorquidia. Es relativamente frecuente, especialmente en bebés prematuros. Es
asintomático. Si el testículo permanece en la cavidad abdominal hay una temperatura
demasiado elevada, lo que inhibe la espermatogénesis. Puede conllevar a esterilidad si
es bilateral y no se detecta. En las criptorquidias no tratadas hay elevada incidencia de
tumores.
Proceso de descenso gonadal
x Fase transabdominal: Está regulada por las células de Sertoli, que segregan el
factor MIS (sustancia inhibidora mulleriana), clave. En su ausencia (o de su
receptor) no se produce el descenso. No es el único factor implicado, ya que las
células de Leydig segregan INSL3, que actúa sobre miocitos desencadenando
unas señales imprescindibles en esta fase.
x Fase inguinoescrotal: Es dependiente absolutamente de los niveles de
andrógenos. Está inducida por CGRP, relacionado con a calcitonina. Junto al
testículo, en el cordón espermático está el plexo venoso pampiniforme. Este
plexo es el encargado de disipar calor mediante un mecanismo contracorriente.
Cuando la temperatura externa es demasiado baja, hay un reflejo de los
músculos cremáster y dartos, produciendo su contracción y acercando los
testículos a la pared abdominal. (Para la espermatogénesis se requiere una
temperatura inferior a la corporal)
Origen embriológico
Está en estrecha relación con la formación del aparato urinario. Ocurre en la
cavidad abdominal.
El mesodermo intermedio se encarga de formar las crestas urogenitales,
tapizadas por epitelio plano llamado mesotelio celómico.
Células germinales que no forman parte ni de las crestas ni del mesotelio, sino
que vienen de fuera (del saco vitelino) y pueblan las gónadas en desarrollo, y forman las
espermatogonias. Estas células germinales proliferan e inducen a las células del
mesotelio celómico y las crestas a que también se dividan. Dentro de la gónada, se
243
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
desarrollan unos cordones sexuales que acaban siendo cordones testiculares. Desde
estas células se forman los túbulos seminíferos y rectos y la rete testis.
Estructuras del cordón espermático
x Conducto deferente.
x Arteria testicular.
x Arteria del conducto deferente.
x Arteria cremastérica.
x Plexo pampiniforme.
x Fibras nerviosas simpáticas.
x Nervio genitofemoral.
x Vasos linfáticos.
Vascularización del testículo
x Arteria testicular (aorta abdominal). Ramas helicoidales.
x Arteria cremastérica (epigástrica inferior).
x Arteria deferencial (vesical inferior).
x Plexo venoso pampiniforme: mecanismo contracorriente intercambiador de
calor.
244
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
APARATO GENITAL FEMENINO
FUNCIONES
x Producción gametos femeninos
x Recepción de gametos masculinos
x Entorno adecuado para fecundación
x Entorno adecuada para la implantación
x Nutrición del embrión/feto
x Expulsión de feto maduro
x Producción y secreción de hormonas
CARACTERíSTICAS
Inicialmente las gónadas están indiferenciadas. El desarrollo puede ser hacia ovario (en
ausencia del factor determinante de testículo y de factor inhibidor mülleriano) o hacia
testículo, produciéndose el desarrollo subsiguiente de las vías y genitales externos Sufre
cambios estructurales desde la infancia hasta la madurez y luego va sufriendo cambios
cíclicos hasta la menopausia de la mujer. Este desarrollo se va completar en la pubertad,
momento a partir del cual tienen lugar distintos hechos:
x Menarca: Primera menstruación. No tiene una edad fija, entre 9-16 años. Estos
variaciones son perfectamente normales.
x Telarca: Desarrollo de la mama femenina. Suele tardar en producirse dos años a
partir de la menarca.
x Pubarca: Desarrollo del vello pubiano.
x Adrenarca: Desarrollo del vello axilar.
Cada 28 días hasta que llega a los 50 años hay un ciclo menstrual, a partir de este
momento se produce el climaterio: la regla se mantiene pero hay ciclos irregulares. Tras
ella tiene lugar una lenta atrofia de los órganos reproductores durante la posmenopausia.
COMPONENTES
x Genitales externos:
o Monte de venus
o Vulva: labios mayores y menores
o Clítoris
x Genitales internos:
o Ovarios
o Oviductos o trompas de Falopio
o Útero
o Vagina
245
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
DESARROLLO
El desarrollo es igual que en el masculino porque comenzamos con unas células
germinales que parten en la tercera semana de vida, migran e invaden el blastema
gonadal que se está formando y que era indiferenciado.
A partir de la cuarta semana hasta la sexta migran, a la vez que proliferan, a lo largo
del mesenterio dorsal del intestino posterior mediante movimientos ameboides.
Encontramos entre 2500 y 5000 células que pueblan el blastema gonadal (el
mesénquima) y que inducen la proliferación del epitelio celómico.
Este proceso migratorio es posible gracias a la interacción del factor de célula madre
con el receptor c-Kit. Si el ligando o el receptor faltan, tenemos un déficit de células
germinales primordiales, lo cual afecta a la hematopoyesis, desarrollo de melanocitos y
mastocitos.
Es decir, que a partir del 2º mes, las células germinales se encuentran a la altura de la
décima vértebra dorsal, donde inducen la proliferación de células del mesonefros y el
epitelio celómico (segregando factores que inducen a ello). Las células del mesonefros
revestidas por celoma formarán el blastema gonadal, que es una estructura
indiferenciada (cordones que vienen del mesonefros revestidos de epitelio).
Las células que se meten en el blastema reciben el nombre de ovogonias. Estas células
se sitúan en los cordones que aquí se llaman cordones sexuales. Estos primeros
cordones que se forman comienzan a atrofiase y a degenerar a partir de la 7º semana de
vida.
Después, nuevas células invaden el epitelio celómico y se asocian con las ovogonias de
la gónada, rodeándolas y formándose así cordones sexuales secundarios (lo que no
ocurre en el caso masculino). Las ovogonias de estos cordones entran en la primera
división meiótica por lo que se denominan ovocitos de primer orden, y esta división se
246
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
detiene en la primera profase. Así, tenemos
una gónada que está diferenciada en
femenina pero los ovocitos están detenidos
en primer orden en la profase I.
En el caso masculino los cordones
testiculares se formaban a partir de la zona
medular y la cortical involucionaba. En
cambio, en el caso femenino es al contrario
por lo que los cordones se forman de la
corteza de la gónada indiferenciada. Estos
cordones sexuales son los que van a
originar el ovario.
En este caso, al no haber sustancia
antimülleriana, los cordones de Müller se desarrollan. Durante el desarrollo, los
extremos craneales de los conductos de Müller siguen separados para formar los
oviductos, que desembocan en la cavidad celómica (la futura cavidad peritoneal). Los
segmentos caudales de estos conductos se fusionan para formar el primordio
uterovaginal, que posteriormente se convierte en el útero y la parte superior de la
vagina. Los ligamentos anchos del útero, derivados de dos pliegues peritoneales, se
aproximan entre ellos cuando los conductos de Müller se fusionan.
La cloaca primitiva se divide por el tabique urorrectal en dos regiones: el seno
urogenital ventral y el conducto anorrectal dorsal. El tabique urorrectal se fusiona con la
membrana cloacal (el lugar de futura localización del cuerpo perineal), que se divide en
la membrana anal dorsal y la membrana urogenital ventral, de mayor tamaño.
A la semana 7, estas membranas se rompen. El contacto del primordio uterovaginal con
el seno urogenital determina la formación de la lámina vaginal. La canalización de esta
lámina vaginal se traduce en la aparición de los tercios medio e inferior de la vagina:
El seno urogenital también da lugar a la vejiga urinaria, la uretra, las glándulas
vestibulares y el himen.
Síndrome de Rokitansky-Kuster-Hauser o agenesia de Müller: en donde no se va a
formar el útero, cérvix y tercio superior vaginal. Sin embargo el ovario y la ovulación se
producen con normalidad. Con cierta frecuencia se produce la falta de riñón también
entre otras cosas.
A la semana 4 se desarrolla el tubérculo genital o falo en el extremo craneal de la
membrana cloacal. Posteriormente se desarrollan las tumefacciones labioescrotales y los
pliegues urogenitales a ambos lados de la membrana cloacal.
El tubérculo genital se hipertrofia tanto en el varón como en la mujer. Cuando no hay
andrógenos, los genitales externos se feminizan: el falo se convierte en el clítoris. Los
pliegues urogenitales dan lugar a los labios menores y las tumefacciones labioescrotales
evolucionan a labios mayores.
247
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
GENITALES EXTERNOS
El himen imperforado se debe a canalización incompleta de la lamina vaginal: Realizar
himenotomia.
Monte de Venus
Prominencia en la sínfisis del pubis. El monte de Venus corresponde a piel revestida por
epitelio escamoso estratificado queratinizado con folículos pilosos oblicuos (a partir de
la pubertad), que cubre la grasa subcutánea que reviste la sínfisis del pubis. El pelo es
ensortijado en todas las razas.
Clítoris
Se localiza debajo del monte de Venus, es el equivalente al pene en la mujer. Al igual
que el pene, está constituido por dos cuerpos cavernosos laterales (tejido vascular
eréctil) separados por un tabique incompleto y rodeados de una vaina de colágeno
fibroso. El clítoris se recubre parcialmente de piel fina con abundantes nervios
sensitivos y receptores, pero que no tiene folículos pilosos ni glándulas.
Su crecimiento se produce tras la pubertad. Durante la excitación sexual va a hincharse.
Los dos cuerpos cavernosos acaban formando el glande.
Vulva
Es un pliegue cutáneo par situado bajo el Monte de Venus y equivalente al
escroto masculino.
Los labios mayores son extensiones del monte de Venus a ambos lados del introito
vaginal. El músculo liso en los labios mayores va a separar las caras internas y externas:
x Cara externa: Vello, glándulas sebáceas y sudoríparas ecrinas y apocrinas. Tras
la pubertad se pigmenta. Posee abundante grasa subcutánea y una capa de
músculo liso equivalente al dartos del escroto.
x Cara interna: Lisa, recta y lampiña, con glándulas sebáceas y sudoríparas.
Los labios menores son un par de pliegues subcutáneos más finos que están debajo de
los mayores. Delimitan un espacio denominado vestíbulo. Están tapizados por epitelio
plano estratificado, con gran riqueza en papilas. Carecen de vello y su tejido conjuntivo,
su dermis, es rica en fibras elásticas y en glándulas sebáceas. No encontramos ni tejido
muscular ni grasa subcutánea. Además destaca en estos labios una muy abundante
vascularización. La cara externa está más pigmentada.
Vestíbulo vaginal
El vestíbulo vaginal se corresponde al espacio entre los labios menores, al que se abren
la uretra, la vagina y los conductos de las glándulas vestibulares mayores y menores.
Glándulas
Glándulas sebáceas y sudoríparas en Monte de Venus y vulva. Las glándulas
vestibulares pueden ser:
248
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Menores, parauretrales o de Skene: Sobre el clítoris y alrededor de
desembocadura de la uretra. Recuerdan a las glándulas de Littré de la uretra
masculina. Tienen epitelio prismático pseudoestratificado y son de naturaleza
mucosa.
x Mayores, vulvovaginales o de Bartholin: A ambos lados del vestíbulo, en parte
la inferior de vagina. Son túbuloalveolares mucosas. Sus conductos tienen
epitelio prismático estratificado. Se abren en cara interna de labios menores,
cerca del orificio vaginal.
Su función es lubrificar la entrada de la vagina, en especial en la excitación sexual.
Inervación de los genitales externos
Están tremendamente inervadas, sobre todo sensitivas. Gran cantidad de terminaciones
libres. Muchos corpúsculos tanto de Meissner (monte de Venus y labios mayores) como
de Paccini (labios mayores y cuerpos cavernosos del clítoris). Son esenciales para la
respuesta fisiológica en la excitación sexual.
Himen
Hay una membrana, una mucosa muy fina que ocluye el paso a la vagina tiene dos
caras: cara vulvar (externa), tapizada por epitelio plano estratificado queratinizadao, y la
cara vaginal (interna), que no está queratinizada. Entre los dos epitelios hay conjuntivo
vascularizado y por eso cuando se rompe el himen hay sangrado. (ella lleva tanto sin
follar que le ha vuelto a crecer el himen).
OVARIO
Órgano par e intraperitoneal, suspendido por ligamentos y por el mesovario. Antes de la
pubertad es un órgano con superficie lisa y con el paso del tiempo va cogiendo forma
irregular y forma cicatrices.
Funciones
x Oogénesis: Producción de gametos femeninos.
x Esteroidogénesis: Forma hormonas esenciales para el ciclo menstrual.
o Los estrógenos son esenciales para promover el crecimiento y
maduración de genitales externos e internos. Es responsable del
desarrollo de caracteres sexuales femeninos y estimulan el crecimiento
de conducto y estroma de glándulas mamarias. Lo estrógenos son clave
en la distribución del tejido adiposo de la mujer.
o Forman progestágenos y estos son esenciales para preparar el útero y la
glándula mamaria para el embarazo y la lactancia resptivamete.
Hormonas esteroides (derivados del colesterol)
x Estrógenos:
o Estradiol: Sintetizada por la granulosa y las células luteínicas foliculares.
Predomina a partir de los 50 años en la perimenopausia y en la
menopausia. Es el estrógeno más potente y responsable de:
Caracteres sexuales secundarios
249
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Crecimiento uterino
Engrosamiento de la mucosa vaginal
Moco vaginal acuoso
Desarrollo de glándulas mamarias
Evitan enfermedades cardiovasculares
o Estrona: Predomina en la postmenopausia
x Progesterona: Hormona sintetizada por la granulosa y el cuerpo lúteo que
predomina en la segunda mitad el ciclo femenino. Sus funciones son:
o Prepara el útero para una posible implantación (fase secretora o
progestacional)
o Inhibe contracciones uterinas
o Aumenta la viscosidad del moco cervical
o Intensifica el desarrollo de las glándulas mamarias
o Eleva la temperatura corporal basal
x Andrógenos: Las células de la teca interna segregan andrógenos, que son
transformados por la granulosa en estrógenos. ¨Tienen acción masculinizante en
la mujer.
o Androstendiona
o Dehidroepiandrosterona
Otras hormonas
Hay otras hormonas como la inhibina, la activina y la relaxina, además de la oxitocina y
la vasopresina.
Clínica
Sindrome de Turner: Falta un cromosoma X o el segundo cromosoma X no está
completo, pero en concreto se forman cintillas ováricas. Además hay una talla baja,
alteraciones esqueléticas y una producción nula o baja de estrógenos que está asociada
con altas concentraciones de gonadotropinas que desemboca en una falta de desarrollo
sexual secundario.
Ciclo
El ovario durante la etapa fértil presenta un ciclo en el que hay 3 fases:
x Folicular: Desarrollo del folículo primordial a terciario.
x Ovulatoria: Ovulación.
x Luteínica: Folículo que ha ovulado se transforma en cuerpo lúteo, una glándula
endocrina por í mismo.
Estructura histológica
Un ovario esta formado por un epitelio que lo delimita y que se llama epitelio
germinativo. Bajo este hay una zona de tejido conjuntivo que recibe el nombre de túnica
albugínea. Debajo está la corteza y después la médula. El ovario también tiene un hilio
por donde entran y salen los vasos y nervios, unido por el mesovario al ligamento
ancho.
250
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Epitelio germinativo
El epitelio germinativo se creía que era el encargado de formar los ovarios.
El epitelio germinativo es plano simple en la infancia y a lo largo de la pubertad se va a
hacer cúbico simple, en algunas partes casi plano. Estas células no tienen un gran
desarrollo, pero tienen muchas vesículas de pinocitosis, microvellosidades, algunos
cilios apicales y mitocondrias. Este epitelio secreta líquido.
Durante la ovulación se rompe, por lo que pueden formarse islotes dentro de la corteza,
conocidos como quistes de inclusión.
Túnica albugínea
Capa de conjuntivo bastante denso, con fibras situadas en paralelo al epitelio. El nombre
de esta parte de debe a que la matriz del conjunto tiene una MEC bastante blanquecina
porque no hay apenas vasos.
Corteza
La estructura principal es el folículo. Hay, entre ellos, tejido intersticial En el tejido
intersticial hay un conjuntivo (estroma). A partir del estroma se diferencia la teca
(conjunto que se incorpora a la estructura de folículo) y después la glándula intersticial
y las células hiliares que no tienen que ver con el folículo. Su límite con la médula no es
nítido.
El estroma es tejido conjuntivo donde predominan las fibras reticulares. Podemos
encontrar mocitos, no muy abundantes, y muchos vasos asociados a fibras elásticas
situados alrededor de folículos en desarrollo, atrésicos y de cuerpos lúteos.
A partir del estroma se diferencia alrededor de los ovocitos la teca para formar
parte del folículo. Cuando se desarrolla se divide en interna (en contacto con la
granulosa, secretora y muy vascularizada) y la externa (se continúa con el estroma
intersticial y es fibrosa).
Las células luteínicas, que quedan libres en el estroma tras la involución del cuerpo
lúteo, pueden conservar su actividad secretora y formar las denominadas glándulas
251
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
intersticiales (proceden de la teca hipertrofiada de folículos atrésicos, secundarios).
Estas células glandulares intersticiales no son abundantes en el ovario humano. Están
formadas por acúmulos dispersos de células epitelioides. Estas glándulas intersticiales
contienen vesículas lipídicas. Tienen carácter secretor y se encargan de la producción de
andrógenos (estrógenos). Son más abundantes en la infancia e involucionan tras la
menarca.
Las células hiliares son histológicamente idénticas a las células de Leydig. Se
encuentran en la corteza, pero son muy abundantes cerca del hilio y del mesovario. Son
células de carácter epitelioide y bastante grande. Su citoplasma es espumoso y está
lleno de inclusiones lipídicas. También hay esteres de colesterol, pigmento lipocrómico
y cristales de Reinke. Son prominentes en gestación y menopausia y se encargan de la
producción de andrógenos.
Estas células hiliares pueden sufrir hiperplasia o malignización produciendo
masculinización.
Folículos
Un folículo es una estructura en la que hallamos una célula germinal femenina y células
acompañantes, esenciales para la maduración de la célula germinal, y sin las cuales no
podría haber ovulación. El grado de desarrollo de la célula germinal y las acompañantes
y, en consecuencia, del folículo, son variables. En general, clasificamos los folículos en:
x Unilaminares: una sola capa de células acompañantes o foliculares
x Multilaminares: varias capas de células acompañantes o foliculares
Folículos primordiales
Son los más pequeños, no superan las 40 micras, en parte porque son siempre
unilaminares. Su situación en el ovario es periférica, los encontramos por debajo de la
túnica albugínea. Están constituidos por un oocito primario y una sola capa de células
foliculares epiteliales planas, aunque en ocasiones podemos encontrar folículos
poliovulares (con 2 oocitos).
Oocito
Se trata de un oocito primario detenido en diplotene de la profase I meiótica. Su
posición en el folículo es central y su tamaño no supera las 30 micras.
Microscópicamente distinguimos tres características fundamentales:
1. Núcleo: grande, excéntrico, con un nucléolo.
2. Cuerpo de Balbiani: se trata de una estructura celular compuesta por un gran
dictiosoma que constituye el aparato de Golgi, mitocondrias y RER.
3. Laminillas anulares.
Células foliculares
Las células foliculares son epiteliales planas que forman una sola capa llamada
granulosa, que envuelve al oocito y está separada de él por medio de una lámina basal
extremadamente delgada.
252
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Folículos primarios
Tienen un tamaño mayor, de entre 80 y 100 micras. Ya no son unilaminares, sino
multilaminares. Como consecuencia de su crecimiento van a entrar en la corteza. Están
compuestos por:
x Un oocito en el centro.
x Una lámina basal llamada zona pelúcida que separa el oocito de la granulosa
x La granulosa multilaminar.
x Una lámina basal llamada membrana limitante externa, que separa la granulosa
de la teca.
x Teca.
Oocito
El oocito sigue detenido en la misma fase, aunque aumenta de tamaño. En cuanto a sus
características microscópicas:
x Ya no se observa el cuerpo de Balbiani.
x Aumenta la cantidad de orgánulos, sobre todo de RER y ribosomas.
x Hallamos un Golgi compuesto por múltiples dictiosomas pequeño en vez de un
dictiosoma grande y único
x Aparecen los gránulos corticales, que son esenciales para la fecundación y se
liberan en ese momento.
x Inclusiones lipídicas y pigmento lipocromo en el citoplasma.
x La membrana presenta cortas microvellosidades que se dirigen al espacio
perivitelino.
Zona pelúcida
La zona pelúcida es una zona de MEC muy refringente, PAS positiva, que observamos
cuando el folículo es mayor de las 50-60 micras. Esta MEC es sintetizada por las células
de la granulosa y el oocito, y en ella podemos hallar las microvellosidades del oocito y
prolongaciones delgadas de las células foliculares.
253
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Células foliculares
Las células foliculares presentan receptores de FSH, hormona que estimula su
crecimiento y división. Por este motivo, el epitelio folicular se hace multilaminar y
aumenta la altura de las células que lo componen, las cuales se hacen cúbicas.
Este epitelio no está vascularizado, pero tampoco existen uniones herméticas entre las
células que impidan el paso de sustancias. En su lugar, hallamos uniones tipo GAP
formadas por conexina 43, así como nexos entre las células foliculares y el oocito,
formados por conexina 37, cuya mutación produce cambios en la maduración del
oocito, la meiosis y en la impronta.
Las células foliculares producen testosterona a partir de la androstenodiona sintetizada
por las células de la teca. Tras ello, modifican la testosterona para producir estrógenos,
en esencia estradiol.
Membrana limitante externa y teca
Es una lámina basal que separa las células foliculares de la teca, un conjuntivo
diferenciado alrededor de la granulosa. La teca tiene dos estratos:
1. Teca interna: formada por fibroblastos con función secretora. Se encarga de la
síntesis de colágeno, y también sintetiza la androstenodiona a partir de la cual
las células foliculares producirán testosterona, de ahí su amplia vascularización.
Además, a este nivel las células tienen recetores para LH. Dichas células tienen
un metabolismo claramente lipídico, lo cual es la razón de la presencia de
mitocondrias con crestas tubulares, REL y vesículas lipídicas.
2. Teca externa: es un tejido conjuntivo denso donde podemos hallar miocitos lisos
que se disponen de forma concéntrica alrededor de la teca interna y se continúan
con el estroma conjuntivo (la capacidad contráctil tendrá importancia en la
extrusión). También está vascularizada, aunque no tanto como la teca interna.
Resumen
En resumen, el desarrollo folicular depende de:
x Conexinas 37 y 43.
x C-kit y factor de células madre.
x BMP15 y GDF19, sintetizados por el oocito, que por tanto contribuye a la
estabilidad de la granulosa.
x MIS.
x Activinas.
x Inhibinas
Clínica
El síndrome del ovario poliquístico es una enfermedad importante en la mujer en la que
se produce una degradación del mecanismo de transmisión de señales entre el oocito y
la granulosa, por tanto, está implicada la conexina 37.
254
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Folículo secundario o antral
Su tamaño es todavía mayor, hablamos ya de entre 0’2 y 10 mm. Presenta los mismos
componentes que el folículo primario, aunque va a aparecer en él una nueva cavidad: el
antro.
Son folículos multilaminares, encontramos entre 6 y 12 capas de granulosa, pero entre
las células de la granulosa empiezan a abrirse espacios, que se llenan de líquido antral
(PAS+). Estos espacios se llaman cuerpos de Call-Exner, y van a ir convergiendo hasta
formar una cavidad en el centro del folículo: la cavidad antral, llena de líquido antral
(con ácido hialurónico, factores de crecimiento, esteroides y gonadotropinas).
Oocito
Es todavía un oocito primario detenido en diplotene de la profase I de la meiosis. Ha
aumentado del tamaño hasta las 125-150 micras. Sobre él actúa el factor inhibidor de la
maduración (OMI) segregado por células de la granulosa, que impide la maduración
meiótica espontánea y el excesivo aumento de tamaño.
El oocito ya no ocupa el centro del folículo, se halla en el antro rodeado de células
foliculares y zona pelúcida (cuyo conjunto se conoce como corona radiata). El conjunto
de oocito y corona radiata está suspendido en el antro por medio del cúmulus oophorus,
que lo ancla a la granulosa.
Células foliculares
Las células de la granulosa
siguen teniendo receptores
para FSH, y las que están
junto a la zona pelúcida
presentan microvellosidades
cuyo número aumenta con la
maduración. Aún existen
uniones GAP entre el oocito
y las células germinales.
Teca
La teca es igual pero más
grande debido al aumento de
tamaño del folículo.
Folículo de Graaf
Hemos dicho que el folículo
va a ir hundiéndose a medida
que progresa por los distintos estadíos, sin embargo llegados a este punto va a llegar a
tal tamaño que le es imposible profundizar más, y empezará a crecer protruyendo en la
superficie
Este folículo es básicamente aquel que está listo para la ovulación. Las células de la
corona radiada forman una capa simple debido a que las mitosis en la granulosa
255
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
disminuyen. Aun así, siguen presentando receptores de FSH y LH. La teca va a ser más
prominente.
Como mecanismo de preparación para la ovulación, disminuye el grado de unión entre
las células foliculares que constituyen el cúmulus oophorus o colina germinal. En el día
de la ovulación, el número 14 del ciclo menstrual, el conjunto formado por oocito y
corona radiada se libera, por lo que ha de estar disgregado del cúmulus.
Ovulación
Como ya se ha mencionado, consiste en la extrusión del óvulo, que tiene lugar en el día
14 del ciclo menstrual. Se produce una vez por ciclo, esto es, cada 28 días, y está
controlada por FSH y LH, hormonas para las cuales las células de la granulosa
presentan receptores.
Justo antes de la ovulación, la corona radiada y el oocito van a quedar libres en el
líquido antral, más acuoso que en días previos.
En la zona del ovario donde está el folículo de Graaf aparece el estigma pelúcido o
mácula pelúcida, que posee una tonalidad hialina, transparente o blanquecina, debida a
la interrupción del flujo en esa zona.
256
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
A la par, aparecen más enzimas proteolíticas en la teca externa y la túnica albugínea,
cuyo objeto es degradar estas capas, lo cual va acompañado por una desintegración del
epitelio germinativo y una disminución del estroma. Llega un momento en que el
folículo estalla y se libera el oocito con la corona radiata. Todos estos procesos son
paralelos a un pico de hormonas estrogénicas.
Simultáneamente, el extremo craneal del oviducto aproxima las fimbrias a la superficie
del ovario.
Si el ovocito no entra en el oviducto y es fecundado fuera de él, se produce una
implantación ectópica, que puede tener lugar en sitios variables:
1. Superficie del ovario
2. Saco de Douglas
3. Pared intestinal
Lo más frecuente es que esto no prospere, pero si lo hace puede comprometer
seriamente tanto la vida del feto como la de la madre.
En las horas previas a la ovulación, por acción
del MPF o factor promotor de la maduración, el
oocito supera la profase, con lo que culmina la
meiosis I, y se forman:
1. Oocito 2: se trata ya de una célula
haploide, que entra en meiosis II y se
detiene en metafase II por acción del
CSF o factor citostático, estado que solo
sobrepasará en caso de que se produzca
fecundación. En caso de que se finalice la
meiosis II se va a obtener un segundo
cuerpo polar, que degenerará como lo
hace el primero.
2. Primer cuerpo polar: degenera.
257
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Cuerpo lúteo
El folículo maduro no va a degenerar sin más, se va a transformar en una glándula
endocrina llamada cuerpo lúteo. Tras la liberación del oocito con la corona radiata, por
acción de los miocitos de la teca externa, se va a producir el colapso y plegamiento del
cuerpo lúteo o amarillo.
En el cuerpo lúteo desaparece la membrana limitante externa, que bloqueaba el paso de
los vasos sanguíneos, y se produce angiogénesis. La angiogénesis va a favorecer la
formación de un coágulo que será invadido por fibroblastos, fibras colágenas y vasos
(angiogénesis) y dará lugar a un centro fibroso.
En esta nueva glándula tanto las células foliculares como las células de la teca interna se
van a transformar a células luteínicas. Las células luteínicas son grandes y están
orientadas al metabolismo lipídico, de ahí su color amarillento y la presencia de
inclusiones lipídicas, REL y mitocondrias con crestas tubulares. Precisamente el
pigmento lipocromo luteína de estas células es el que da el color amarillo al cuerpo
lúteo. Además estas células secretan estrógenos y progesterona (estimulación
hormonal). Distinguimos dos tipos de células luteínicas en función de su origen:
1. Células luteínicas de la teca interna: Tienen un
tamaño menor, de unas 15 micras. Como es
lógico, son las más periféricas, y también las
que se hallan en menor número. Se tiñen
intensamente, y las localizamos junto a los
tabiques de la teca externa. Se encargan de
secretar progesterona y androstendiona en
respuesta a estímulo de LH.
2. Células luteínicas foliculares: Tienen el doble
de tamaño, y se sitúan en la parte central de la
célula. Secretan progesterona y estrógenos en
respuesta al estímulo de FSH y LH. Sin
embargo, carecen de las enzimas necesarias
para la síntesis de estradiol, por lo que lo
forman a partir de la androstenodiona
producida por las células luteínicas de la teca
interna.
La progesterona y los andrógenos segregados por el cuerpo lúteo estimulan el
crecimiento y la actividad secretora endometrial, preparando el endometrio para una
eventual implantación del cigoto en caso de que se produjese la fecundación. El
desarrollo máximo del endometrio se alcanza en el día 20 del ciclo menstrual.
258
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Cuerpo lúteo de la fecundación
Si no ha habido fecundación el cuerpo lúteo se conoce como cuerpo lúteo de la
menstruación. La luteólisis determina la apoptosis, a la cual contribuyen la hipoxia y la
invasión de linfocitos, que segregan interferón gamma y atraen a macrófagos que
segregan TNF-alfa, provocando la degeneración de las células luteínicas. Estas células
van a disminuir de tamaño, y disminuirá también su contenido lipídico, a la par que cesa
la síntesis de estrógenos alrededor del día 26.
259
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
La degeneración no implica la desaparición
completa del cuerpo lúteo, sino la formación del
cuerpo blanco o cuerpo albicans, más pequeño.
El cuerpo blanco sigue siendo una estructura
activa, pero no hormonalmente: las células
luteínicas van a dejar de sintetizar hormonas
para sintetizar colágeno a partir del día 26. El
cuerpo albicans va hundiéndose y persiste años
al fondo de la corteza ovárica. Debido a la
síntesis de colágeno, el cuerpo albicans cada vez
es más fibroso y pobre en células, y acaba
tabicándose.
Cuerpo lúteo de la gestación
Si hay fecundación el cuerpo lúteo se conoce como cuerpo lúteo de la gestación.
Aumenta de tamaño y sigue sintetizando estrógenos y progesterona por estimulación de
GH. Tiene que continuar siendo activo porque el endometrio se tiene que mantener,
pero a partir de la 10ª semana comienza la intervención en este proceso de la placenta,
el hígado y la glándula suprarrenal, lo cual implica que a partir del 6º mes se produce la
luteólisis (sin menstruación, por favor, Mari Pili se desespera si le decís lo contrario).
Otras funciones del cuerpo lúteo
x Segrega relaxina en el primer trimestre, que contribuye a inhibir las
contracciones espontáneas del miometrio. Ello en un principio interesaría en
etapas más tardías del embarazo, no se sabe muy bien el porqué de esta
contradicción.
x Estimula la síntesis de colagenasa, que relaja los ligamentos de la sínfisis
pubiana, lo cual es clave para el ablandamiento del cérvix previo al parto.
Desarrollo embrionario de los folículos
Los folículos ováricos aparecen durante el periodo embrionario, son primordiales y su
desarrollo es independiente de GH. A partir de la 10ª semana entran en meiosis, con lo
que se empiezan a detectar progresivamente folículos primarios. Entre el 5º y el 7º mes
de VIU de la niña hay unos 7 millones de folículos, pero cuando nace solo tiene 1
millón.
Al llegar a la menarca han disminuido a los 400 000, que son estimulados por FSH, LH
y GH. En cada ciclo, solamente un pequeño grupo se va a convertir en folículos
secundarios (6 o 7), y de ellos solo uno va a ser un folículo de Graaf. Al folículo de
Graaf se llama folículo dominante, y le distingue la cantidad de receptores para FSH
que posee. Al final de la etapa de madurez se habrán ovulado unos 400 ovocitos, y
llegamos al final de la etapa reproductora cuando no quedan ovocitos en los ovarios.
260
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Atresia
En la mujer hay degeneración/atresia absoluta de los
folículos de todo tipo formados en el desarrollo
embrionario.
No es patológico y afecta a más del 99% de los folículos
formados en el embrión. Ocurre en el desarrollo
embrionario y después del nacimiento. La velocidad a la
que degeneran los folículos aumenta tras los 40 años.
Cuanto más pequeña es la estructura del folículo (folículos
primarios o primordiales) más fácil es que se produzca la
atresia y que esta no deje huella, pero puede producirse en
cualquier tipo de folículo.
Los folículos secundarios sí que dejarán una huella que será
un cuerpo fibroso (que deja un rastro) que no se debe
confundir con el cuerpo blanco, aunque se parezcan mucho.
Cuando un folículo primordial o primario sufren atresia. El
primero que sufre apoptosis es el oocito y después las células foliculares degeneran.
Este folículo que es pequeño se va a retraer y el estroma que lo rodea invade por
completo el espacio que ocupaba. En un folículo secundario, en cambio, los primeros
cambios están en la pared y es el oocito el que sufre una degradación secundaria tras la
de la teca y la granulosa.
Las células foliculares presentan un factor NAIP (proteína nerviosa inhibidora de
apoptosis) que inhibe la apoptosis de estas células. Se expresa en todos los folículos,
pero cuando deja de expresarse esta proteína, el folículo empezará a sufrir atresia.
Un folículo atrésico tiene un oocito con un núcleo excéntrico que se hace picnótico
(muy irregular con cromatina extremadamente densa) progresivamente. Hay una clara
alteración del citoplasma del oocito y las células foliculares ya que están en apoptosis.
La zona pelúcida se mantiene aparentemente intacta. Los vasos que estaban a nivel de la
teca invaden la granulosa y la colina germinal, vascularizando esa zona. Un signo más
tardío de atresia es la dispersión de las células foliculares y la llegada de macrófagos,
fibroblastos y neutrófilos a la granulosa. Más tarde el antro es invadido por tejido
conjuntivo.
La membrana limitante externa, que separa la granulosa de la teca, comienza a
engrosarse y a sufrir plegamientos, adquiriendo una tonalidad refringente y brillante y
pasando a llamarse membrana vítrea o esmerilada. Se constata mayor separación entre
granulosa y teca.
Por último, la teca se hipertrofia y las células epitelioides tienen un aspecto que
recuerda bastante a las luteínicas. Se encargan de sintetizar estrógenos (metabolismo
lipídico).
Los folículos atrésicos en estado avanzado recuerdan mucho al cuerpo blanco. En
algunos textos se denominan cuerpos lúteos atrésicos pero no es un cuerpo lúteo sino
261
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
que es un folículo que no ha ovulado y sufre atresia mientras que el cuerpo lúteo sí que
ha ovulado.
En el folículo atrésico el estroma acaba rodeándolo y los cordones de células de la teca
hipertrofiada se fragmentan, quedando como células dispersas en vez de como una capa
continua. Esto es lo que se denominaba glándula intersticial, es decir, que parece que las
células de la glándula intersticial proceden de la teca de estos folículos atrésicos.
Médula
El ovario presenta una médula que es tejido conjuntivo laxo, no hay ningún tipo de
folículos pero sí una gran vascularización con vasos tanto sanguíneos como linfáticos.
Hay además un plexo venoso pampiniforme y fibras nerviosas.
262
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
ÚTERO
Es un órgano impar hueco situado en la zona de la pelvis menor. Se describe en forma
de pera, aplanado en el plano dorso-ventral. Se caracteriza porque su pared es gruesa
por el desarrollo de la capa muscular.
Funciones
Recibe al blastocisto y permite su implantación. Después sufrirá cambios que permitan
que permanezcan las relaciones vasculares y el embrión se nutra y crezca. Una vez
maduro el feto, contribuye a su expulsión debido a las contracciones que realiza el
miometrio.
263
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Cuerpo, istmo y cérvix
Anatómicamente se distinguen los dos tercios superiores que son el cuerpo y el tercio
inferior o cérvix. Entre ambos encontramos una zona estrecha o istmo.
En el cuerpo es la zona de mayor tamaño y donde hay más probabilidad de que ocurra la
implantación/anidación. Dentro de él hay una zona por encima a los orificios en los que
desembocan las trompas de Falopio, el fondo. El orificio interno permite comunicar la
luz del cuerpo con el cérvix.
El istmo es una región muy corta y estrecha (1cm) que está en la zona superior del
cuello y que histológicamente es igual al cuerpo, pero con una mucosa más delgada. En
la mujer gestante no se distingue y pasa a ser una parte más en el cuerpo.
El cuello o cérvix es una parte en forma de tubo con los extremos más estrechos (forma
abombada). Es simétrico y tiene una pared bastante delgada. En las mujeres que ya han
tenido embarazos adquiere forma de tonel o barril.
El orificio externo comunica el cuello con la vagina. Ese orificio externo, que en
principio es redondo, tras un parto natural adquiere la forma de hendidura transversal.
La porción del cuello que está metido en la vagina se denomina porción vaginal.
El cuerpo está organizado en tres capas histológicas:
x Endometrio (mucosa)
x Miometrio (muscular)
x Perimetrio (serosa)
Endometrio o Mucosa
El endometrio muestra cambios histológicos a lo largo de la vida. En la infancia tienen
epitelio cúbico bajo con un estroma poco desarrollado (recuerda al mesénquima) con
glándulas poco formadas e inactivas. En la época postmenopáusica tiene epitelio cúbico
bajo aunque el estroma es más denso y compacto y es frecuente la presencia de quistes
de las glándulas además de algunas glándulas activas.
En la época fértil se puede dividir en dos estratos funcionales:
x La capa más superficial o funcional es la capa que va a experimentar cambios
cíclicos y que en cada uno de estos cambios se desprende y tiene que volver a
regenerarse. Es la sensible a hormonas.
x La capa basal o profunda contacta con el miometrio y no experimenta cambios,
sino que siempre se conserva. A partir de ella se regenera en cada ciclo la capa
superficial.
Durante la época fértil el epitelio es prismático simple recubierto por una cubierta
proteica protectora. En él tenemos células ciliadas y células secretoras (secreción
proteica).
Debajo encontramos una lámina propia (TC) que ocupa un grosor importante con una
constitución bastante laxa que recuerda mucho al mesénquima y que tiene cambios
cíclicos en su composición a lo largo del ciclo. Luego la matriz de la lámina varía a lo
264
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
largo del ciclo. También es importante la enorme cantidad de glándulas tubulares
simples que encontramos. En principio las glándulas están restringidas a esta capa.
Clínica: Adenomiosis
El desarrollo de las glándulas es tan abundante que invaden el miometrio (no se quedan
en el endometrio).
Capacitación
La secreción propia varía a lo largo del ciclo. Permite cambios en los espermatozoides
para que puedan madurar y circular (capacitación). También son esenciales para lograr
la implantación de un blastocisto. Las porciones profundas de las glándulas están en la
capa basal y son las que permiten la regeneración de las glándulas y el epitelio
superficial.
La capacitación es un proceso muy corto en nuestra especie (minutos). Los
espermatozoides sufren una serie de cambios en su membrana que les permiten ser
capaces de adaptarse al canal reproductor femenino y fecundar un ovocito. Entre estos
cambios está la pérdida de la cubierta de glucoproteínas de la membrana plasmática del
acrosoma.
También hay cambios en la disposición de determinadas moléculas de la membrana, lo
que permite la activación de ciertos receptores para ligandos de la zona pelúcida.
Además, los espermatozoides se hacen hiperactivos y nadan mucho más rápido
(movilidad mayor).
Vascularización
La lámina propia también está muy vascularizada. Los vasos entran por la zona inferior,
en el perimetrio (arterias uterinas) y alcanzan el miometrio (arterias arqueadas)
atravesándolo hasta la zona más interna junto al endometrio, donde forman
ramificaciones denominadas arterias basales que irrigan el estrato basal del endometrio
y de las cuales nacen otras que ascienden por la zona superficial, con forma espiralada,
hasta alcanzar la capa funcional que se denominan arterias espirales. Las arterias
espirales o helicinas son arteriolas realmente y se caracterizan porque en un momento
dado del ciclo sufren contracciones y dilataciones de forma rítmica. Después
encontramos un plexo capilar superficial que sale de las arterias espilares (que tienen
una trayectoria tortuosa). La sangre venosa se recoge en el plexo venoso debajo del
epitelio.
Ciclo del Endometrio
Es un ciclo continuo que dura 28 días. Se suele dividir en tres períodos:
265
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Fase Proliferativa (o reparadora)
La fase proliferativa ocurre a partir del día 4 y dura unos 10 días hasta el momento de la
ovulación (hasta el día 14). En este periodo el endometrio mide como máximo 5mm
(delgado) y sólo tiene capa basal, la cual es capaz de proliferar, respondiendo a los
estrógenos del ovario y se regenera la capa funcional que se encuentra al final.
266
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Se caracteriza por un gran número de mitosis a nivel del estroma y en los extremos
desgarrados de las glándulas. Esta fase recibe su nombre por las abundantes mitosis que
tienen lugar. Las células epiteliales en proliferación regeneran las glándulas y el epitelio
de revestimiento, consiguiendo un estrato funcional. Se consigue aumentar las glándulas
en número y longitud de forma que acaban siendo rectas y estrechas.
También prolifera la lámina propia, aumentando la sustancia fundamental y el colágeno
por las células estromales haciendo el estroma metacromático. Durante los diez días
también se recuperan los vasos superficiales que se han perdido (a partir de las arterias
del estrato basal) los cuales son bastante rectos. El tercio superior aún no mostrará
vasos.
Las arterias espirales se regeneran pero tendrán una trayectoria no muy tortuosa (son
bastante rectilíneas) y no alcanzan toda la longitud del estrato. Las células epiteliales de
las glándulas empiezan a acumular cantidades importantes de glucógeno en su
citoplasma basal.
Las células epiteliales expresan en esta etapa la Bcl2 (proteína antiapoptítica).
Fase Secretora o Progestacional
Va desde el día 16 al 27. El endometrio sigue creciendo, pero no a un ritmo tan
marcado, hasta 6-7mm, dándose un notable aumento de grosor. Esta fase depende de la
progesterona (fundamentalmente) y de los estrógenos que son segregados por el cuerpo
lúteo. Además, sigue habiendo mitosis pero no tantas, a menor ritmo y las arterias
siguen desarrollándose (de manera que se hacen espiraladas y alcanzan todo el grosor
del endometrio). Así todo el estrato funcional del endometrio está irrigado (se ha
reconstruido el plexo superficial). Las arterias espirales tienen una morfología
contorneada y espiralada.
Por otro lado, las glándulas se hacen más tortuosas (aserradas) y más grandes. En esta
fase las células glandulares comienzan a acumular en su citoplasma basal glucógeno
formando una estructura denominada vacuola retronuclear. Según avanzan los días y sus
respectivas noches, la vacuola se desplaza hacia el citoplasma apical mientras que el
núcleo migra hacia el polo basal.
En esta etapa es típico observar imágenes típicas de apocrinia. En las luces de las
glándulas se acumula una secreción eosinófila. Tienen una luz bastante amplia (dejan de
ser tan estrechas para dilatarse) y su secreción es un moco viscoso acidófilo. Aumenta la
cantidad de secreción. Luego las glándulas crecen en tamaño y complejidad.
Por último el estroma también experimenta cambio: aumenta su grosor. Se constata una
proliferación y se edematiza, lo cual es un paso previo para dejarlo listo para la
decidualización que se produce si se da la implantación. Si durante esta fase no ocurre
una fecundación y una implantación, al final de la fase secretora el endometrio pasa a la
fase isquémica o premenstrual.
Fase isquémica
En caso de que no haya implantación se da paso a la fase premenstrual o isquémica (1
día) en la que se da involución y retracción del endometrio. En el estroma se produce
267
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
una infiltración leucocitaria (gran cantidad de leucocitos), la actividad glandular
disminuye, la secreción se hace más espesa y las arterias espiraladas sufren periodos de
contracción y dilatación intermitentes que duran horas y suponen dejar sin riego
(isquemia e hipoxia) al tejido del estrato superficial, lo que supone que este se muera, el
estroma pierda grosor, sea menos edematoso y las glándulas dejen de segregar.
Se piensa que las contracciones sostenidas están estimulados por la prostaglandina 2alfa
(que se cree que se secreta por la caída de los niveles de progesterona, como
consecuencia de la retracción del cuerpo lúteo).
Aumenta el número de leucocitos y fibras reticulares (en el estroma) y el estroma se
adelgaza y se vuelve menos edematoso.
Fase menstrual
Esta fase isquémica, como hemos dicho antes, dura un día y da paso a la fase menstrual
que se numera en los días 0 a 4. Las arterias finalmente acaban rompiendo su pared
habiendo una hemorragia al estroma y, finalmente, a la luz (porque además de las
paredes de las arterias se rompe el estrato superficial) que supone el sangrado uterino
con eliminación del endometrio que estaba preparado para la implantación. Esta
hemorragia supone perder sangre arterial y venosa (sobretodo) alteradas, así como todas
las células epiteliales y conjuntivas, glándulas y secreción glandular.
Durante este periodo la coagulación esta inhibida parcialmente y la pared uterina tiene
la mucosa desgarrada ya que se ha desprendido todo el estrato superficial o funcional y
ha quedado únicamente la capa basal sin alterarse, conservando su funcionalidad, sus
vasos y su irrigación en perfecto estado. La pared uterina se encuentra en carne viva y
todo lo que se elimina se expulsa por la vagina.
Cuando hay una implantación, el blastocisto se forma y alcanza la cavidad uterina,
donde se implanta. No hay fase isquémica ni menstrual y las células del corion de la
placenta (en concreto las del trofoblasto) en desarrollo comienzan a secretar hCG y
otras hormonas que mantienen al cuerpo lúteo y lo estimulan para que continue
sintetizando estrógenos y progesterona. Así, se impide la involución endometrial, este
continua desarrollándose durante las primeras semanas de gestación a la vez que se
transforma, pues se produce la decidualización (transformación que se produce antes y
después de la llegada del blastocisto) definitiva del endometrio, que supone que este
aumente, se edematice más y adquiera capacidad secretora.
Las células deciduales del estroma son fibroblastos en las que se producen cambios
metabólicos y morfológicos. Son de mayor tamaño, pálidas, cargadas de glucógeno y
lípidos y responden a progesterona y a los estrógenos.
268
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Las células deciduales tienen diversas funciones:
x Aportan nutrientes para embrión en desarrollo
x Modulan la invasión de células trofoblásticas
x Impiden rechazo inmunológico de tejidos embrionarios y fetales
x Segregan prostaglandinas, relaxina y prolactina decidual
En los ciclos anaovulatorios no hay cuerpo lúteo y, por tanto, tampoco fase secretora.
La fase proliferativa se va a alargar hasta la eliminación.
Clínica
El carcinoma endometrial es el tumor maligno más frecuente del aparato genital
femenino.
La endometriosis afecta al 12% de la población femenina en edad reproductora, y hasta
al 30% de parejas con infertilidad. Implantación de agregados de endometrio fuera del
útero, el tejido endometrial implantado prolifera, secreta y sangra igual que el
endometrio. La hemorragia atrapada puede causar quistes, cicatrices y adeherencias. Se
producen periodos de dolor (dismenorrea), pudiendo causar sangrado excesivo
(menorragia) o hemorragia entre ciclos (menometrorragia). Prueba hematológica de
detección de proteína CA-125 sólo es eficaz en endometriosis avanzadas.
269
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Miometrio
Capa mayor de unos 2 cm, con una capa de miocitos de 20 micras. Está rodeado de un
conjuntivo rico en fibras elásticas. Hay tres capas: la superficial y la profunda con fibras
longitudinales u oblicuas, y la media con fibras circulares.
Tiene abundantes vasos sanguíneos y linfáticos en la capa media, por lo que se
denomina capa vascular. Las arterias de paredes gruesas se contraen tras el parto para
reducir las hemorragias posparto.
La orientación muscular no es distintiva, durante la contracción uterina las tres capas del
miometrio actúan en conjunto como un sinticio funcional.
En la gestación los miocitos muestras hipertrofia e hiperplasia (miocitos de 600
micras por hipertrofia y aumento del número de miocitos por hiperplasia). Además,
también hay aumento del tejido conjuntivo. Se acumula agua en el conjuntico y las
capas se adelgazan al irse estirando según crece el feto. Responde a la relaxina,
procedente del ovario y la placenta, y se inhiben las contracciones.
En el parto el miometro responde a oxitocina (neurohipófisis), que estimula las
contracciones. Tras el parto los miocitos se acortan, aunque muchos miocitos persisten
y el útero es mayor.
Clínica
Los miomas son tumores benignos en miometrio. Tiene forma redondeada y su
crecimiento es dependiente de hormonas, con una sintomatología muy variables. Puede
haber alteraciones cromosómicas en las células miomatosas.
Perimetrio
Es un mesotelio (peritoneo) con conjuntivo submesotelial que recubre el cuerpo.
Istmo
Mucosa similar a la del cuerpo, pero más fina
con epitelio más bajo, menos glándulas y los cambios
cíclicos menos definidos.
Cérvix
Zona cilíndrica con una luz con abundantes
pliegues transversales. Se comunica con el cuerpo
mediante el orificio interno y con la vagina por el
externo.
El canal cervical está lleno de moco en la época
de la madurez reproductiva.
270
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Endocérvix
Su mucosa se denomina endocérvix, con menor grosor
que el endometrio y superficie más irregular (plicae
palmatae). Hay invaginaciones profundas, las criptas
secretoras, con glándulas tubuloalveolares ramificadas
mucosas. Sufre cambios cíclicos muy leves,
principalmente en las glándulas para permitir el
transporte de los espermatozoides. Carece de arterias
helicinas y generalmente no sangra.
El epitelio es prismático simple, con células
secretoras y ciliadas. Las células ciliadas baten hacia la
vagina para eliminar el moco. Las células secretoras can
a sufrir cambios cíclicos:
x Fase proliferativa: Prismáticas bajas, con núcleos basales se convierten debido a
la secreción de estrógenos en prismáticas altas con el citoplasma lleno de
mucígeno.
x Fase secretora: Recupera la forma inicial de la fase proliferativa. Producen
lisozimas, proteínas antibacterianas y moco, que sirve como lubricante vaginal y
tapón o barrera ante patógenos.
x Ovulación: Moco acuoso y claro, muy abundante y con pH alcalino, permitiendo
la entrada de espermatozoides. En este momento es extensible y hay Na, K y Cl.
Se produce cristalización en hojas de helecho.
x Fase secretora: Secreción más viscosa y menos abundante, con un pH ácido, de
forma que se sella el útero para la preparación de cara a una posible
implantación y se impide el paso de espermatozoides.
x Embarazo: Se forma un tapón que cierra el cuello. Se elimina al iniciarse el
trabajo de parto (sangrado señal).
La secreción de las glándulas que hay en las
invaginaciones puede quedar retenida y producirse
quistes de Naboth.
Estroma
Debajo del epitelio hay un estroma conjuntivo denso
con miocitos lisos y abundante vascularización
(aunque no como en el endometrio).
Muscular
La muscular es más fina, rica en conjuntivo denso
con fibras colágenas y elásticas, por lo que es dura
(especialmente en la gestación). Al final de la
gestación se producen cambios en la MEC,
271
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
ablandándola. Esto hace que sea posible la dilatación del cuello y perita el paso de la
cabeza del feto.
Serosa
Solo en la parte posterior del cérvix. .
Porción vaginal
La parte del cuello uterino que se
proyecta dentro de la vagina se denomina
porción vaginal, hocico de tenca, ectocérvix o
exocérvix.
La mucosa es lisa. El epitelio es plano
estratificado no queratinizado, en continuidad
con el epitelio vaginal. Alto contenido en
glucógeno. El cambio de la porción anterior a
esta es brusco.
Este cambio se denomina zona de
transición o unión escamosocolumnar del cérvix, que en las mujeres en edad fértil está
situada justo por fuera del orificio externo. Aquí hay un aumento de glándulas, que es
frecuente que queden cubiertas por epitelio (quistes de Naboth).
Esta localización es dependiente de hormonas maternas. Tras la menarca, las hormonas
provocan una extensión del endocérvix (erosión cervical o ectropión). Se forma una
mancha roja de epitelio rojizo, la eritroplasia. Con el embarazo, el ectropión aumenta.
En esta zona de transformación son frecuentes los focos de displasia, puntos de
desorganización celular, que se desprenden prematuramente. Pueden llevar a un
carcinoma in situ (neoplasia intraepitelial cervical, CIN). Si el cuadro progresa se
convierte en un carcinoma infiltrante. Ambos cuadros patológicos son detectados con
una citología cervical.
La citología cérvico-vaginal también permite detectar si hay infección con VPH.
Distintas cepas del virus se han relacionado con la mayor parte de los cánceres de
cérvix. La detección de cepas de alto riesgo de ADN antes de que ocurra CIN. Se
detectan células planas con núcleos grandes y pálidos y un halo externo.
272
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
VAGINA
Órgano copulador femenino en la parte
inferior del canal del parto. Se extiende desde el
vestíbulo al cuello uterino, de unos 8-9 cm. Entrada
ocluida por el himen. Es un tubo fibromuscular, en
reposo colapsado anteroposteriormente. Forma un
ángulo aproximadamente recto con el útero.
Mucosa
Recubierta de moco, con abundantes arrugas transversales (pliegues). Tras el
climaterio se adelgaza y atrofia. El epitelio depende de estrógenos.
La vagina carece de glándulas, por lo que el moco procede de las glándulas
cervicales y de Bartholin. También puede haber trasudación de líquidos desde capilares
de la lámina propia durante la excitación sexual.
El epitelio es plano estratificado no queratinizado, es empujado por papilas de
tejido conjuntivo de la lámina propia subyacente. Se distinguen tres zonas:
x Profunda: Células cúbicas-cilíndricas, con núcleo redondo y citoplasma basófilo.
x Media: Células aplanadas, con núcleos redondeos retraídas, el citroplasma
menos basófilo y gránulos de glucógeno.
x Superficial: Células muy aplanadas, con citoplasma eosinófilo núcleos
picnóticos, glucógeno y gránulos de queratohialina.
Hay una exfoliación constante. Los Bacilos de Dölerdein (lactobacillus vaginalis) lo
transforman en ácido láctico e inducne que el pH sea ácido. Es un factor protector frente
a patógenos, pero no protege frente a enfermedades de transmisión sexual. Tras el
climaterio el pH aumenta, disminuyendo el ácido láctico, y las infecciones son más
frecuentes.
273
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
pH alcalino favorece frente a la proliferación de patógenos (estafilococos,
protozoos como Trichomonas vaginalis y hongos como Candida albicans). Aumenta
trasudado vaginal y se produce inflamación mucosa vaginal y piel vulvar
(vulvovaginitis). Diagnóstico sencillo con frotis vaginales. El tratamiento requiere
restaurar pH Bajo y uso de antibióticos y
sulfonamidas.
Este epitelio vaginal sufre cambios
cíclicos:
x Fase proliferativa: Aumenta el número
de capas hasta 40. Las células son
eosinófilas.
x Ovulación: Máximo grosor y
exfoliación. Gránulos de
queratohialina con núcleos picnóticos
en células superficiales.
x Fase secretora: Disminuye el grosor,
con menos contenido en glucógeno.
Las células son basófilas y hay
infocitos y bacterias.
En una citología:
x Fase proliferatica: =% células basófilas y eosinófilas
x Ovulación: >% células eosinófilas
x Fase secretora: >% células basófilas.
El método Papanicolaou es una tinción a base de hematoxilina, naranja G y eosina azur
utilizada en Frotis vaginales. Tiene valor diagnóstico sobre mucosa vagina y cervical.
Permite detectar alteraciones patológicas de la mucosa, respuesta de la mucosa a
cambios hormonales durante el ciclo menstrual y situación microbiana vaginal. Pap
cervicovaginales se utilizan también para el diagnóstico precoz del cáncer de cuello de
útero y del carcinoma endometrial. Células cancerosas se detectan fácilmente,
pudiéndose determinar su lugar de origen.
En la lámina propia se distinguen dos zonas:
x Superior: Conjuntivo laxo con linfocitos y leucocitos, que aumentan en la
menstruación. Se observan nódulos linfoides.
x Inferior: Conjuntivo denso, con fibras colágenas y elásticas. Hay venas de
paredes finas, que es tejido eréctil durante la excitación sexual. Escasas
terminaciones nerviosas.
Muscular
Capa interna circular y externa longitudinal, más gruesa, aunque ambos están
entremezclados y son difíciles de distinguir. En la zona inferior se encuentra el músculo
bulboesponjiso, que actúa a modo de esfínter y tiene fibras musculares estriadas.
274
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Adventicia
Estrato interno con conjuntico denso con fibras elásticas, lo que da elasticidad y
resistencia, y estrato externo con conjuntivo laxo, por donde entran y salen vasos
sanguíneos y linfáticos.
OVIDUCTO
El oviducto o trompa de Falopio es un tubo de naturaleza muscular par que se extiende
unos 12 cm y que presenta los dos extremos abiertos. Un extremo proximal abierto en la
cavidad peritoneal y un extremo distal abierto en la cavidad uterina.
Este oviducto proporciona el entorno adecuado para recibir al óvulo una vez producida
la ovulación y también para que se pueda producir la fecundación.
Distribución anatómica
Anatómicamente, el oviducto se puede dividir en cuatro partes:
x El infundíbulo o pabellón
x La ampolla
x El istmo
x El segmento intramural, el cual transcurre ya dentro de
la propia pared uterina.
En el esquema vemos tres secciones transversales del oviducto.
La primera y más grande corresponde a la zona de la ampolla.
La segunda, la intermedia corresponde a la zona del istmo y la última corresponde a la
zona del segmento intramural.
Conforme nos vamos acercando al útero el diámetro va decreciendo, y con ello la
proporción relativa de los componentes histológicos varía.
En la parte final, la pared es fundamentalmente muscular con una luz muy pequeña;
mientras que, en la zona más proximal, es decir, en la ampolla, tenemos un tamaño
mucho más grande, un diámetro mucho más amplio, pero sin embargo la luz es
tremendamente tortuosa y laberíntica.
Infundíbulo o pabellón
Tiene el extremo abierto hacia la cavidad peritoneal quedando cerca del ovario. Este
extremo no tiene unos bordes lisos, sino que se proyectan cortas prolongaciones que
reciben el nombre de fimbrias.
Sabemos que, en las horas previas a la ovulación, los vasos que hay en estas fimbrias
aumentan la perfusión de sangre y esto hace que crezcan de tamaño. Esta mayor
turgencia permite que las fimbrias se aproximen mucho más al ovario y esto aumenta
las probabilidades de que el óvulo que se va a liberar quede dentro del infundíbulo y no
en la cavidad peritoneal.
275
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Ampolla
Es una zona intermedia, con un gran diámetro y una luz amplia que va a ser muy
tortuosa.
Las paredes de esta zona son delgadas y en esta ampolla se da un transporte lento del
óvulo que ha entrado. Esto lleva a que aquí sea mayor la probabilidad de que se
produzca la fecundación.
Recordemos que, dentro de los posibles embarazos ectópicos, uno de los lugares donde
se puede implantar erróneamente el óvulo fecundado es la pared de la ampolla. Cuando
este embarazo ectópico ocurre, no va a poder progresar porque directamente la pared del
oviducto se rompe. Esto puede provocar una hemorragia interna y muy intensa en la
mujer.
Istmo
Es un segmento intermedio de luz más estrecha con una pared más gruesa. Sabemos que
en estas paredes se van a producir contracciones rítmicas que se dirigen hacia el útero.
Segmento Intramural
Esta dentro de la propia pared uterina y permite comunicar la luz del oviducto con la luz
del útero.
La luz de la ampolla se caracteriza por ser generalmente estrecha y laberíntica. Esto se
debe a la gran cantidad de pliegues de la mucosa. Estos pliegues son mucho más
abundantes, acusados y ramificados a nivel de la ampolla. Este número de pliegues y su
complejidad va a ir disminuyendo en el istmo para acabar siendo mínimos en el
segmento intramural.
Estructura histológica
Histológicamente el oviducto se compone de tres capas:
mucosa, muscular y adventicia. Como veremos, ocurren
cambios cíclicos.
En el esquema de la derecha nos representan fimbrias del
oviducto, y en la zona superior nos resaltan el epitelio
donde se ven tres tipos de células: BC (Células Basales)
que son pequeñas; y dos células de tipo prismático unas
con cilios y otras sin ellos.
Mucosa
El epitelio es prismático simple, teniendo una altura máxima a nivel de la ampolla. Esta
altura va decreciendo a medida que nos aproximamos hacia el útero. Como se ha
mencionado anteriormente tenemos tres tipos de células:
x Células Basales o células Madre
x Células Ciliadas
Presentan como función básica ayudar al transporte del óvulo desde el infundíbulo o
desde la ampolla, hasta el útero. Sabemos que el número de cilios, su altura y la
276
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
cantidad de células ciliadas varía a lo largo del ciclo femenino; siendo máximo en la
ovulación.
Posteriormente el número de células ciliadas, de cilios y su altura va disminuyendo. Al
final del ciclo femenino o cuando se termina la gestación se reanuda el crecimiento de
células ciliadas y cilios. Los cambios cíclicos mencionados son fundamentales a nivel
de las fimbrias y de la ampolla y tienen un control claramente endocrino.
Fundamentalmente dos hormonas: progesterona y estradiol.
Esto se pone claramente de manifiesto en mujeres a las que se les practica una
ovariectomía. Como consecuencia de ella, el epitelio del oviducto se atrofia y las células
ciliadas desaparecen por completo. Si a estas mujeres se les suministra estradiol, el
oviducto muestra una recuperación de su epitelio y de nuevo las células ciliadas
aparecen.
x Células Secretoras
Son células que carecen de cilios y sí tienen cortas microvellosidades en su membrana
plasmática apical. Estas células también pierden altura tras la ovulación.
Estas células forman una secreción liquida, muy acuosa y rica en iones potasio y
cloruro. Además, contiene proteínas séricas. Estos elementos son importantes para las
primeras horas tras la ovulación y en el caso de que ocurra la fecundación, es importante
para los primeros días de desarrollo embrionario.
La lámina propia es el tejido conjuntivo que encontramos bajo este epitelio. En el caso
del oviducto muestra una gran cantidad de fibras reticulares. También son muy
abundantes las células conjuntivas; no sólo fibroblastos, sino también células del
sistema inmunológico.
En este tejido encontramos una gran cantidad de vasos. Luego se trata de una lámina
propia muy vascularizada y, en el caso de la mujer gestante, la lámina propia se
hipertrofia.
Muscular
Es una capa muy gruesa en la que aparentemente encontramos dos estratos: Circular
Interno (CI) y Longitudinal Externo (LE). A estos dos estratos e le suma un tercero en
las zonas más próximas al útero.
En realidad, no hay tales estratos en sí, sino que se trata de una disposición helicoidal o
espiralada.
Esta capa se encarga de crear los movimientos peristálticos, los cuales van a ser
máximos en la ovulación y son responsables del avance del embrión en el caso de que
se haya producido la fecundación. Estos movimientos están controlados de manera
hormonal.
Serosa
Es la adventicia que corresponde al peritoneo.
277
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Cambios cíclicos
Para terminar, recopilemos los cambios cíclicos que hemos ido mencionando. Sabemos
que durante el periodo de la ovulación se producen niveles máximos de peristaltismo
que ocurren desde el infundíbulo hacia el útero, de batido ciliar que se bate en la misma
dirección y finalmente del desarrollo vascular. Este desarrollo vascular es
fundamentalmente patente en la ampolla y en las fimbrias.
278
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
PLACENTA
IMPLANTACIÓN
La implantación del cigoto se produce hacia el día 6 en la parte superior de la
pared uterina posterior, e implica básicamente dos procesos:
1. Adhesión inicial y posterior penetración del blastocisto en el endometrio
2. Transformación del endometrio grávido en decidua o decidualización
En un principio, la adhesión que establece el blastocisto con la parte superior de
la pared posterior uterina es altamente inestable. La membrana plasmática de las células
del epitelio uterino muestra el factor de crecimiento ligado a la heparina o HB-EGF,
capaz de unirse a los receptores para HB-EGF presentes en las células trofoblásticas (y
también al heparán sulfato). Además, contribuyen a la unión las prolongaciones de
ambas células: las células epiteliales uterinas muestran pinópodos, que interaccionan
con las microvellosidades de las células trofoblásticas.
Una vez establecida esta interacción, el endometrio se transforma en una zona
decidual primaria: las células estromales comienzan a proliferar y se transforman en
células epiteloides. Se encargan de secretar:
x Moléculas propias de la matriz celular normal: fibronectina, laminina, colágenos
de tipo I, II, III y IV, entactina…
x Moléculas exclusivas de la matriz decidual.
Además, se ha de destacar que estas células expresan proteínas morfogenéticas
óseas BMP-1 y BMP-2, así como Wnt-4 y proteína Hedgehog.
Tras esta fase de proliferación epitelial, comienza una etapa de erosión:
desaparecen las uniones intercelulares entre las células epiteloides del estroma decidual
y esto posibilita la apoptosis de muchas de las células, y con ello la reducción de la
población. Además, en el punto en el que el trofoblasto ha entrado en contacto con el
epitelio va a aumentar la permeabilidad vascular.
La decidua va a continuar su proceso de diferenciación, de manera que se va a
constituir la llamada zona decidual secundaria. Este proceso de decidualización
secundaria viene determinado por la prostaglandina 2, que activa un proceso en el que
participan metaloproteinasas, que posibilitan la remodelación (inhibidores de MMP y de
la remodelación y activadores del plasminógeno?).
Entretanto, el proceso de implantación se va a estabilizar y el blastocisto va a
comenzar a penetrar en la decidua. En el blastocisto, las células del trofoblasto ya se han
diferenciado formando dos capas diferentes:
x Citotrofoblasto (CT): Es la capa más interna, cuyas células siguen dividiéndose
activamente.
x Sincitiotrofoblasto (SCT): Es la capa que queda externa, está constituida por un
conjunto de núcleos que forman un sincitio funcional. Esta capa tiene carácter
invasor, puesto es la que va a permitir el avance del blastocisto hacia el interior
de la decidua.
279
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
En el proceso de penetración distinguimos tres grandes fases:
1. Invasión por fusión: Las células del
SCT se fusionan con las células
deciduales, formándose una masa
que contiene tanto núcleos maternos
como núcleos trofoblásticos.
2. Invasión intersticial: Conlleva la
acción de enzimas proteolíticas
segregadas por el sincitiotrofoblasto
3. Invasión endovascular: Se produce
la erosión de las ramas de las
arterias espiriladas del útero, que
contribuyen a la formación de la red
lacunar.
En el SCT, a medida que este
penetra, se van a comenzar a formar lagunas de sangre materna, que irán aumentando a
medida que el SCT progresa y va erosionando los capilares maternos dilatados. Este
aumento de tamaño va a conducir finalmente a la fusión de las lagunas, con lo cual se
constituye una red lacunar hacia la cual fluye la sangre materna: se trata de la antefase
de la circulación úteroplacentaria, que llamamos circulación úteroplacentaria primitiva.
En el proceso de penetración se van a constituir vellosidades de tres tipos:
x Entre los días 11 y 13, el mesodermo induce la proliferación del citotrofoblasto
(CT), que empieza a desplazar al sincitiotrofoblasto (SCT), formando cordones
en su interior. Se forman así las vellosidades primarias, que están constituidas
por una masa de CT rodeada de la capa del SCT.
x Unos 5 días más tarde, el mesodermo que ha inducido la proliferación invade los
cordones de citotrofoblasto, de manera que se constituyen las vellosidades
secundarias, formadas de interior a exterior por mesodermo, CT y SCT.
x Hacia el día 20 el mesodermo va a comenzar a estar vascularizado debido a la
diferenciación en su seno de capilares y células sanguíneas. Una vez que ha
comenzado este proceso la vellosidad es vascular y pasa a llamarse vellosidad
terciaria.
280
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Hemos hablado del carácter invasor del
sincitio y de la formación de lagunas, pero las
células del CT también proliferan y penetran
hasta el endometrio. La parte externa del CT,
denominada corteza citotrofoblástica, va a
actuar como límite del CT, rodeando al
blastocisto y fijándolo a la decidua por medio
de las vellosidades de anclaje, que poseen unas
ramificaciones laterales denominadas
vellosidades terminales o libres porque quedan,
efectivamente, libres en el espacio lacunar.
Recordemos que ya se ha formado una primitiva red úteroplacentaria, pero el
corazón aún no funciona. Hacia el día 21, el corazón comienza a latir, por lo que entran
en juego dos circulaciones:
x Circulación embrionaria: Es una circulación cerrada en la que la sangre
embrionaria fluye hacia los capilares que habían invadido las vellosidades, a su
vez inmersas en la sangre materna de la red lacunar.
x Circulación materna: Es abierta. La sangre materna penetra a baja presión en la
red lacunar.
Por tanto, la sangre materna y la embrionaria entran en contacto a la altura de las
vellosidades, inmersas en la red lacunar, y es ahí donde se va a producir el intercambio
de productos.
Las arterias umbilicales van a llevar la sangre desoxigenada del embrión hacia
las vellosidades. Ahí, dicha sangre va a tomar el oxígeno y los nutrientes de la sangre
materna, que a su vez tomará los desechos y quedará desoxigenada. La sangre
embrionaria oxigenada retornará al embrión a través de la vena umbilical, y la sangre
281
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
materna desoxigenada retornará a la
circulación materna a través de las venas
uterinas (cuidado aquí, sucede como en el
caso de los pulmones: la arteria umbilical
lleva sangre desoxigenada).
DECIDUA
Como ya hemos dicho,
denominamos decidua al
endometrio del útero grávido, es
decir, de una mujer gestante. Su
función es protectora y de nutrición
del embrión (acumulan glucógeno
y lípidos). Está organizada en
varias capas:
x Basal: Es la única que no se
elimina tras la conclusión de la gestación, en el parto. Se sitúa inmediatamente
bajo la zona de implantación del embrión, fijada a la placa coriónica, de manera
que constituye el componente materno de la placenta.
x Capsular: Es el endometrio que recubre al corion y al blastocisto implantado,
que ya han penetrado y han quedado englobados en la decidua.
x Parietal: Es el endometrio que recubre el resto del útero, que no está en contacto
estrecho con el corion.
Vemos que los tejidos embrionarios están en estrecho contacto con los maternos,
con lo que cabría esperar una reacción inmunológica. Sin embargo, las propias células
deciduales se encargan de segregar prostaglandinas e inmunosupresores que inhiben la
activación de los NK, posible porque aunque las células del SCT no expresan MHCII sí
expresan MHC no clásicas (E, G...). Los linfocitos T tampoco se activan. Los leucocitos
infiltrados segregan IL2 e IL4 y TNF-beta, que contribuyen a evitar el rechazo materno
del embrión.
La capa coriónica que queda por debajo de la decidua va a sufrir distinta
evolución zonal:
x La decidua capsular está asociada con el corion, que está cubierto hasta la 8ª
semana por vellosidades en toda su extensión. En este momento, las vellosidades
comienzan a degenerar hasta desaparecer completamente en la 13ª semana,
constituyendo el corion liso o leve, asociado por tanto a la decidua capsular.
Finalmente la decidua capsular degenerará y el corion leve se acabará
fusionando con la decidua parietal a medida que el feto crezca, quedando la
cavidad uterina con una luz casi inexistente.
x La decidua basal está asociada con la placa coriónica, y en este punto las
vellosidades van a aumentar y ramificarse, constituyendo el corion frondoso o
componente fetal de la placenta (recordemos que el materno es la propia decidua
basal).
282
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
PLACENTA
Es un órgano en
forma de disco. Cuando
está madura, a partir del 5
mes, pesa 500g y tiene
20cm de diámetro. Ocupa
el 30% de la superficie
interna de la cavidad
uterina. El aumento de
tamaño se debe al aumento
del número de las
vellosidades y su mayor
complejidad debido a las
ramificaciones que sufren.
Anatómicamente observamos dos caras:
x Fetal: Lisa y brillante, debido a que está recubierta por el amnios. En el centro
de esta cara está el cordón umbilical.
x Materna: Mucho más opaca, dividida en pequeñas secciones de aspecto cóncavo.
Se encuentra delimitada por zonas que se hunden (surcos). Cada zona delimitada
recibe el nombre de lóbulo o placentomo.
Así podemos definir placentomo como zona de la placenta delimitada por dos
anillos tabicarios consecutivos. Estos tabiques son estructuras que parten de la decidua
basal pero no tabican del todo la placenta pues no llegan al lado opuesto (placa
coriónica).
Histológicamente hay tres zonas, que se irán describiendo:
283
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Placa coriónica: Está formada por corion + amnios
x Placa basal: Decidua basal (lugar en el que se mantienen las vellosidades
coriónicas) + la capa basal del endometrio. Se observa dividida en lóbulos o
placentomos.
x Vellosidades: Estructuras que van de un punto a otro, son de anclaje. Se
encuentran flotando en la sangre materna.
Placa Coriónica
El amnios es epitelio cúbico simple, es frecuente encontrar en él procesos de
metaplasia. Bajo este epitelio encontramos una pequeña zona de conjuntivo que carece
de vasos. Está en continuidad con el corion.
A continuación, viene el corion, el cual está formado por el mesodermo
extraembrionario, citotrofoblasto y sincitiotrofoblasto.
El primer conjuntivo que hay está asociado al amnios y no presenta vasos. Por el
contrario, el conjuntivo del corion sí tiene vasos y muy grandes (vasos umbilicales).
Los tabiques nacen de la parte materna, tienen distintas alturas, pero nunca
llegan a contactar con la placa coriónica.
Placa Basal
Formada por una capa de citotrofoblasto que hace de límite y a la cual se anclan
las vellosidades coriónicas. Por fuera de esa corteza/límite, está la decidua basal. En esta
decidua basal encontramos la matriz y sus células correspondientes. También podemos
hallar células X. No son de la madre, es decir, no son deciduales; sino que son células
procedentes del citotrofoblasto, aunque quedan aisladas dentro de la célula. Se han
independizado de él, perdiendo el contacto. Se caracterizan por ser muy grandes y
contener muchos núcleos.
En cada lóbulo o placentomo, hay múltiples vellosidades de anclaje. Si cogemos
el espacio en el que solo hay una vellosidad de anclaje, eso es el lobulillo o cotiledón.
Aparte de estos elementos, están las estructuras llamadas fibrinoides. Son zonas
eosinófilas, siendo además elementos de la matriz.
En algunas placentas la cantidad de estos fibrinoides es tan grande que llega a
sustituir vellosidades completas. No se sabe ni su causa ni su origen. Hay teorías, como
que pueden ser por envejecimiento del órgano, que pueden contener células muertas de
trofoblasto… Pero no se tiene nada seguro.
Hay dos grandes grupos:
x Tipo fibrina: Se constata que este fibrinoide está formado sobre todo por fibrina.
En este caso se trata de puntos en los que ha habido lesión en el sincitio. Luego
la sangre que está en contacto con él, empieza a coagular y se forma fibrina.
Aquí se quedan fijados anticuerpos maternos.
x Tipo matriz: Hay fibrinoides que no son compatibles con fibrina. Estos se
forman debido a un depósito que aparece en la matriz segregado por el
citotrofoblasto.
284
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
En función de sus localizaciones distinguimos:
x Fibrinoide de Langhans, sobre la placa coriónica.
x Fibrinoide de Nitabuch, en la placa basal.
x Fibrinoide de Rohr, en las vellosidades.
Vellosidad coriónica terciaria o madura
La unidad estructural y funcional de la placenta es la vellosidad coriónica
terciaria. Ya está vascularizada. Hay 15 m2 de vellosidades coriónicas. Hasta el 4º-5º
mes las vellosidades están formadas por (de dentro a fuera):
x Un eje de mesodermo con células mesenquimáticas las cuales se van a
diferenciar a fibroblastos y participan en la síntesis de diversos tipos de
colágenos (tipos I, III, V y VI). El eje de mesodermo también presenta un tipo
de macrófago, que es específico de esta zona, denominado célula de Hofbauer
(aparecen en las primeras fases del embarazo). Este tipo de célula es más
abundante en vellosidades primarias, pero persiste en todos los tipos de
vellosidades.
x Capilares.
x Lámina Basal que separa el mesodermo del citotrofoblasto.
x Citotrofoblasto: Es una capa de células epiteliales que se están dividiendo,
unidas entre sí por abundantes desmosomas. Estos desmosomas también unen el
citotrofoblasto con el sincitiotrofoblasto que es la siguiente capa. Se dividen y se
diferencian en células del sincitiotrofoblasto.
x Sincitiotrofoblasto está en contacto con la sangre materna. Las células son
postmitóticas.
Es frecuente identificar depósitos de fibrina en la superficie de la vellosidad en
áreas sin células del sincitiotrofoblasto y antes de la reepitelización.
Cambios
A partir del 4º-5º mes las vellosidades adelgazan y poseen menos componentes.
El primer componente que se pierde es el citotrofoblasto. Quedan unas pocas células,
pero no dan para formar un epitelio continuo.
El mesodermo aunque disminuye de tamaño, presenta un aumento del número y
tamaño de los vasos. Luego los capilares van a estar más cerca de la zona periférica de
la vellosidad.
El sincitiotrofoblasto es una masa polinucleada en la que sus núcleos se van
juntando formando pequeños agregados. Estos núcleos se desplazan hacia la sangre
materna y quedan como colgajos.
En la vellosidad el sincitiotrofoblasto está muy adelgazado porque apenas hay
núcleos. Estos quedan como brotes sincitiales. Muchos brotes se desprenden y se
incorporan a la circulación materna, acabando siendo degradados en los pulmones
maternos. Este último proceso es una estrategia que persigue acercar la sangre fetal a la
materna, pues hace que la distancia entre ambas sangres disminuya.
285
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Esta distancia es la barrera o membrana placentaria. La distancia presenta seis
elementos y a partir del 5º mes, la distancia de las sangres disminuye. Esto quiere decir
que la maduración de la placenta consigue reducir esta barrera placentaria, que hace que
el intercambio de productos sea más rápido y eficaz.
Funciones
x Actuar como órgano que nutre al embrión y al feto: Importante durante el
desarrollo y sobre todo al principio. Sintetiza glucosa, colesterol y ácidos grasos.
x Permite el transporte de múltiples moléculas: Las moléculas que traviesan la
membrana placentaria, lo hacen de manera bidireccional. El aumento de
vellosidades permite que una placenta madure, y que la superficie de
intercambio aumente de 15 m2 a 100 m2.
Mediante mecanismos como difusión facilitada, transporte activo, pinocitosis...
pero sólo pasan células sanguíneas cuando hay fallos en la barrera.
o Algunos elementos que se transportan tienen que ver con la nutrición.
Estos son agua, aminoácidos, glucosa, iones... Sustancias como
triglicéridos, proteínas o péptidos no atraviesan la membrana.
o Otros tienen que ver con la excreción. El feto genera catabolitos que solo
puede eliminar a través de la madre, como ácido úrico, urea o bilirrubina.
o Intercambio de gases, como oxígeno, monóxido de carbono o CO2. La
sangre fetal solo se puede oxigenar a través de la placenta.
Además, hay que añadir los gases anestésicos, que afectarían a la salud
del feto.
La reducción en la funcionalidad de la membrana puede disminuir el
oxígeno, generando hipoxia y daños cerebrales, incluso la muerte fetal.
o Las Igs maternas pasa a través de la membrana en grandes cantidades y
le confieren inmunidad pasiva al feto.
Cuando se presenta el caso de una madre embarazada Rh- y el
feto Rh+ la sangre fetal y la materna entran en contacto en el
parto, momento suficiente para que se generen anticuerpos en la
madre. Si esta vuelve a quedar embarazada y el hijo también es
Rh+, es peligroso pues puede generar hemolisis de los hematíes
fetales. La madre reconocerá ya el Rh+ como “antígeno”, ya que
había generado anticuerpos en el embarazo anterior.
o Las hormonas maternas: Atraviesan la placenta ejerciendo un efecto
sobre el embrión. Como los andrógenos, generando la masculinización
de los fetos femeninos en determinadas patologías.
o Fármacos y drogas también atraviesan la membrana placentaria. Los
medicamentos también en su mayor parte. El caso de la talidomida ha
sido el más sonado. Fármacos empleados para facilitar el parto pueden
dañar al feto, causar dificultades respiratorias.
o El alcohol también puede generar retraso mental. Las drogas pueden
provocar toxicomanía fetal.
o Microorganismos como virus pueden atravesar la membrana, que puede
dar lugar a malformaciones congénitas o incluso la muerte. Los daños
286
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
del virus dependen del momento concreto de la infección. Esto es, un
agente puede ser muy peligroso o no generar daños según el momento.
x Funciona como órgano endocrino:
o Gonadotropina Coriónica (hCG): Se sintetiza desde el día 6. Es clave
para que el cuerpo lúteo no sufra luteolisis, sino que se mantenga.
Estimula la secreción de tiroxina.
Sin embargo, el cuerpo lúteo va perdiendo protagonismo en favor de la
propia placenta. La placenta va a asumir funciones que sólo lleva a cabo
el cuerpo lúteo. Esto es el desplazamiento luteo-placentario y ocurre a
partir de la 7ª semana de gestación.
o Progesterona: Está sintetizada y segregada por el sincitiotrofoblasto y
mantiene la decidua durante toda la gestación. Lleva a cabo funciones
inhibitorias sobre el endometrio, inhibe las contracciones y es una
hormona fundamental para el desarrollo de la mama.
o Estrógenos: El sincitiotrofoblasto no puede sintetizarlos por sí mismo
pues le faltan enzimas. Así lo que se establece es una colaboración feto-
placentaria. La progesterona del sincitio llega a la corteza suprarrenal
fetal. Y ahí si podemos transformar la progesterona en andrógenos.
El DHEA sulfatado no alcanza de nuevo la placenta y ahora ya sí, el
sincitio toma este DHEA y con ello forma estradiol y estrona. Hay un
tercer estrógeno que es el estriol. Este se forma a partir del DHEA
cuando esta hidroxilado. Los estrógenos son claves para el
mantenimiento de la gestación, para el desarrollo de los conductos
mamarios y la estimulación del desarrollo de conductos y órganos
sexuales fetales.
o Lactógeno placentario: Sintetizado y segregado por el sincitio. Es clave a
partir de la semana 20 para estimular el crecimiento fetal y también para
la lactancia. Antes de la semana 20 no se sintetiza y en ese caso depende
de la GH materna.
El problema del lactógeno es que también actúa sobre la madre, haciendo
que el metabolismo materno se desvíe. Esto quiere decir que la madre
desvía sus hidratos de carbono para dárselos al feto. Con ello, la madre lo
que hace es consumir lípidos para a partir de ellos obtener glucosa.
Esto, si sobrepasa un cierto límite, puede causar un efecto diabetogénico:
durante el embarazo la mujer ha sido diabética sin serlo antes por lo
explicado anteriormente. En algunos casos después del embarazo siguen
siéndolo.
Así que no sólo es perjudicial para la madre; se ha demostrado que el
feto tiene problemas: tiene una macrosomía fetal. Este hecho se
correlaciona con que el bebé acabe siendo un adulto diabético y obeso.
o IGF I e IGF II: Son dos moléculas de carácter endocrino hormonal
sintetizadas por el citotrofoblasto. Tienen más bien un carácter paracrino
porque actúan sobre sí mismas estimulando la mitosis y el transporte del
y al sincitiotrofoblasto.
287
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
o EGF: Sintetizado en el citrotrofoblasto y a partir de la 6ª semana por el
sincitiotrofoblasto. Inicialmente incide sobre la tasa de proliferación y
posteriormente favorece el mantenimiento de funciones del trofoblasto.
o Relaxina: Sintetizada por células deciduales. Colabora reblandeciendo el
cérvix. Actúa sobre la capa muscular y los ligamentos medianos dando
su relajación en el parto.
o Lectina: Sintetizada por el sincitiotrofobalsto sobre todo al final de la
gestación. Actúa sobre el metabolismo materno haciendo que la madre
almacene más nutrientes cada vez para el feto. Luego aumenta la
disponibilidad de nutrientes para el feto.
Clínica
x Placenta previa: Se produce cuando la implantación del blastocito no ocurre en
el cuerpo, en las zonas más frecuentes como es el fondo, son en el cuerpo. El
blastocito se implanta en zonas más próximas al cérvix o cuello uterino. Esto
hace que cuando se desarrolle la placenta, en muchos casos acabe metiéndose en
el canal cervical. Se da de manera espontánea. Es indolora y muy peligrosa tanto
para la madre como para el feto.
x Desprendimiento de la placenta: La placenta se desprende antes de tiempo, pero
la implantación es normal, no es de riesgo. La separación espontánea es dolorosa
y aquí si hay factores asociados: consumo de drogas, traumatismo o hipertensión
materna o eclampsia.
x Atonía Uterina: Una vez que nace el niño se tiene que expulsar la placenta. Para
ello es necesario la contracción del miometrio. No sólo es necesario para
expulsar la placenta, sino también para que los vasos se contraigan y haya un
menor sangrado. Cuando estas contracciones no se producen, la placenta puede
desprenderse o no, pero no acaba de expulsarse.
x Placenta accreta, increta y percreta: Hacen referencia a placentas en las que el
desarrollo de las vellosidades coriónicas es masivo, no están sólo en la decidua
basal, sino que se extralimitan. En la increta se dan casi en el miometrio las
vellosidades, en la percreta están por todas partes casi llegando al perimetrio. A
más vellosidades, más difícil es una buena separación completa.
x Molas hidatiformes: Óvulos vacíos fecundados por espermatozoides. Cuando
esto ocurre, en la placenta el tejido es anormal, aparecen vellosidades muy
edematosas y grandes. La quinta parte de los embarazos con molas, dan igual a
coriocarcinomas.
288
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
OÍDO
Órgano sensorial complejo, que forma parte del sistema auditivo (audición) y del
sistema vestibular (equilibrio).
Se divide en tres porciones: el oído externo y el medio reciben y transmiten la
energía sonora, mientras que el oído interno la transforman en energía eléctrica.
DESARROLLO
Oído externo
El ectodermo del primer surco branquial forma el conducto auditivo externo y su
epitelio. El endodermo del primer arco branquial forma el martillo y el yunque, mientras
que a partir del segundo se forma el estribo. El endodermo de la primera bolsa faríngea
forma la trompa de Eustaquio, el oído medio y su epitelio. Los cartílagos, huesos y
músculos derivan del mesénquima que rodea los epitelios correspondientes.
289
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Oído interno
El ectodermo lateral al miencéfalo forma una invaginación par, la vesícula ótica
(otocisto), bajo control del FGF3 secretado por rombómeros 5 y 6. Da lugar al epitelio
del laberinto membranoso del oído interno.
El desarrollo está controlado por el gen Pax-2, que controla la región vestibular
dorsal y de la región coclear ventral, y los genes Prx1 y Prx2, que van a controlar la
formación de los conductos semicirculares.
Las células neuroepiteliales forman crestas ampollares, máculas y el órgano
espiral. La cresta neural origina melanocitos de la estría vascular y células de Schwann
del ganglio estatoacústico. El mesénquima va a formar la cápsula ótica y el laberinto
óseo.
290
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
OÍDO EXTERNO
Pabellón auricular
Apéndice ovalado en la superficie lateral de la cabeza. Es vestigial. Se encarga
de localizar, captar y amplificar el sonido y determina la dirección de la onda sonora.
Está revestido por piel fina con vello, glándulas sebáceas y sudoríparas apoyada
sobre cartílago elástico o hueso. Los músculos de la mímica son muy reducidos en esta
zona.
Las glándulas de esta localización forman cerumen, mezcla de secreciones y
restos celulares. Se encarga de lubrificar (reviste los pelos y el tímpano),
impermeabiliza (protege de la desecación) y protege frente a agentes externos
(insectos). Si hay un exceso de cera hay hipoacusia.
Conducto auditivo externo
Tubo en S itálica (lo que quiere decir una línea con dos pequeñas onditas, pero
debe ser que prefieren complicarse la vida), de 25 mm.
Porción externa
Tercio proximal. Piel fina con abundantes folículos pilosos (pelos sobre trago),
glándulas sebáceas y ceruminosas. Tiene cartílago elástico en continuidad con el de la
oreja.
Porción interna
Dos tercios distales. La piel es fina, con folículos pilosos y glándulas. Está
dentro del hueso temporal y termina en el tímpano o membrana timpánica.
OÍDO MEDIO
Especio aéreo en el hueso temporal (cavidad timpánica). Su mucosa está
formada por epitelio cúbico bajo y una lámina propia delgada, sin glándulas.
Encontramos tres huesos y dos músculos.
Límites:
x Anterior: inicio de la Trompa de Eustaquio
x Posterior: Hueso esponjoso de la apófisis mastoides (antro y celdas mastoideas).
x Lateral: Tímpano
x Medial: Pared ósea del oído interno, con dos orificio: ventana oval (a altura
vestibular) y membrana redonda (a altura coclear).
291
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Trompa de Eustaquio
Conducto estrecho y aplanado, de 3,5 cm. La porción proximal es ósea, formada
por el hueso temporal, mientras que después encontramos cartílago elástico y en la zona
distal cartílago hialino. Está formada por epitelio respiratorio.
La amígdala tubárica se sitúa en su extremo faríngeo. Puede producirse una otitis
media por diseminación de la faringe.
La luz queda ocluida, solo se abre cuando se produce un bostezo o la deglución.
Permite la entrada de aire en el oído medio e iguala la presión atmosférica con la del
oído medio.
Antro y celdas mastoideas
Espacios aéreos en apófisis mastoides del hueso temporal. Tienen epitelio cúbico
plano. Por diseminación de la otitis desde el oído medio se puede producir una
mastoiditis.
Tímpano
Membrana semitransparente delgada, inserta en el surco timpánico óseo a través
de cartílago fibroso.
Se divide en dos zonas:
x Porción flácida: Superior y pequeña. Más delgada y laxa.
x Porción tensa (100 micras): Inferior, grande, distendida y fusionada con el
manubrio del martillo. El ombligo es una depresión. Se encarga de mantener el
tímpano estirado.
Está formado por piel del conducto auditivo externo y mucosa del oído medio,
entre las que hay una capa intermedia de conjuntivo fibroso.
La piel es delgada, no queratinizada y sin pelos ni
glándulas.
La capa intermedio representa la mayor parte del
grosor, con haces densos de fibras colágenas radiales
externas (periféricas) y circulares internas (centro)
Además, hay fibras elásticas subepiteliales y pequeños
vasos sanguíneo y linfáticos y fibras nerviosas aferentes.
La mucosa del oído medio es un epitelio cúbico
plano, con parches de epitelio pseudoestratificado con
células ciliadas, secretoras, principales y basales que
originan una capa de moco que va a ser batida por los
cilios hasta el conducto auditivo.
292
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Las ondas sonoras hacen vibrar al tímpano. Puede perforarse de forma temporal
o permanente.
Cadena de huesecillos
Tejido óseo compacto, revestido por epitelio del oído medio. Están conectados
por articulaciones sinoviales. Se encarga de conectar el tímpano con la ventana oval,
pasando por martillo, yunque y estribo. Transmite energía sonora.
Músculos
El músculo del matillo, el tensor del tímpano, se inserta en el manubrio del
martillo y esta inervado por el trigémino.
El músculo del estribo es el estapedio, es menor y se inserta en la cabeza del
estribo. Amortigua el movimiento del estribo en la ventana oval. Está inervado por el
facial.
Modulan el movimiento del tímpano y modifican la amplificación de las ondas.
En caso de que la amplitud sea muy elevada la reducen mediante el reflejo de
atenuación, protector frente a sonidos de alta intensidad: la contracción de ambos
músculos hace que la cadena sea más rígida y los sonidos se transmitan peor.
OÍDO INTERNO
Hay dos compartimentos laberínticos, uno dentro de otro:
x Laberinto óseo: Cavidades o conductos intercomunicados en la porción petrosa
del temporal.
x Laberinto membranoso: Sacos o conductos comunicados dentro del laberinto
óseo.
Contiene tres espacios llenos de líquido:
x Endolinfáticos: dentro del laberinto membranoso. Revestidos por epitelio sobre
la lámina basal. Contienen endolinfa, similar al líquido intracelular (K+).
x Perilinfáticos: entre los dos laberintos. Espacios estrechos de conjuntivo pobre
en fibras, con melanocitos. Contienen perilinfa, similar al líquido extracelular
(Na+, Ca2+, Cl-, HCO3-). Similar al líquido cefalorraquídeo.
x Cortilinfáticos: Dentro del órgano de Corti. Contienen cortilinfa, similar al
líquido extracelular.
Hay un equilibrio iónico entre endolinfa y la perilinfa, con una diferencia de
potencial de +80 mV. El aumento del volumen de Linfa produce la enfermedad de
Menière: vómitos, naúseas, pitidos, mareos...
293
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Laberinto óseo
El laberinto óseo está dentro del hueso temporal y contiene perilinfa y al
laberinto membranoso. Posee 3 conductos semicirculares, el vestíbulo y la cóclea o
caracol.
El vestíbulo es una cavidad ovalada central, hacia atrás suya se extienden los
conductos semicirculares y hacia delante la cóclea. La pared lateral contiene la ventana
oval.
Los conductos semicirculares son
3: superior, posterior y lateral o externo,
perpendiculares entre sí, ocupando tres
planos (sagital, frontal y horizontal).
Junto al vestíbulo se dilatan formando
ampollas. Salen del vestíbulo por 5
orificios (rama ósea común para
conductos superior y posterior). Hay
cinco porque cada conducto entra y sale,
siendo una común.
La cóclea o caracol forma 2
vueltas y ¾ desde la base al vértice. La
columela o modiol es la zona de hueso
esponjoso central (eje), en cuyo interior
está el ganglio espiral o de Corti. En su
base se encuentra la ventana redonda,
cubierta por la membrana timpánica secundaria.
294
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Laberinto membranoso
Suspendido en la perilinfa, con endolinfa en su interior. Se divide en laberinto
vestibular y coclear.
El laberinto vestibular está
formado por tres conductos
semicirculares membranosos dentro
de los conductos semicirculares
óseos. Hay dos sacos: utrículo y
sáculo, conectados por el conducto
utriculosacular membranoso. Además,
hay un conducto endolinfático que
sale del conducto utrículosacular, sale
del oído y acaba en un
ensanchamiento, el saco
endolinfático, que acaba en la
durameadre.
El laberinto coclear tiene un conducto coclear dentro de la cóclea, la cual recorre
por el centro formando la rampa intermedia. Acaba en el vértice. Está unido a la lámina
espiral (columela) y es continuo con el sáculo mediante el conducto reuniens. Por
encima queda la rampa vestibular, que comienza en la ventana oval. Por debajo queda la
rampa timpánica, que comienza en la ventana redonda. Estas dos rampas están
comunicadas por el helicotrema. Las rampas vestibular y timpánica contienen perilinfa.
295
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Membrana de Reisner
Se trata de una membrana constituida por un doble epitelio, es decir, que está
organizada en dos capas superpuestas entre las que queda un espacio mínimo con tejido
conjuntivo:
x Mesotelio: se encuentra en contacto con la perilinfa de la rampa vestibular
x Epitelio plano simple: en contacto con la endolinfa de la rampa intermedia
Las células mesoteliales son más altas y convexas que las células epiteliales
planas, y en su citoplasma basal encontramos una masa electrodensa que se corresponde
con el núcleo. Otra característica diferenciadora es que las células epiteliales tienen en
su membrana apical, esto es, la que está en contacto con la endolinfa de la rampa
intermedia, prolongaciones a modo de microvellosidades, que en la imagen observamos
como si fuesen gotas.
Estría vascular
Descansa sobre el ligamento espiral, más basófilo, que contacta con la lámina
propia. Está formada por un epitelio que, al contrario de lo que sucede con normalidad,
se encuentra vascularizado, y que se combina con tejido conjuntivo (lámina propia).
Este epitelio es de tipo pseudoestratificado, y en él encontramos núcleos a tres alturas,
correspondientes a tres tipos celulares diferentes:
x Células basales: se encuentran en la zona
más profunda del epitelio. Poseen
prolongaciones que les permiten establecer
contactos entre sí y con los fibroblastos del
tejido conjuntivo
x Células intermedias: situadas en la zona
media del epitelio
x Células marginales: se hallan en contacto
con la luz, y entre ellas hay una gran
cantidad de complejos de unión. Tienen
cortas microvellosidades en la membrana
apical y una gran cantidad de mitocondrias,
pero su característica más distintiva es su
laberinto basal, formado por repliegues de
la membrana.
296
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Por debajo del epitelio, como en cualquier otro, hay una lámina propia. Existe
también, especialmente en la infancia, una lámina basal muy tenue situada entre la
lámina propia y el epitelio, que en el adulto desaparece. En el tejido conjuntivo de la
lámina propia es muy frecuente encontrar melanocitos, cuya función a este nivel no está
clara, pero que se relacionan con patologías auditivas como el síndrome de Wanderburg,
una sordera congénita asociada con defectos en la pigmentación y en la interacción del
receptor c-kit con el factor de células madre.
Los capilares que encontramos en la estría vascular son continuos, por lo tanto
poseen pericitos. En realidad, las asas capilares se originan a nivel de la LP, pero acaban
por introducirse en el epitelio, lo cual hace que esté vascularizado.
Funciones de la estría vascular
x Síntesis de endolinfa y excreción de iones K+, claves para la generación de
diferencias de potencial. Está función de la estría se puede ver afectada por:
o Mutación del gen ISK: modifica un canal de K+ y con ello provoca una
alteración en la producción de endolinfa y la degeneración del órgano de
Corti, en cuyo metabolismo interviene también la estría vascular
o Mutación del gen codificante para la conexina 26 presente en las células de
Deiters, que interfiere con el reciclaje del K+ y también conlleva la
degeneración del órgano de Corti
x Metabolismo del órgano de Corti.
Receptores sensitivos
Se trata de mecanorreceptores que transducen energía mecánica a eléctrica, y
que encontramos agrupados en 6 regiones concretas del laberinto membranoso:
x 3 en las crestas ampollares
x 2 en las máculas
x 1 en el órgano de Corti
Sistema vestibular
En él, el tipo celular principal son las células ciliadas. Tienen dos características
importantes para su función:
x Un solo cinocilio grande y ancho (50 micras), pero inmóvil.
x Estereocilios muy organizados que se disponen formando una V o una W, cuyas
alturas son distintas y van en escala, como en disposición escalonada. Existen
enlaces apicales y laterales (estos últimos más inferiores) que unen a los
diferentes estereocilios entre sí. A nivel basal, los MF de actina del eje se anclan
en la placa cuticular, y a nivel apical la punta de los estereocilios está incrustada
en una matriz gelatinosa.
297
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Podemos hacer una sistematización de las
células ciliadas, clasificándolas como:
x Ciliadas tipo 1 o piriformes: son altas
pero están ensanchadas, especialmente en
su base (fofisanas o rechonchitas), donde
se encuentran rodeadas por un cáliz que
constituye una terminación nerviosa
aferente, con la que a su vez sinaptan
terminaciones eferentes.
x Ciliadas tipo 2: son células prismáticas
sobre las que terminan fibras aferentes y
eferentes, de manera que se postula que
las eferentes modularían la cantidad de
información que las aferentes transmiten
al SNC. Sus estereocilios con más cortos
y poseen una gran cantidad de REL.
Sistema auditivo
Del mismo modo, está formado por células ciliadas, pero el único cilio del que
disponen se pierde en el proceso de desarrollo, manteniéndose solamente el cuerpo
basal del mismo. Por lo demás, sus características y las de sus estereocilios son iguales
a las de las células ciliadas del sistema vestibular, con matices según la subclase, ya que
tenemos dos tipos de células ciliadas:
x Internas: sus estereocilios no están incrustados en la matriz gelatinosa
x Externas: sus estereocilios sí se hallan incrustados en la matriz gelatinosa
Así, en la mayoría de los mecanorreceptores los extremos apicales de los
estereocilios y cinocilios están incrustados en la MEC gelatinosa opuesta al epitelio.
Cuando la matriz gelatinosa se desplaza los estereocilios se inclinan, y en este proceso
pueden alejarse o acercarse del quinocilio:
x Acercamiento: el cinocilio se hunde y se
abren canales de Na, Ca y K, lo que produce
la despolarización de la membrana y la
transmisión de un impulso nervioso
excitatorio (los neurotransmisores empleados
son Glu y Asp).
x Alejamiento: el cinocilio se eleva y la
membrana se hiperpolariza, con lo que se
inhibe el impulso.
Como ya hemos avanzado, encontramos
mecanorreceptores agrupados en 6 regiones
concretas del laberinto membranoso:
298
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x 3 en las crestas ampollares
x 2 en las máculas
x 1 en el órgano de Corti: porción auditiva
Receptores vestibulares
Las 3 crestas ampollares y las 2 máculas
forman el aparato vestibular, que registra los
cambios de posición, movimientos de la cabeza y
aceleraciones, en definitiva, la orientación
espacial.
Crestas ampollares
Se encuentran en las ampollas de los conductos semicirculares, cuyo epitelio
presenta un engrosamiento que esta buena mujer ha calificado como cresta de hippie
(creo que se “está burlá” con hippie y punk), y que es perpendicular al eje del conducto.
En las crestas ampollares encontramos dos tipos de células:
x Células de sostén: no tienen cilios ni estereocilios, pero sí microvellosidades
cortas, y forman una especie de depresión donde se ubica la célula ciliada.
x Células ciliadas: unidas las de sostén y recubiertas por una matriz gelatinosa que
llamamos cúpula. La cúpula es una matriz gelatinosa rica en proteínas y
azúcares en la que hallamos excavados unos conductos alargados donde se
introducen los estereocilios de las células ciliadas. Su extremo no está libre, sino
anclado al epitelio opuesto por proteínas como la otogelina, y bañado en
endolinfa. Encontramos dos tipos de células ciliadas:
o Tipo I: predominantes en la zona apical.
o Tipo II: predominantes en la zona basal.
Las crestas ampollares actúan como
receptores de los movimientos rotacionales:
cuando giramos la cabeza giran el laberinto óseo y
el membranoso, pero la endolinfa se mueve con un
cierto retardo, y esta diferencia de velocidades
hace que la cúpula empiece a desplazarse y con
ella los estereocilios, que en esa flexión se pueden
acercar o alejar del cinocilio dando como resultado
las dos respuestas ya comentadas. En cualquier
caso, el proceso está muy relacionado con la
información visual (sí, probad a cerrar los ojos a
ver qué tal): el laberinto cinético regula
básicamente los movimientos oculares, de modo
que los movimientos oculares rápidos al girar la
cabeza dependen de la inclinación de la cúpula.
299
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Máculas
Las hallamos en utrículo y sáculo. Poseen un epitelio muy sellado con los
mismos tipos celulares que las crestas, y también con engrosamientos. Ambas máculas
son perpendiculares entre sí: en posición erecta la mácula del utrículo es horizontal y la
del sáculo es vertical.
Las células ciliadas se encuentran en este caso cubiertas por la membrana
otolítica, en la que se hallan incrustados los estereocilios. Se trata de una masa
gelatinosa formada por azúcares en cuya porción externa encontramos los otolitos o
cuerpos otolíticos, que son cuerpos cristalinos de carbonato cálcico y proteínas.
En la zona media de la mácula parece que vemos una línea imaginaria
correspondiente a una zona hundida, llamada estriola, que permite dividir las máculas
en dos mitades simétricas. En el utrículo los estereocilios se orientan con las puntas
hacia ella, y en el sáculo con las puntas hacia afuera.
300
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Los de las máculas son mecanorreceptores que registran la posición de la cabeza
en reposo, es decir, el efecto de la gravedad sobre la misma, o el desplazamiento lineal:
del mismo modo, se produce el desplazamiento del laberinto óseo y el membranoso,
pero la endolinfa se mueve con un cierto retardo, y esta diferencia de velocidades hace
que la cúpula empiece a desplazarse y con ella los estereocilios, que en esa flexión se
pueden acercar o alejar del cinocilio dando como resultado las dos respuestas ya
comentadas.
Órgano espiral de Corti
Contiene receptores para la audición y va ascendiendo a lo largo del conducto
coclear, recorriendo el conducto auditivo desde su base hasta la cúpula. Dentro de él
podemos encontrar tres espacios tubulares rellenos de cortilinfa:
x Túnel externo: su suelo es la
membrana basilar de la rampa
intermedia.
x Túnel interno o de Corti:
polariza el órgano de CortiÆ a
un lado del mismo tendremos la
parte externa y a otro la interna.
No se asienta sobre la membrana
basilar.
x Espacio de Nüel.
El epitelio va a estar formado:
x Células sensitivas: ciliadas
x Células de sostén: pueden ser
falángicas o de los pilares
x Otras células: interdentales, de
Claudius, de Böttcher y de
Hensen.
También vamos a encontrar a este nivel la membrana tectoria, con una sustancia
amorfa rica en colágeno II, V, IX y XI, queratinas y tectorinas, segregada por las células
interdentales y adherida a la columela en su extremo medial, mientras que en el lateral
se halla libre sobre el órgano de Corti.
301
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Este órgano forma parte del conducto coclear situado a lo largo formando la
rampa intermedia. En él encontramos Mecanorreceptores auditivos y tres espacios que
son alargados o tubulares, al igual que el órgano, y que están llenos de líquido,
cortilinfa. Son el túnel interno o de Corti, el espacio de Nüel y el túnel externo. Todos
son importantes pero el primero es el punto que nos permite distinguir entre la parte
interna y externa del órgano de Corti, es decir, tiene gran importancia la hora de
localizar las estructuras en este órgano. Desde el túnel de Corti hacia el hueso hablamos
de parte interna y hacia la estría vascular de parte externa.
El órgano de Corti se forma por un epitelio asentado sobre la membrana basilar.
Muestra:
x Células Sensitivas Ciliadas: Pueden ser internas o externas y esto viene definido
por el túnel interno o de Corti. También hay dos espacios tubulares.
x Células de Sostén, de dos tipos:
o Falángicas.
o De los pilares.
Existen además otro tipo de células: interdentales, de Claudius, de Böttcher y de
Hensen (menores). Existe también una matriz gelatinosa, la membrana tectoria.
Células Ciliadas
Las células ciliadas son un tipo de
mecanorreceptores que encontramos a lo
largo del conducto coclear y no se clasifican
en tipo I y II (eso es para la porción
vestibular) sino en internas y externas.
Las internas están dispuestas en
forma de eje. Son una única hilera a lo largo
de todo el conducto, en el lado interno cerca
de la lámina espiral, que están
completamente rodeadas por unas células de
sostén, las falángicas internas.
En cambio, las ciliadas externas forman entre 3 y 5 hileras, dependiendo el
número de filas del punto concreto que miremos del conducto coclear (base, punta…).
Además, las externas no están completamente rodeadas por las falángicas externas sino
que sólo rodean su base. Tienen un gran desarrollo de REL denominado inicialmente
cuerpo de Hensen, cisternas subsuperficiales y una enorme cantidad de estereocilios
alineados (que apuntan hacia la estría vascular).
Las células ciliadas son células con estereocilios organizados en V y estos
estereocilios se organizan de forma decreciente desde el cinocilio.
302
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Los estereocilios se orientan hacia la estría vascular y el crecimiento del eje de
filamentos de actina está controlado por la miosina XVa y la whirlina. Además, este
crecimiento se estabiliza por otra proteína denominada radixina.
Este eje sigue anclado a la placa y los estereocilios están unidos entre sí y
pueden encontrarse en dos situaciones:
x Apicales, en los extremos (de un estereocilio a otro).
x Laterales, entre los estereocilios (une estereocilios lateralmente).
En estos enlaces participan miosina VIIa (en los enlaces laterales) y miosina Ic
(en los enlaces apicales) y cadherina 23 (apical también).
Estos enlaces son importantes porque dan cohesión al conjunto de estereocilios y
así puedan trabajar de manera uniforme. Puede haber mutaciones en estos enlaces
causando patologías como:
303
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Síndrome de Usher: el paciente tiene
hipoacusia neurosensitiva por defectos en
las proteínas de estos enlaces (sí hay
estereocilios pero están mal organizados).
x Otro defecto hipoacústico consistiría en
una desorganización de los haces, lo que
se conoce como retinitis pigmentosa.
Estos enlaces no solo dan cohesión, sino
que tienen que ver con la apertura y cierre de
diferentes canales iónicos (sobre todo las
cadherinas y miosinas apicales) lo que asegura y
favorece que haya una respuesta uniforme y
sincronizada, como el canal de calcio TRPA1 que
asegura una respuesta uniforme. Estos canales son
los canales iónicos MET o de transducción
mecanoeléctrica. Lógicamente el canal de Ca2+
contribuye a que haya una descarga más uniforme.
Cuando los estereocilios se flexionan hacia donde estaría el cinocilio (que no
está) los canales se abren produciéndose una despolarización y con ello un impulso
eléctrico. Los movimientos hacia el estereocilio más alto hacen que se abran los canales,
lo que permite la entrada de K+ produciendo la despolarización de la célula. Como
consecuencia entra Ca2+ lo que hace que se libere el neurotransmisor. Cuando la flexión
es alejándose lo que ocurre es que los canales se cierran en caso de estar abiertos
produciéndose la hipopolarización.
No todas las células ciliadas tienen los estereocilios de la misma longitud, sino
que en las porciones más basales del caracol las células ciliadas tienen estereocilios más
cortos y según avanzamos en el conducto hacia la punta del caracol los estereocilios de
las células ciliadas son más largos. Luego según avanzamos las células ciliadas tienen
más estereocilios y más largos
Una diferencia entre las células ciliadas internas y externas, es que en las
internas las puntas de los estereocilios no se incrustan en ninguna matriz, mientras que
en externas sí. Además, los estereocilios de las células ciliadas internas están bañados
por endolinfa mientras que los de las externas están embebidos en una membrana en
contacto con la membrana tectoria.
Células Falángicas
Al igual que en el caso de las células ciliadas, encontramos células falángicas
internas y externas (o de Deiters).
Las células ciliadas internas formaban una única interna así que las falángicas
internas, que envuelven a las anteriores, también. Cada una envuelve una ciliada interna
304
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
por completo, entonces las células ciliadas y las terminaciones nerviosas que sinaptan
con estas células ciliadas quedan rodeadas por las falángicas internas.
Por otro lado, las ciliadas externas formaban de 3 a 5 hileras por lo que vamos a
encontrar también de tres a cinco filas de falángicas externas que las rodean. Estas
falángicas externas también se denominan células de Deiters.
Las falángicas internas rodean totalmente la ciliada pero las externas solo rodean
la parte más basal, así como a las terminaciones nerviosas.
La célula de Deiters, además de rodear solo la parte más basal, emite una
prolongación ascendente denominada falange que en un cierto punto se aplana, llegando
a la altura más apical de la célula ciliada externa y contactando con las prolongaciones
adyacentes formando una estructura hermética que se sella y se denomina
lamina/membrana reticular o placa cuticular apical.
Luego la membrana es una estructura formada por las falángicas que va a
permitir sellar un espacio bajo esta placa. Y ese espacio está lleno de cortilinfa. Debajo
de la placa encontraremos la endolinfa.
Esta membrana sella y permite aislar completamente al compartimento inferior
del superior. Así, por encima de la placa tendríamos endolinfa (pues el órgano de Corti
formaba parte del conducto coclear) y por debajo, cortilinfa. Estas células se
encuentran unidas entre sí y con las ciliadas. Es decir, la lámina reticular señala el
espacio interno del órgano de Corti.
Células de los Pilares
Además de las células falángicas
tenemos otras células de sostén como las
de los pilares que delimitaban el túnel de
Corti o interno, de modo que hay células
del pilar interno y del externo.
Son células altas, con citoplasma
estrecho, zona apical y base muy
ensanchadas y zona media estrecha. El
citoplasma es estrecho y como tiene que
305
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
dar sostén a esa estructura tan alargada, se necesita un gran desarrollo de microtúbulos
en el citoesqueleto (especialmente abundante en la zona media). El núcleo se aloja en la
zona más basal, así como los orgánulos. Estas células sólo contactan entre sí por las
porciones apicales y basales.
Las bases de las células contactan con la célula adyacente y con la célula del
pilar opuesto y en la zona media deja huecos por donde pasan las fibras nerviosas. Es
importante saber también que solo la parte externa del órgano de Corti (desde el pilar
externo) se apoya sobre la membrana basilar y en cambio el órgano de Corti interno se
apoya en la lámina espiral (sobre una parte denominada labio timpánico)
Así, vistos estos tipos de células,
podemos decir que el túnel interno está
delimitado por las células de los pilares. El
túnel de Nüel es un espacio también
tubular que está delimitado por las células
del pilar externo y las células de Deiters
(siendo externo al pilar externo) y, por
último, el túnel externo está delimitado
por células de Deiters y otras células de
sostén que están más hacia la estría
vascular (surco espiral externo). Estos tres
túneles están rellenos de cortilinfa.
306
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Células Interdentales
Son células epiteliales con forma de botella o de pera, que emiten una
prolongación apical ascendente que se va a aplanar y que va a tapizar el surco espiral
interno. Se sitúan en la zona más interna del órgano de Corti, sobre el limbo espiral
(zona hundida o depresión que está excavada en el conjuntivo).
Las prolongaciones que se aplanan contactan unas con otras y por medio de ellas
las células segregan unas gotas que contienen moléculas que formaran la membrana
tectoria (fundamentalmente proteoglucanos). En este conjuntivo las fibras y las células
tienen una disposición perpendicular a la superficie y como son células epiteliales
tenemos una lámina basal que rodearía por completo a cada célula.
Estas células se continúan con la membrana de Reissner y tienen el soma dentro
de una especie de cavidad o túnel, donde se pueden alojar varias células, que estaría
excavada en el conjuntivo del limbo espiral (es decir, que se asientan sobre él). Cuando
se alojan más células en una cavidad, se produce una comunicación directa conocida
como cráter central. Aparecen unas 2 o 3 células por cavidad. Además, tienen una gran
cantidad de orgánulos y gránulos que tienen que ver con la síntesis. Hay abundantes
lisosomas.
Las células interdentales están en medio de un conjuntivo denso de fibras y
fibrillas de colágeno dispuestas verticalmente por lo general. Los fibroblastos de este
conjuntivo se disponen debajo de las interdentales.
Estas células tienen una porción ascendente que llegado el momento se aplana y
al final se queda con las células vecinas, encontrándose completamente sellado.
Membrana Tectoria
La membrana tectoria es una sustancia amorfa y gelatinosa rica en proteínas en
la que intervienen colágenos II, V, IX y XI y, además, distintas queratinas, otogelina (ya
mencionada en las máculas y crestas en la cúpula y membrana otolítica) y la α y β
tectorina.
Las células interdentales son las que segregan los diferentes tipos de colágeno.
Todos los componentes de la membrana son segregados como gotas y es entonces
cuando se forma la membrana tectoria.
La porción medial de la membrana esta fija unida a la columnela y la lateral cae
libremente sobre el órgano de Corti.
Si observamos esa membrana tectoria vemos que paralela a ella, bajo su
superficie, se encuentra una banda de Hensen que es más basófila y que corre
longitudinalmente. Esta banda se transforma en una capa más superficial que termina
formando la red marginal la cual está formada por finas trabéculas de aspecto gelatinoso
307
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
que permitirían anclar el extremo lateral a unas células de sostén llamadas células de
Hensen (que son más externas que las células de Deiters)
Estas células de Hensen empiezan siendo prismáticas, pero van disminuyendo y
tapizan el surco espiral externo hasta la estría vascular.
Células de Hensen
Se encuentran externamente a las células de
Deiters y por tanto van desde las células de Deiters
hasta la estría vascular, tapizando el surco espiral
externo. Son de morfología cúbica-prismática.
Otras Células
En otras especies (no se sabe si en la humana también) se asientan sobre la
basilar las células de Böttcher (tapizando el surco espiral externo) y de Claudius, estas
últimas en contacto con la estría vascular.
Características del Suelo del Órgano de Corti
Sostiene al órgano de Corti (no a todo) al ser su superficie. Esta membrana se
extiende hasta una lámina basal y un conjuntivo que tiene una capa de fibroblastos, de
aspecto epiteloide, los cuales están en contacto con un revestimiento de células planas,
formadas también por este conjuntivo, denominadas perilinfáticas pues constituyen la
perilinfa. Este es el límite superior. Bajo ellas no hay lámina basal.
En este conjuntivo, además, se observan filamentos de 10nm que forman una
matriz amorfa, rica y abundante y que se organizan en paralelo los unos con los otros,
formando dos estratos, de tal manera que los filamentos adquieren una orientación
transversal con respecto al órgano de Corti. Los filamentos se asocian formando las
fibras basilares o cuerdas acústicas, de las cuales tenemos unas 20.000 a lo largo del
conducto coclear. El conjuntivo está inervado con fibras nerviosas amielínicas y
vascularizado.
Estos filamentos o fibras basilares no tienen longitud o diámetro fijo, sino que
varía según nos vamos desplazando a lo largo del conducto de tal manera que en la base
las cuerdas acústicas son más cortas y según avanzamos, van aumentando de longitud
(40 μm a 500 μm).
Con el diámetro sucede al contrario, siendo mayor en la base y menor en el
ápice. Esto es importante porque en función del diámetro y longitud cada fibra acústica
o basilar tiene una rigidez distinta, lo que supone que los distintos sonidos afecten sobre
todo a ciertas fibras y menos a otras, es decir, en función de la frecuencia del sonido se
produce una vibración máxima en cierta parte de la membrana basilar, no toda vibra
igual.
308
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Al principio se pensó que las cuerdas eran algún tipo de colágeno, pero en
realidad son fibras elásticas. Como la longitud y el grosor varían, cada tipo de
frecuencia de sonido provoca vibración en todas ellas y solo unas pocas van a conseguir
vibrar al máximo.
De manera que a cada tipo de sonido, según su frecuencia le corresponde una
zona concreta de vibración máxima.
Percepción del Sonido
La energía sonora viaja por el aire en vez de pasar al líquido viscoso (perilinfa)
que está en el compartimento óseo. Cuando por fin se transmite al medio líquido, la
mayor parte de esa energía se pierde (impedancia acústica) quedando solo un 0,1%. El
oído medio es un mecanismo que trata de reducir esa pérdida de energía, aumentando
ese 0,1%, a través de la reflexión de las ondas sonoras.
Las ondas sonoras cuando entran en el oído interno van a provocar la vibración
de la perilinfa. Estas vibraciones tienen que llegar a las células ciliadas que se
encuentran en un compartimento óseo que no es deformable.
La energía sonora debe pasar a un medio líquido para poder alcanzar las células
sensitivas, los mecanorreceptores están dentro de un compartimento óseo que es
incompresible y es líquido. El compartimento tiene dos orificios: la membrana oval (de
entrada) y la redonda (salida), que actúa como válvula de escape.
Así, la energía que entra en el oído interno tiene que volver a salir. Se capta por
el oído externo, alcanza el tímpano en el oído medio y las vibraciones aéreas se
transforman en mecánicas-timpánicas, que se transmiten por la cadena de huesecillos.
El estribo golpea entonces la membrana oval que se encontraba en la pared ósea.
Las vibraciones mecánicas pasan a transmitirse al líquido que tenemos en el
vestíbulo (perilinfa) de la rampa timpánica y aparece una vibración hidráulica que se
comunica por el helicotrema con la rampa timpánica, es decir, las dos rampas, vestibular
y timpánica se comunican por el helicotrema, por lo que cuando comienza a vibrar se
transmite a la perilinfa de la rampa timpánica (vibran ambas rampas) lo que produce
movimientos en la membrana timpánica secundaria.
Cuando la perilinfa vibra en la rampa timpánica, el techo de esta rampa es la
membrana basilar y también vibra; dando lugar a ondas vibratorias.
309
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Es importante saber que en función de la frecuencia del sonido la membrana
basilar que vibra más es una en concreto, en función de la longitud de la cóclea. Si el
sonido tiene alta frecuencia la zona que más va a vibrar va a estar en la zona más
inferior y los sonidos de baja frecuencia producen una vibración máxima en la
membrana basilar más próxima al vértice, teniendo entonces toda una escala. Esto
supone que la membrana basilar, conducto coclear y órgano de Corti muestren una
diferenciación topográfica. Esta diferenciación se transfiere al SNC de manera que en la
corteza auditiva los sonidos no se registran en toda la zona de la corteza, sino que
determinados sonidos van a determinados puntos de la corteza.
Cuando la membrana basilar vibra, evidentemente afecta a lo que está encima de
ella, es decir, al órgano de Corti (solo a la parte que se apoya en esa membrana basilar).
La zona interna, en cambio, tiene células envueltas por falángicas pero cuyos
estereocilios están rodeados de endolinfa y así, si la endolinfa vibra estos estereocilios
también.
En cambio, en la zona externa los estereocilios no está en contacto con la
endolinfa, sino que estos estereocilios están embebidos en la membrana tectoria. El
hecho de que en un caso haya membrana tectoria y en otra no supone una diferencia
importante en la membrana funcional porque en las externas se producen fuerzas en
direcciones contrarias lo que provoca fuerzas de cizalla sobre los estereocilios. Esto
obliga a los estereocilios a flexionarse pudiendo despolarizarse o hiperpolarizarse la
membrana. Si no hubiera fijación de la tectoria, no habría fuerzas de cizalla.
Lo que se consigue con las cuerdas es que haya una diferenciación topográfica.
Esto se correlaciona con las diferentes zonas en las que se divide la corteza cerebral y
con ello decimos que la división no es más que el traslado de la división topográfica al
cerebro.
Cuando la membrana basilar vibra, vibra todo lo que se apoye en ella: las células
falángicas y las ciliadas. Luego hay un baile en el que se van moviendo todas las células
dependiendo del movimiento. Quien no tiene los estereocilios incrustados en la
membrana tectoria son las células ciliadas internas. Estas no se ven afectadas puesto que
los estereocilios están bañados por la endolinfa.
Esto significa que las células ciliadas externas potencian la respuesta al estímulo
sonoro que fundamentalmente es captado por las ciliadas internas. Para ayudar a la
potenciación tenemos también una diferencia de potencial (80mV normalmente) de
150mv entre endolinfa y perilinfa
La diferencia de potencial es clave para que la respuesta de las internas se vea
potenciada y podamos oír. La diferencia de potencial se crea básicamente por los iones
K+ que se segregan en la estría vascular.
310
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Clínica
Causas posibles de sordera: Pueden deberse a una mutación en el gen Isk. Isk se
expresa en la estría vascular; al mutar, se produce alteración del canal que permite la
secreción de K+, de modo que la endolinfa generada no es normal, lo que hace que el
órgano de Corti degenere. El gen para la conexina 26 también está implicado en la
secreción de potasio, se expresa en las células falángicas; si está mutado, hay reciclaje
defectuoso de potasio y sordera. El gen de la α-tectorina también produce hipoacusia.
Hay otro tipo de sordera causada por infecciones víricas, ruido sostenido y
elevado y la vejez. Este último causado por muerte o atrofia de células ciliadas externas,
perdiendo la capacidad de oír el sonido agudo.
Inervación
El oído interno está inervado por el par VIII con su porción vestibular, que
inerva los receptores vestibulares, y su porción coclear, que inerva los receptores
auditivos.
311
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Porción Vestibular
Las neuronas están situadas en el ganglio vestibular o de Scarpa, marcando sus
dendritas hasta los receptores vestibulares (acaban junto a las células ciliadas I/II. Se
produce sinapsis en cinta. Los axones se dirigen al SNC, acabando en el tronco
encefálico, concretamente dentro de los núcleos vestibulares, aunque un pequeño
porcentaje de fibras se dirigen al cerebelo y terminan en los núcleos de los pares II, IV,
y VI; que tienen neuronas que inervan la musculatura ocular.
Porción Coclear
Aquí encontramos neuronas situadas en el ganglio espiral o de Corti.
Encontramos neuronas de dos tipos:
x Las neuronas tipo I (90% neuronas son de tipo I) son muy grandes, más
abundantes, de tipo bipolar, sus dendritas se dirigen a la cóclea e inervan
prácticamente solo células ciliadas internas. Cada ciliada interna es inervada por
10 de estas neuronas.
x Las neuronas tipo II son más pequeñas, bipolares, y sus dendritas se dirigen al
órgano de Corti, a las células ciliadas externas. Se produce sinapsis en cinta (de
la parte vestibular).
Los receptores auditivos son sólo las células ciliadas interna. Parece ser que las
externas son unas células capaces de modificar su forma de contraerse, por una parte, de
manera espontánea pero también como respuesta a un estímulo, lo que les permite a
ellas solas generar una energía en forma de emisiones otoacústicas o EOA,
intensificando la capacidad de respuesta de las ciliadas internas. Luego las células
ciliadas externas van a poder producir la energía acústica o estatoacústica en relación a
las relaciones que sufren.
Los axones de las neuronas I y II se dirigen al SNC, acaban en el tronco
encefálico, en los núcleos cocleares del bulbo raquídeo. Desde aquí, la información
pude seguir ascendiendo y alcanza el tálamo, en los cuerpos geniculados. La
información puede seguir hasta la corteza auditiva (lóbulo temporal)
Hay una inervación eferente, denominadas eferencias cocleares de Rasmussen,
que parten del cerebro y van paralelas a las aferentes, en el par VIII. Estas eferencias
sinaptan con las aferentes en el oído (procedentes de ciliadas internas y externas),
controlando la llegada al SNC de la información recogida; es decir, actúan modulando
la cantidad de información que son capaces de transmitir las aferencias.
Las eferencias incluyen las
neuronas de tipo I, de tipo II y las
cocleares, que son capaces de transmitir las
aferencias.
312
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
OJO
SISTEMA VISUAL
La fotosensibilidad es una característica inherente a todo ser vivo, tanto
unicelulares como pluricelulares (en estos hay formaciones laminares especializadas
denominadas retinas). La fotorrecepción tiene estructuras accesorias que forman ocelos,
ojos compuestos y el globo ocular.
En los vertebrados hay células especializadas, conos y bastones, y anejos.
DESARROLLO
x Neuroectordermo lateral del diencéfalo
x Ectodermo de la superficie de la cabeza
x Mesénquima
Unas evaginaciones laterales de la zona derecha e izquierda del diencéfalo
originan dos vesículas ópticas neuroepiteliales, cada una de las cuales sigue unida a la
pared del encéfalo a través de un tallo óptico hueco
313
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
La vesícula óptica induce cambios en el ectodermo próximo haciendo que se
invagine. La invaginación se cierra sobre sí misma y da lugar a la vesícula del cristalino,
que originará el cristalino.
La vesícula óptica empieza a su vez a invaginarse y forma una copa de doble
capa, que va a dar lugar a las dos capas de la retina: externa Æ epitelio pigmentado,
interna Æ retina neural, con una elevada capacidad mitótica para formar el resto de
capas.
El mesénquima origina el componente vítreo, la coroides y elementos fibrosos
de esclerótica y córnea, esta última inducida por el cristalino. Las células del
mesénquima forman estroma primario de la córnea, con colágenos I y II. Las células
endoteliales capilares emigran hacia el estroma primario y producen ácido hialurónico,
lo que determina aparición de edema en el estroma. Las células mesenquimales del
espacio circundante emigran hacia el estroma y secretan hialuronidasa. El estroma se
retrae y la córnea adquiere la forma y la transparencia adecuadas.
Posterior al cristalino, la capa coroides vascular genera el cuerpo ciliar, el
músculo ciliar y los procesos ciliares. Anterior al cristalino, la capa coroides forma el
estroma del iris. Los procesos ciliares secretan el humor acuoso, que se acumula
primero en la cámara posterior (entre el iris y el cristalino) y después pasa a la cámara
anterior (entre el iris y la córnea) tras atravesar la pupila. El humor acuoso sale de la
cámara anterior a través del conducto de Schlemm, una pequeña vena (seno venoso de
la esclerótica) que rodea al ojo de forma circular en la parte anterior de la coroides.
Alrededor del reborde de la copa óptica, las capas interna y externa forman el
epitelio posterior del cuerpo ciliar y el iris. Los músculos esfínter y dilatador de la
pupila se desarrollan a partir del epitelio posterior.
La capa neural de la retina se diferencia en células fotosensibles, neuronas
bipolares y neuronas ganglionares. Los axones de las neuronas ganglionares forman la
capa de fibras nerviosas de la retina, que convergen sobre el tallo óptico que ocupa la
cisura óptica formando el nervio óptico.
GLOBO OCULAR
Formación esférica, par, con un diámetro de 2,5 cm. El ojo puede autoenfocarse,
realizar ajustes según la intensidad de la luz y convertir la luz en impulsos eléctricos que
el encéfalo puede interpretar. En las personas el ojo se encuentra protegido dentro de la
órbita ósea y se conecta con el encéfalo a través del nervio óptico. El globo ocular
protege y facilita la función de la retina fotorreceptora, la capa más interna del globo
ocular.
Hay 6 músculos que permiten la rotación. Las fosas ópticas están en depresiones
craneales:
314
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Pareced óseas excepto parte anterior
x Tejdo adiposo
x Cápsula de Tenon de conjuntivo denso.
Funciones
x Fotorrecepción: Captan intensidad y color de la luz.
x Codifican los parámentros para su transmisión al SNC.
x SNC interpreta conjuntamente las dos imágenes que recibe.
x Visión binocular permite percibir profundidad y distancia.
Estructura anatómica
Capas concéntricas (3)
x Túnica fibrosa o esclerocórnea: esclerótica + córnea
x Túnica vascular o úvea: coroides + cuerpo ciliar + iris
x Túnica nerviosa o retina: pars pigmentosa + pars nervosa
Cámaras (3)
x Anterior: entre córnea e iris
x Posterior: entre iris y cristalino
x Cuerpo vítreo: tras cristalino
Medios ópticos de difracción (4)
x Córnea
x Humor acuoso: en cámaras anterior y posterior
x Cristalino
x Humor vítreo: en cuerpo vítro
315
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Túnica fibrosa
Esclerótica
5/6 posteriores. Capa dura, opaca y blanca (azulada en niños, amarillenta en
ancianos. Grosor: 0,5 (anterior) – 1 (posterior) mm.
Atravesada por nervio óptico, 4 venas vorticosas, vasos y nervios ciliares
anteriores. Es de conjuntivo denso con haces de colágeno aplanados en todas las
direcciones y algunas fibras elásticas.
316
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
3 capas fibrilares:
x Lámina epiescleral o epiesclerótica: externa, separada de la cápsula por el
espacio de Tenon. Conjuntivo laxo, rico en capilares.
x Sustancia propia o estroma: Avascular, con haces de colágeno gruesos
(conjuntivo denso) con ramificación serpenteante. Escasos fibroblastos y
sustancia fibrilar.
x Lámina fusca o supracoroidea: Está adyacente a la coroides, con haces de
colágenos pequeños y fibras elásticas. Hay células conjuntivas y melanocitos.
Sus funciones son la de protección y proveer puntos de fijación para la inserción
de músculos.
Limbo esclerocorneal
Zona de transición entre la esclerótica y la córnea. Hay un epitelio anterior que
da paso a un epitelio conjuntival bulbar al empezar el limbo. La membrana de Bowman
se interrumpe.
Contiene células madre del epitelio corneano y frena el avance del epitelio
conjuntical hacia la córnea. Se puede producir una conjuntivalización de la córnea, con
vascularización, aparición de caliciformes, epitelio irregular y disminución de la visión.
Además, contiene el aparato de drenaje del humor acuoso, para drenar existen
una red trabecular o espacios de Fontana, que son un conjunto de canales revestidos de
endotelio. Tras ello se encuentra el conducto de Schlemm, que circunda la córnea, y tras
ello las venas acuosas y el plexo espiescleral. El volumen de drenaje es igual al volumen
de secreción, por lo que la presión intraocular es constante.
El 85% del líquido alcanza el conducto de Schlemm. que circunda la córnea.
Hay venas acuosas que se dirigen hacia el plexo venoso epiescleral. El otro 15% se
dirige al flujo uveoescleral y se produce mediante percolación al tejido conjuntivo que
rodea a las fibras musculares del cuerpo ciliar
317
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Clínica
El glaucoma es una obstrucción del drenaje del humor vítreo con aumento de la
presión intraocular, lo que altera la nutrición y función de la retina. Se producen daños
en la retina, dolor, naúseas, visión borrosa, alteración en adaptación a la oscuridad,
halos en objetos luminosos y ceguera.
Córnea
1/6 anterior. Es transparente y el primer medio de difracción. Su grosor para de
0,5 (centro) a 1 (periferia) mm. Está cubierta por secreción lagrimal y es avascular.
10,5x11x5 mm
318
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Epitelio anterior o corneano: Estratificado plano no queratinizado (6 capas), con
microvellosidades que retinen la secreción lagrimal (si se reseca hay úlceras) en
las células superiores plasnas. Poseen queratinas K5/14 y K 3/12, para cuya
expresión se necesita vitamina A. Los núcleos tienen ferritina sobre ellos, que
protege frente a UV.
Hay uniones con hemidesmosomas, con una inervación rica gracias a
terminaciones nerviosas libres. Parten de ella aferencias del reflejo del parpadeo
(oclusión palpebral, secreción lagrimal, dolor).
x Membrana de Bowman: Lámina basal de 6-9 micras, homogénea. Es la parte
más anterior del estroma corneal, aunque se organiza de forma distinta.
Tiene MEC y fibrillas colágenas I, II, V y VI. No hay fibras elásticas, sino que
aporta resistencia. Se encarga del freno a infecciones y lesiones.
No se regenera, si se lesiona se producen cicatrices opacas, con lesiones
asociadas a erosiones córneas recidivantes.
x Estroma corneano o sustancia propia: 90% de grosor, con 60-70 láminas anchas
de fibrillas de colágeno I y V en haces paralelos. Entre los haces de cada lámina
hay MEC rica en glucosaminoglucanos sulfatados (condroitín y queratán
sulfato) y queratocitos (fibroblastos muy aplanados). Hay nervios mielínicos
hacia el epitelio anterior y es avascular.
La orientación de láminas es alterna, lo que aporta resistencia a tracción y
provoca que haya disposición pseudocristalina, aportando transparencia a la
córnea.
Tras su lesión puede haber infiltración de neutrófilos y linfocitos o una
alteración de la distribución ortogonal de las laminillas que conduce a la
opacidad de la córnea.
x Membrana de Descemet: Lámina basal, de 5-10 micras. Su grosor aumenta con
los años. Es una capa hialina, PAS+, con mallas hexagonales de colégeno IV y
VIII. Es producida por el epitelio posterior.
Se encarga de frenar infecciones y lesiones y sí se regenera. Se extiende bajo la
esclerótica (ligamento pectíneo o pectinado), por lo que al llegar al limbo no
desaparece
x Epitelio posterior, endotelio corneal o endotelio de la cámara anterior: Epitelio
plano simpe con uniones de sellado. Su membrana tiene transportadores de iones
y gran desarrollo de RER, Golgi y mitocondrias.
Es permeable al oxígeno, interviene en la ruta del glutatión para neutralizar el
exceso de oxígeno y metaboliza glucosa.
Se encarga de la difusión de nutrientes desde el humor acuoso y regula el
contenido de agua. Si el estroma se hidrata se produce una opacidad.
Tiene capacidad de proliferación limitada. Si se daña se realiza una
querastoplastia penetrante, con un 90% de éxito, gracias a la expresión de MHC
II, la expresión de inmunodepresores, la escasez de células de Langerhans y
CPA, la expresión del ligando Fas y carecer de vasos sanguíneos y linfáticos.
319
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Coroides
La coroides es una lámina de color pardo oscura insertada entre la esclerótica y
la retina. Por la parte anterior se continúa, a través de la ora serrata, con el cuerpo ciliar,
mientras que por la parte posterior termina en el nervio óptico, con el que está muy
unida, dado que sus conectivos son continuos. Por la parte externa, la coroides está
unida en forma laxa con la esclerótica, separadas por un espacio potencial, el espacio
pericoroideo. Por la parte interna, la coroides está unida al epitelio pigmentario de la
retina.
Lámina epicoroidea
El espacio supra o pericoroideo (entre coroides y esclerótica) se encuentra
atravesado por la conocida como lámina supracoroidea, epicoroidea o epicoroides. Al
igual que en la coroides, en la epicoroides encontramos abundantes vías linfáticas en
relación con los espacios linfáticos epicoroideos, así como vasos sanguíneos (conocidos
como vasos ciliares) y nervios. Esta lámina a su vez está constituida por abundantes
laminillas, en las que podemos hallar:
x Melanocitos: contribuyen a otorgar a la coroides su color característico
x Células linfoides: linfocitos, plasmocitos, macrófagos y células cebadas
x Fibroblastos y fibras colágenas y elásticas
x Miocitos
Capas de la coroides
x Capa vascular: es la más externa. Se denomina así porque contiene grandes
arterias y venas, que se encuentran rodeadas de un conjuntivo con fibras
elásticas y colágenas, miocitos y fibroblastos. Los vasos disminuyen de calibre a
medida que se acercan hacia la retina, y los mayores llegan incuso a introducirse
en el cuerpo ciliar. Su tamaño es tal que podemos observar con un
oftalmoscopio los vasos venosos, que tienen forma espiral y se conocen como
venas vorticosas.
x Capa coriocapilar: es la más interna, y su función es la nutrición de la retina.
Debe su nombre a la gran cantidad de capilares fenestrados que encontramos en
ella. A la altura de la fóvea, esta capa se engrosa, y la red capilar se hace más
densa.
Membrana de Brunch
Membrana amorfa delgada y retráctil de unas 3 micras de espesor que separa el
epitelio pigmentario de la capa coriocapilar. Se compone de:
x Lámina basal del endotelio de la capa coriocapilar.
x Capa de fibras colágenas.
x Red densa de fibras elásticas.
x Nueva capa de fibras colágenas.
320
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Membrana basal: pertenece al epitelio pigmentario, por lo que en realidad es
parte de la retina.
Clínica: drusas
Denominamos drusas a protuberancias en la membrana de Brunch debidas a la
acumulación de material amiloide. Cuando estas masas crecen, impiden el riego
adecuado de los fotorreceptores de la retina, con lo que inducen la degeneración del
epitelio pigmentario y de los fotorreceptores (recordemos que la encargada de la
nutrición es la capa coriocapilar, con lo que si la membrana de Brunch aumenta de
tamaño habrá una mayor separación entre esta y el epitelio pigmentario y se impedirá la
correcta alimentación del tejido).
La aparición de drusas constituye uno de los primeros indicios de degeneración
macular, lesión degeneraiva de la mácula lútea que cursa con pérdida de visión central.
Cuerpo ciliar
El cuerpo ciliar comprende la continuación anterior engrosada de la túnica
vascular, dado que se extiende como un anillo desde la ora serrata hasta la raíz del iris.
En un corte meridional, el cuerpo ciliar presenta una forma triangular: el vértice del
triángulo está orientado hacia atrás, en la ora serrata, y la base se encuentra dispuesta
hacia delante, hacia el iris.
Recordatorio: la ora serata es el ajustado límite anterior aserrado de la parte
sensorial de la retina, que se distingue con nitidez sobre la superficie interna del ojo
abierto después de retirar el cuerpo vítreo.
321
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Porción uveal
x Capa supraciliar: Continuación de la capa
vascular coroidea. Se trata de una zona de
transición entre el cuerpo ciliar y la
esclerótica cuya estructura laxa permite
cierto desplazamiento del primero frente a
la segunda, lo cual es importante para la
función del músculo ciliar en la
acomodación.
x Capilares fenestrados.
x Músculo ciliar: Permite la acomodación
del cristalino y, con ello, la transición entre
visión cercana y lejana, mediante un
mecanismo de disminución de la tensión
sobre la zónula de Zinn. Está constituido
por células musculares lisas rodeadas de
conectivo vascularizado. Todas las fibras
del músculo se insertan en un tendón
común, pero en tres direcciones distintas:
o Fibras longitudinales o de
Brucke: Son las más periféricas,
bordean la superficie escleral.
Se encargan de estirar la
coroides y abrir el canal de
Schlemm, que permite el
drenaje del humor acuoso.
o Fibras radiales: Se localizan en
la parte interna de las
longitudinales, y se orientan en
forma de abanico.
o Fibras circulares: Son las que
actúan modo de esfínter.
Porción neuroepitelial
Está constituido por un doble epitelio. No se trata de un epitelio estratificado,
sino de dos epitelios diferentes, cuyas capas apicales se encuentran enfrentadas, dejando
entre ellas el conducto ciliar:
x Epitelio pigmentado externo: Se encuentra en continuidad con el epitelio
pigmentado de la retina, y se apoya en una lámina basal que es continua con la
membrana de Brunch.
x Epitelio no pigmentado interno: Continuo con la retina sensitiva.
En ambos casos, las membranas de los polos basales se encuentran replegadas.
Podemos dividir esta porción neuroepitelial en:
322
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Pars plana: Es la región posterior. Presenta un doble epitelio cuya superficie es
lisa, con pigmentación oscura.
x Pars plica: Se compone de un doble epitelio con unas 70 crestas o pliegues
blancos de recorrido longitudinal, los procesos ciliares, separados por surcos
pigmentados oscuros. El epitelio de los procesos ciliares se encarga de:
1. Segregar el humor acuoso.
2. Segregar y anclar fibras zonulares (fibrilina), que constituirán el
ligamento suspensorio del cristalino.
3. Participa en la barrera hematoacuosa.
Producción de humor acuoso
El humor acuoso es un líquido claro como el agua y con el mismo índice de
refracción que ésta, que también contiene glucosa, proteínas, aminoácidos, ácido
ascórbico, Na+ y Cl-. Ocupa las cámaras anterior y posterior del ojo, dado que tras ser
secretado a la cámara posterior por el cuerpo ciliar filtra a la cámara anterior por la
pupila, desde donde atraviesa la malla esclerocorneana y es eliminado a través de la red
trabecular del espacio iridocorneal (drena por los espacios de Fontana y el conducto de
Schlemm).
Su función es mecánica (mantiene la forma del ojo) y de nutrición de las
estructuras avasculares del ojo.
Secretado en forma activa por el epitelio ciliar no pigmentado. Los capilares
fenestrados del conjuntivo subyacente son muy permeables, pero el paso de sustancias a
través del epitelio ciliar no es libre, dado que existe una barrera hematoacuosa cuya base
estructural es la presencia de uniones ocludens entre las superficies apicales de las
células. Es decir: las sustancias atraviesan libremente la pared capilar, el estroma
conjuntivo y las hendiduras del epitelio pigmentado, pero son retenidas a nivel de la
barrera hematoacuosa.
Por este motivo, es necesario un transporte activo para atravesar el epitelio no
pigmentado.
Iris
Representa la parte más anterior de la túnica vascular: es el límite anterior del
cuerpo ciliar. Tiene la forma de un delgado disco situado en el plano frontal, justo por
delante del cristalino. Se encuentra adherido a la esclerótica por detrás del limbo externo
(margen ciliar).
Separa las cámaras posterior y anterior del ojo (puerta para flujo del humor
acuso). El iris actúa como diafragma contráctil del ojo, dado que el diámetro de la
pupila varía para controlar la cantidad de luz que entra.
323
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Pupila
Es una mancha redonda negra en el centro del iris cuyo diámetro varía
constantemente para regular la cantidad de luz que ingresa en el ojo:
x El estrechamiento de la pupila o miosis disminuye el diámetro de la pupila hasta
alrededor de 1 mm, y se debe a la acción del músculo esfínter de la pupila, cuya
inervación es parasimpática y procede del III par.
x La dilatación de la pupila o midriasis puede causar un aumento máximo de la
pupila de unos 8 mm y se debe a la acción del músculo dilatador de la pupila,
cuya inervación es simpática y procede del ganglio cervical superior.
Superficie anterior del iris
La superficie anterior del iris es irregular, y presenta numerosas crestas y surcos.
Su origen es mesenquimal y está cubierta por la lámina marginal anterior. Dicha capa se
denominaba antes erróneamente endotelio del iris, y está constituida en su mayor parte
por fibroblastos y melanocitos:
x Fibroblastos: Forman una capa bastante continua de células aplanadas con
prolongaciones ramificadas que dejan huecos entre sí. Los huecos permiten la
comunicación directa entre la cámara anterior y el estroma, puesto que
posibilitan el paso de líquidos y partículas de hasta 200 micras.
x Melanocitos: Forman una capa aplanada paralela a la superficie del iris justo por
detrás de la capa de fibroblastos, y también poseen prolongaciones, similares a
las de los fibroblastos. De ellos depende el color del iris: a mayor número de
melanocitos, más oscuro será.
Estroma del iris
Se compone de tejido conjuntivo laxo que contiene vasos radiales que se adaptan
al diámetro pupilar. Así, cuando la pupila se dilata los vasos se acortan, y cuando se
contrae los vasos se alargan.
Músculo esfínter de la pupila
Cerca del margen pupilar se sitúa una banda circular de musculatura lisa cuyo
origen es distinto del de los demás: neuroectodérmico.
324
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Superficie neuroepitelial posterior
Epitelio anterior
La capa epitelial anterior de la parte posterior del iris está formada por una única
capa de células, cada una con una parte muscular basal y una porción apical epitelial:
x La parte muscular basal es de origen neuroectodérmico y se extiende en forma
radial hacia el interior del estroma del iris. El conjunto de las prolongaciones
basales forma el músculo dilatador de la pupila, de orientación radial. Este
músculo desaparece en la periferia.
x La parte epitelial está constituida por un epitelipo pigmentado en continuidad
con el epitelio ciliar pigmentado.
Epitelio posterior
Contiene melanina y se halla en continuidad con el epitelio ciliar no pigmentado.
Hacia el interior de la pupila, el epitelio anterior y el posterior convergen para formar el
borde negro pupilar.
Clínica
x Uveitis: Se trata de una inflamación de la úvea, que puede ser a nivel:
1. Anterior: Afecta al cuerpo ciliar y al iris. Es una complicación secundaria
de diversas enfermedades que provoca un enrojecimiento y dolor del ojo.
2. Posterior: Infección directa de la úvea.
x Melanoma: A este nivel existen una gran cantidad de melanocitos, con lo que no
es raro que aparezcan tumores de tipo melanoma, que se pueden extender
(metástasis).
x Albinismo: Del mismo modo que sucede con los melanocitos de la piel, el
albinismo afecta a los del iris, provocando una hipopigmentación que conduce a
una mala visión e incluso a ataxia cerebelosa.
325
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Retina o Túnica Nerviosa
Se encuentra adherida a coroides, estando su parte más interna en contacto con
el humor vítreo. Presenta dos regiones porque no toda ella es fotosensitiva:
x Región Fotosensorial: Se halla detrás de la ora serrata. Contiene conos y
bastones. Aquí se encuentra la papila o disco óptico, que es un punto ciego que
no contiene axones.
x Región No Sensorial: No presenta ni conos ni bastones.
Disco Óptico
Es el lugar en el que se reúnen todos los axones de las células ganglionares, y
desde donde se lleva la información hasta el SNC. Estos axones son de las células
ganglionares. En el centro de este disco hay una depresión pálida, denominada Copa
óptica.
Esa depresión se suele revisar ya que un aumento de su tamaño puede indicar
patología. En patologías como el glaucoma, la copa óptica es muy grande. Además de
axones, en esta zona encontramos vasos, arterias y venas.
326
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Fóvea Central
Es una región deprimida en el centro de la retina, a 2.5mm de la papila óptica.
Aquí no encontramos vasos, es completamente avascular.
En vez de tener la misma estructura de la retina, sólo encontramos dos tipos de
células: conos y células de Müller (gliales). Estos conos son especiales porque son
bastante más largos y estrechos y la luz incide sobre ellos directamente. Además, se
disponen oblicuos a la superficie.
La retina es una formación que supone la llegada indirecta de la luz al cono y al
bastón. Esto significa que, para llegar a estas células, la luz tiene que atravesar las diez
capas de la retina (primero la capa 10, luego la 9 y así sucesivamente).
Esta fóvea central es la zona de la retina donde se consigue máxima capacidad
visual, tanto en buena como en mala iluminación.
Mácula Lútea
Es la zona que rodea a la fóvea. Aquí sí encontramos vascularización (a través
de una red capilar). En fresco, esta zona de la retina es roja, no amarilla. Se denomina
lútea porque encontramos un pigmento: la xantofila. La xantofila se acumula en
neuronas ganglionares y bipolares.
Este pigmento se acumula debido a que hay una máxima concentración de
conos, los cuales absorben la luz de onda corta. Por ello es aquí donde tenemos la visión
central, visión aguda y la capacidad de distinguir los colores.
En el resto de la retina, que no es ni mácula ni fóvea, hay una gran cantidad de
bastones. Es la región fotosensible, la cual permite captar distintas intensidades
luminosas. Estos bastones son los encargados de que podamos ver con poca o con casi
sin luz. No permiten distinguir los colores, pero son los que detectan el movimiento en
la visión periférica.
Estructura Histológica de la Retina
A partir de la vesícula óptica se forman dos capas:
x Pars Pigmentosa: Es la más externa y está formada por epitelio pigmentario
adherido a la coroides.
x Pars Nervosa: Tiene una gran capacidad de proliferación, llegando a tener una
estructura de 9 capas en la que se pueden observar fotorreceptores, neuronas y
glía.
La retina tiene una organización invertida. Esto significa que la luz no va
directamente a conos y bastones, sino que primero tiene que atravesar la capa 10 que es
la más interna.
327
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
La retina posee dos barreras vasculares para que su medio interno se mantenga
en condiciones óptimas y esté protegido frente a la entrada de posibles tóxicos. Está
parcialmente vascularizada desde la membrana limitante interna por ramas de arterias y
venas radiales. Desde estos vasos penetran capilares a una cierta distancia formando
unos lechos capilares. Estos capilares muestran un epitelio muy sellado que forma la
barrera hematorretiniana. La barrera retiniana externa está formada por las uniones
estrechas entre las células del epitelio pigmentario. Ésta es atravesada por un flujo
bidireccional de iones y moléculas de bajo peso molecular ya que buena parte del
material degradado por el epitelio pigmentario (restos de los discos de conos y bastones)
sale hacia la capa coriocapilar.
Capas de la Retina
328
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Capa 1: Epitelio Pigmentario
Es un epitelio prismático simple, apoyado en la lámina basal de la coroides:
membrana de Bruch o vítrea. Este epitelio es más alto en la fóvea.
Es una zona muy activa y presenta células típicas epiteliales, unidas por muchos
tipos de uniones tanto de sellado como de adhesión (ZO, MA, GAP). El núcleo de las
células está relegado al tercio basal y su envuelta tiene invaginaciones y escotaduras.
Tiene gran desarrollo de orgánulos como lisosomas, Golgi, REL y gránulos de melanina
(por eso está pigmentado).
La membrana basal se apoya en la vítrea. No es una membrana rectilínea, sino
que está muy invaginada, presentando una gran cantidad de pliegues.
La membrana apical se diferencia formando microvellosidades, los cuales
atrapan y rodean el segmento externo de conos y bastones. Entre la membrana apical
prismática y la de este cono o bastón no hay mecanismo de unión, solo matriz
extracelular muy viscosa.
En el desprendimiento de retina, la capa 1 y las de 2-10 se separan ya que no hay
mecanismo que las mantenga unidas. Este desprendimiento se da por traumatismos,
tracción del vítreo… Se trata con cirugía. Cuando una persona fallece, también se
produce esa separación de la pars nervosa.
La luz atraviesa toda la retina y llega hasta la capa 1. Este epitelio tiene que
absorber toda la luz para que no vuelva para atrás y forme un fenómeno de reflexión que
altere la visión.
También sintetiza la membrana de Bruch. Entre las células de este epitelio hay a
muchos mecanismos de unión y estas uniones constituyen una barrera: la Barrera
Hematorretiniana Externa. Con ello aísla la pars nervosa de la sangre.
Esto no significa que no haya intercambio de productos; la mayoría de nutrientes
se obtienen gracias a la coroides, pero el transporte tiene que ser transcelular, no
paracelular. Además, este transporte es bidireccional. Aunque la retina necesita
nutrientes de la coroides, ella misma también produce catabolitos que luego salen del
epitelio.
Estas células del epitelio pigmentario fagocitan unos elementos llamados discos
membranosos (se fabrican en conos y bastones y se encuentran en sus segmentos
externos).
Las células pigmentarias fagoctian y degradan el disco y reciclan los elementos
reutilizables (como los pigmentos visuales). Además, la vitamina A también se
almacena en este epitelio.
329
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Capa 2: Capa de Conos y Bastones
Solo incluye una parte de los conos y bastones. Aquí encontramos los segmentos
externo e interno. Es en el segmento externo donde se encuentran los discos
membranosos.
Capa 3: Membrana Limitante Externa
No es una membrana como tal, sino que es una zona donde encontramos un
anillo de uniones de adhesión (ZA, MA) que unen las células de Müller con los conos y
bastones. Esto proporciona estabilidad y separa las capas 1 y 2 del resto de la retina.
Esta membrana es continua, pero en puntos concretos es perforada por los segmentos
internos y externos que tienen que llegar a la capa 2.
Capa 4: Capa Nuclear Externa
Contiene los núcleos de conos y bastones, no forma una única hilera, sino hasta
9 capas de núcleos. Los de los conos son mayores, más pálidos y a su alrededor hay más
citoplasma. Además, son más próximos al segmento externo.
En los bastones, y no en los conos, está el istmo (entre el segmento interno y la
zona del soma). Además, existe sólo un reborde citoplásmico perinuclear.
Capa 5: Capa Plexiforme Externa
Es la primera en la que encontramos sinapsis (sinapsis en cinta). La sinapsis se
da entre fotorreceptores y células bipolares. También aquí están las células horizontales
de Cajal. Así, los fotorreceptores, las células bipolares y las células horizontales de
Cajal forman una tríada (esto claramente es una broma macabra del destino).
Las células horizontales de Cajal modulan el impulso e integran señales de un
mismo territorio. Hay múltiples sinapsis, por eso se dice que realiza una integración.
Capa 6: Capa Nuclear o Granular Interna
Hay muchos núcleos: de las bipolares, de las horizontales, de las de Müller y de
las células amacrinas.
Las prolongaciones de las horizontales están en la capa 5. Las bipolares tiene el
axón en la 7 y las dendritas en la 5.
Así, las prolongaciones de las células de Müller rodean y dan sostén a todos los
elementos que tenemos aquí. Las horizontales mandan prolongaciones a la capa
plexiforme externa. Las bipolares mandan prolongaciones a la capa plexiforme externa
(dendritas) e interna (axón). Por su lado, las amacrinas mandan prolongaciones bien a la
capa plexiforme externa o a la interna. Por último, existen unas células bipolares enanas
que están en la fóvea central y en la mácula lútea
330
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Capa 7: Capa Plexiforme Interna
Hay sinapsis entre el axón de las bipolares y las dendritas de las ganglionares. La
célula moduladora es la amacrina (también las de Cajal). Se distinguen diferentes tipos
de sinapsis, incluida la sinapsis en cinta.
Capa 8: Capa Ganglionar
Es muy importante, pues contiene células ganglionares. Es de mayor calibre. Los
núcleos están alienados y forman una única hilera, excepto en la mácula lútea donde hay
varios estratos. El nucléolo está bien marcado y los orgánulos bien desarrollados (RE),
llegándose a ver grumos de Nissl.
También hay dendritas que van a la capa plexiforme interna (donde se
ramifican) y un axón descendente que sale de la capa 8 y después se integra con otros
formando el nervio óptico. Los nervios forman una única hilera excepto a nivel de la
mácula lútea donde pueden verse hasta 8 hileras.
Capa 9: Capa de Fibras Nerviosas
Presenta axones de neuronas ganglionares que son amielínicos. Estos se van
aplanando y convergen en el nervio óptico. Estas células forman un estrato o capa plana,
en el que las células van paralelas a la superficie y donde al sumarse axones aumenta de
grosor, creciendo al acercarnos al disco óptico.
Capa 10: Membrana Limitante Interna
Es lisa, está en contacto con humor vítreo y está formada por los extremos más
ensanchados de las prolongaciones de las células gliales (de Müller y astroglía)
aplanadas y unidas. Asociados a esta membrana encontramos una lámina basal.
Vascularización de la Retina
La retina es un órgano que requiere del mantenimiento de la homeostasis. Para
ello contribuyen dos barreras: barreras hematorretiniana externa e interna.
Entre ambas aíslan la retina para protegerla de productos tóxicos y patógenos
que pueden llegar a través de la sangre. Las arterias y venas penetran por la cara interna
y dentro de la retina encontramos algunos lechos capilares continuos que no están en
cualquier capa sino únicamente en las más internas (6, 7 ,8 ,9 y 10) lo que se conoce
como zona vascular de la retina.
La capa interna en el endotelio de los capilares asegurando el control del paso de
moléculas de manera controlada, nada libre.
La externa se sitúa en la capa 1 en las uniones que sellan el espacio entre las
células en el epitelio pigmentario controlando también el paso de moléculas (va por
cuentagotas).
331
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Tipos de Células
Células de Müller
Son células gliales consideradas una variedad de astrocitos que forman columnas
desde la capa 2 a la 10, ocupando todo el espacio. Delimitan espacios, dan sostén,
rodean los somas de las neuronas y forman membranas limitantes (como los astrocitos).
Donde no haya ni prolongaciones ni núcleos de células, encontraremos este tipo de
células.
Es decir, aíslan todas las capas (por lo que protegen) y, además, proporcionan
una barrera externa y una interna con sus prolongaciones. Sus núcleos son aplanados, el
citoplasma es bastante basófilo y dan un soporte metabólico (nutricional) para las
neuronas de las capas internas pasando los nutrientes sobre todo donde no hay capilares
(capas 1 a 5). Sus prolongaciones se asocian a vasos en la porción vascular de la retina y
tienen escaso glucógeno.
Las células de la capa externa pueden encontrar nutrientes por sí mismas,
mientras que las de la parte interna, no.
Fotorreceptores (Bastones y Conos)
Hay muchos más bastones que conos. Tienen una longitud más larga los
bastones, aproximadamente el doble. Es así en toda la retina excepto en la fóvea, donde
los conos son tremendamente largos.
x Los bastones están muy distribuidos. Son muy sensibles a la luz (vemos en
penumbra). Esta visión se llama visión escotópica. Los bastones dan una imagen
en tonos grises, es decir, una visión grosera que no proporciona detalles
La degeneración de los bastones se denomina ceguera nocturna. Esto es un signo
de inicio de retinitis pigmentaria, la cual acaba llevando a la ceguera. Los genes
asociados a la retinitis se encuentran en el cromosoma X y 3.
x Los conos se concentran en la fóvea principalmente. Proporcionan una visión
muy detallada, pero para ello es necesario mucha luz. Por ello funcionan
principalmente de día y dan una visión fotópica.
De día tenemos una mayor agudeza visual y esta se debe a los conos. Los conos
proporcionan una visión en colores. Esto es posible ya que en estos fotorreceptores
hallamos unos pigmentos capaces de absorber la luz. Tenemos 3 pigmentos distintos
según la longitud de onda absorbida: hay pigmentos sensibles al azul (420nm), al verde
(535nm) y al amarillo y rojo (565nm), lo que nos da la imagen en colores. Los genes
que codifican para los fotopigmentos que permiten captar la onda verde y amarilla están
bastante próximos en el cromosoma X.
Así, cuando hay mutaciones de estos genes producen cegueras para los tres
colores, dos o uno de ellos y cuando hay mutaciones en el cromosoma X aparecen
332
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
problemas de distinción para el verde y amarillo (es decir, que es frecuente que, si no se
distingue el verde, no se distinga tampoco el amarillo).
333
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Los conos y los bastones son células polarizadas de forma que encontramos un
segmento externo, cuello, segmento interno (elipsoide y mioide), istmo, soma y por
último el axón (esférula y pedículo).
Segmento Externo
Está en la capa 2. Es cilíndrico en el bastón y cónico en el cono. Está en contacto
con las microvellosidades de las células del epitelio pigmentario, abrazándolas. Entre
medias hay una matriz extracelular rica en glucosaminoglucanos y proteínas en la que
destaca la IRBP (proteína intersticial ligadora de retinoides), que lleva el retinal al
epitelio pigmentario y de ahí al fotorreceptor correspondiente (y viceversa).
El segmento externo es la región fotosensible debido a que aquí es donde se
encuentra el disco horizontal. Estos discos son vesículas aplanadas que están
constantemente formándose y eliminándose.
Discos Horizontales
Los discos horizontales ascienden por el extremo apical y al final son
fagocitados por el epitelio pigmentario. Presentan una vida máxima de 10 días.
Estos discos se forman por una invaginación de la membrana plasmática del
segmento externo en la zona próxima al cuello (parte interna). Esta invaginación
334
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
profundiza y en un momento dado se separa de la membrana quedando como un disco
de membrana independiente.
Según se forman van empujando hacia la capa 1 a los que estaban ya ahí,
desplazándose a la zona más externa del segmento y van quedándose apilados.
Finalmente, a los 10 días son fagocitados por las células del epitelio pigmentario. No se
sabe cuándo ocurre en los conos esta fagocitosis pero en los bastones ocurre por las
mañanas cuando abrimos los ojos (por eso nos molesta tanto abrir los ojos por la
mañana).
Contienen una sola membrana en la que están los pigmentos visuales. En el caso
de los bastones es la rodopsina (o púrpura visual) y en el caso de los conos es la
yodopsina.
En los bastones la rodopsina se sintetiza y se incorpora inmediatamente a la
membrana del disco según el disco se está formado, es decir, nada más producirse la
invaginación de la membrana. La yodopsina, en cambio, se puede incorporar a la
membrana del disco en cualquier momento y zona del segmento externo (en zonas más
alejadas también).
Cada pigmento está formado por dos moléculas distintas, un cromóforo que es el
retinal, formado a partir de vitamina A y una opsina (sensible a una longitud de onda
concreta) que sería la escotopsina en los bastones y la fotopsina en los conos.
Cuello o Pedículo de Fijación
Es la región estrecha entre segmentos externo e interno. Fija el extremo externo
al interno. En su interior se observa la presencia de un cilio modificado en el que faltan
los microtúbulos centrales (9+0) y hay que tener en cuenta en toda molécula que vaya al
segmento externo (ya que en él no se sintetizan) tiene que pasar por aquí por lo que esta
región registra mucho tráfico de moléculas. A esta altura también aparecen las
prolongaciones caliciales, que son prolongaciones cortas que van desde el extremo
distal del segmento interno hasta el proximal del segmento externo, pero no contactan
con él.
Segmento Interno
Es más grueso en los bastones que en los conos y se divide en dos porciones:
x Elipsoide: Es la más externa y eosinófila. En ella tenemos una mayor cantidad
de mitocondrias (que aportan energía para transporte) y un mayor desarrollo del
citoesqueleto, sobre todo microtúbulos.
x Mioide: Es la zona más interna y es donde se encuentran los elementos
relacionados con la síntesis de proteínas y lípidos (hay abundante REL,
ribosomas, Golgi y glucógeno). También tiene microtúbulos. El segmento
interno es la parte de la célula en la que se desarrolla todo el metabolismo para la
actividad del fotorreceptor, por lo que contiene la maquinaria metabólica de la
335
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
célula. Su base y por tanto la base del segmento interno se encuentra perforando
la capa 3 (membrana limitante externa).
El soma de los fotorreceptores se encuentra en la capa 4 (granular externa). El de
los bastones el soma se une al segmento interno por el istmo y tiene un reborde
citoplásmico estrecho, mientras que en los conos no hay istmo, los núcleos son mayores
y más pálidos y el reborde es más amplio.
Desde la capa 4 parte una prolongación estrecha que se mete en la capa 5 y se
denomina fibra interna. Este axón/fibra va a la plexiforme externa, donde sinapta. El
extremo distal de este axón es distinto en cono y bastón. En el caso del bastón, se forma
una estructura llamad esférula y en el cono se forma un pedículo.
La sinapsis en la capa plexiforme externa se forma con la dendrita de una
neurona bipolar y el axón de un cono o bastón.
Cada uno de los conos o bastones sinapta con múltiples núcleos bipolares.
Existen unos cuantos receptores que sinaptan con una bipolar. Esto ocurre con los
fotorreceptores que sinaptan en la fóvea.
Las esférulas son más pequeñas, ovaladas y rodean a los extremos de las
dendritas bipolares y horizontales, formando multisinapsis. En cambio, los pedículos
son terminales axónicos grandes con perfil piramidal y forman tríadas (con tres
elementos), sinapsis sencillas y contactos interreceptores. Entre medias, tenemos las
horizontales con papel integrador y en la fóvea cada cono realiza sinapsis con una sola
neurona bipolar.
336
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Histofisiología
La transducción consiste en la captación de estímulos luminosos y
transformación en impulsos nerviosos.
337
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
En oscuridad los fotorreceptores están despolarizados y los canales para Na+
dependientes de voltaje abiertos, lo que deermina la liberación de un neurotransmisor
inhibidor.
Cuando hay luz en el bastón el cromófero está en una fase de 11 cis-retinal y
reacciona con el fotón. Se produce una isomerización y pasa a 11 trans-retinal. La
molécula pasa de denominarse rodopsina a luminodopsina. La opsina, una proteína G,
actúa sobre la transducción y entra en acción la GMPc fosfodiesterasa, por lo que
disminuye el GMPc. El cierre de canales dependientes de voltaje conlleva una
hiperpolarización y se impide de este modo la liberación de neurotransmisores
inhibidores, lo que estimula la transmisión de estímulos.
Cuando volvemos a la oscuridad la luminodopsina se degrada en opsina y 11
trans-retinal. Los ojos vuelven a su color blanco tras estar rojos puesto que los
productos son incoloros, esto se conoce como blanqueamiento. Si se extrae la retina de
un animal mantenido en la oscuridad se observará roja púrpura, pero se aclarará
rápidamente al exponerla a la luz.
El 11 trans-retinal pasa a la pars pigmentosa vía IRPB. La rerinolisomerasa del
interior convierte en 11 cis-retinal. El 11 cis-retinal vuelve al bastón vía IRPB. Se
combina con opsina para generar rodopsina.
Mientras que estamos en la oscuridad el bastón sintetiza GMPc y se normaliza la
permeabilidad de la membrana, abriéndose canales de sodio.
La energía es aportada por el segmento interno. La reconversión del trans-retinal
en cis implica a las células de Müller y al epitelio pigmentario.
338
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Células bipolares
Las dendritas sinaptan con esférulas de bastones o pedículos de conos (en la
capa plexiforme externa) y los axones con ganglionares (en la capa plexiforme interna).
Las células bipolares de los conos pueden ser directas (1pedículo-1bipolar-1ganglionar)
o difusas (n pedículos-1bipolar-n ganglionares).
Las células bipolares del cono directas establecen sinapsis con un solo pedículo
conal y un axón único que contacta con una sola célula ganglionar. En esencia, las
células bipolares directas unen un solo cono con una fibra del nervio óptico, mientras
que las difusas tienen unas vías aferentes y eferentes más amplias. Los núcleos de las
células bipolares forman parte de la capa de los núcleos interna de la retina.
Células ganglionares
Sus dendritas sinaptan con las bipolares (capa plexiforme interna) y el axón
forma parte del nervio óptico. Existen dos tipos de células ganglionares: las células
ganglionares difusas, que contactan con varias células bipolares, y las células
ganglionares directas, cuyas dendritas contactan con una sola célula bipolar directa.
Obsérvese que las células ganglionares directas sólo reciben impulsos de los conos.
Vía directa por bipolares enanas que contactan por un único cono y pasa, por
tanto, información a una única bipolar. Son específicas para cada cono y están en gran
cantidad en la mácula lútea.
Células horizontales y amacrinas
Las células horizontales y amacrinas no tienen axones ni dendritas, sólo
prolongaciones neuríticas que transmiten en ambas direcciones. Los núcleos de las
células horizontales y amacrinas contribuyen a la capa de los núcleos interna.
Las células horizontales tienen múltiples prolongaciones que sinaptan con varias
esférulas y pedículos (5): Integran conos y bastones de áreas de la retina adyacentes.
En las células amacrinas se distigue una única prolongación que se ramifica y
sinapta con axones de las bipolares y dendritas ganglionares (7). Las sinapsis son
recíprocas y en cinta. Muestrea y modula el estímulo que sale de las bipolares hacia las
ganglionares.
339
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Vías ópticas
Son las diferentes rutas que permiten llevar la información captada por los
fotorreceptores hasta el SNC. De ello se va a encargar el nervio óptico, una estructura
par. Las fibras nerviosas de este medio son:
x Fibras homolaterales: Se sitúan laterales y van a permitir la formación de la
imagen.
x Fibras heterolaterales: Se localizan mediales. Se ocupan de acomodar la lente y
de regular los movimientos de los músculos de la órbita ocular.
La información parte de los fotorreceptores vía células bipolares hasta las células
ganglionares, cuyos axones forman el nervio óptico. Estos axones van a llegar al
diencéfalo, en concreto hasta los cuerpos geniculados laterales, y de aquí ascienden al
telencéfalo alcanzando la corteza occipital.
Por lo que se refiere la acomodación de la lente las fibras llegan hasta el
mesencéfalo, donde las fibras sinaptan en la región pretectal y en el núcleo de Edinger-
Westphall, que contiene neuronas parasimpáticas que se integran en el tercer par desde
donde alcanzan el ganglio ciliar. Estas fibras van a inervar el iris y el cuerpo ciliar.
Los movimientos oculares, del globo ocular, son controlados por distintos
músculos que están inervados por neuronas del mesencéfalo, concretamente en el
tubérculo cuadrigémino posterior. Inervan los músculos del glóbulo ocular permitiendo
que gire dentro de la órbita.
Cristalino
Es una lente biconvexa elástica situada delante del humor vítreo y detrás del iris
y del humor acuoso. Con la vejez pierde la elasticidad y pierde la capacidad para
enfocar sobre la retina. Es una estructura poralizada, siendo el polo anterior más inferior
y el posterior más superior.
Se trata de una estructura elástica que permite enfocar la visión y acomodar la
lente que es el ojo humano. Es transparente, lo que facilita la visión de los objetos. Es
avascular, por lo que su nutrición es llevada a cabo por difusión de los humores acuosos
y vítreo.
El cristalino está suspendido en el ojo por el cuerpo ciliar, al que se conecta por
medio de unas fibras denominadas zónula de Zinn o ligamento suspensorio.
Está formado por millones de fibras transparentes organizadas de manera muy
regular y todas mediante. Está compuesto por una cápsula, un epitelio subcapsular con
células cúbicas y las fibras del cristalino.
340
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Cápsula
Lámina basal gruesa producida por células del epitelio anterior con colágeno IV,
I y II en menor medida. Es laminillar y más gruesa en la zona ecuatorial, donde se
inserta la zónula de Zinn.
Por debajo de su parte anterior se encuentra una sola capa de células epiteliales
cúbicas, que se extienden en sentido posterior hasta la región ecuatorial. No se
encuentran células epiteliales por debajo de la superficie posterior de la cápsula.
Epitelio subcapsular
Está en la superficie anterior con células cúbicas: A y E (mayor índice mitótico).
Los núcleos son centrales redondos y poseen zónulas occludens. Se desplazan y tienen
una elevada tasa de división mitótica.
En región ecuatorial, se alargan y rotan para quedar paralelas a la superficie, a la
vez que sintetizan faquinina, filensina y cristalinas de tipo alfa, beta y gamma. Tienen
metabolismo productor de glucosa, si la glucosa aumenta se acumula sorbitol. El exceso
de sorbitol lleva a la insolubilización de proteínas y a la pérdida de transparencia Æ
cataratas (más adelante desarrollado).
Al llegar nuevas células, las anteriores se desplazan al centro (región de fibras).
Fibras del cristalino
Las células del epitelio subcapsular aumentan de altura y pierden el núcleo y los
orgánulos. Se llenan de proteínas cristalinas y migran hacia posterior. En la zona central
están comprimidas y no se distinguen unas con otras.
Las cristalinas elevan el índice de refracción y persisten toda la vida. Permiten
de esta forma un crecimiento lento y continuo del cristalino. En la época senil las fibras
están retraídas y condensadas.
341
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Refracción
Un dioptrio es la superficie de separación de dos medios en los cuales la
velocidad de la luz no es la misma.
Podemos asumir los frentes de onda luminosas a rayos, que experimentan dos
fenómenos básicos al incidir en un dioptrio: reflexión y refracción. Parte de la onda
vuelve al medio 1. Se rige por la ley de la reflexión. Otra parte de la onda atraviesa el
dioptrio, pero cambia su dirección; se rige por la ley de la refracción (ley de Snell).
El ojo en situación normal enfoca al infinito y la imagen se forma en la retina,
Su potencia es de 56 dioptrias., con una capacidad de acomodación de 4 dioptrías.
Alteraciones: presbicia, miopía e hipermetropía.
342
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Cataratas
Se llama catarata a cualquier opacidad del cristalino, focal, difusa, pequeña o
grande. El factor etiológico más común es la edad, explicado por el aumento progresivo
de la densidad cortical del cristalino por la compactación de las fibras epiteliales.
Existen cataratas secundarias por diversos motivos (muchísmos): causas físicas,
infecciones, metabólicas, inflamatorias...
Cambios bioquímicos:
x Aumento inicial en el contenido de agua.
x Disminución del agua en la medida en que la catarata madura.
x El contenido de potasio disminuye progresivamente.
x El sodio y el cloro aumentan.
x Disminuye el consumo de oxígeno y se reduce la concentración de glutatión y
ácido ascórbico.
x Disminuye la cantidad de proteínas
x Oxidación de proteínas cristalinas.
x Depósitos proteicos de alto peso molecular.
Variación de las proteínas del cristalino por desnaturalización, dispersión
elevada de la luz por la filensina y cristalinas agregadas. Esto afecta a la visión correcta.
La cirugía es la única opción terapéutica. Se quita todo el cristalino y es solo
preservado el saco capsular, dentro del cual se implanta una lente con poder de
convergencia previamente calculado para lograr un resultado refractivo postoperatorio
lo más cercano posible a la emetropía. Complicaciones: rotura de la cápsula posterior Æ
glaucoma y desprendimiento de retina.
Humor vítreo
Sustancia transparente gelatinosa que se sitúa en el espacio existente entre
cristalino y retina. Aproximadamente un 99% de su composición es agua, y en total
hallamos unos 4 mL de líquido en la cámara vítrea.
Es considerado un tejido de sostén
especializado con un mayor contenido en agua
de lo normal: de ahí que en él encontremos
células conjuntivas denominadas hialocitos, que
producen una MEC rica en fibras colágenas y
ácido hialurónico (análogos a los fibroblastos y a
la MEC de un conjuntivo común). A este nivel
en el feto existe una arteria, denominada arteria
hialoidea, que sufre atrofia, quedando su residuo
en el adulto como conducto hialoideo o de
Cloquet.
343
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
ANEJOS AL GLOBO OCULAR
Conjuntiva
Es una membrana translúcida que tapiza la superficie interna de los párpados y
la superficie anterior del globo hasta el limbo esclerocorneal, lo cual nos permite
distinguir en ella dos caras:
x Bulbar: la que tapiza el globo ocular
x Palpebral: tapiza la superficie interna del párpado.
Se trata de una mucosa, compuesta por un epitelio y una lámina propia.
x Epitelio: es diferente en ambas caras:
o Cara palpebral: epitelio estratificado prismático con células caliciformes
o Cara bulbar: epitelio estratificado cúbico a plano no queratinizado (epitelio
corneal anterior).
x Lámina propia: es un tejido conjuntivo laxo rico en adipocitos, linfocitos y vasos
sanguíneos y linfáticos, cuyo contenido en fibras colágenas disminuye con la
edad.
Clínica
x Pinguécula: Es un engrosamiento de la conjuntiva bulbar provocado por una
exposición continua de la misma al polvo, al sol o al aire, que desencadena una
proliferación del estroma.
x Hemorragia subconjuntival: Al contrario de lo que podría parecer, no altera la
visión. Se puede producir por diversas causas (traumatismo, hipertensión), pero
la más frecuente es la conjuntivitis, que provoca una dilatación de los vasos
superficiales, asociada a edema conjuntival con secreción que puede ser:
o Acuosa: si la conjuntivitis es de origen viral
o Purulenta: si es bacteriana
El tratamiento de una hemorragia conjuntival se realiza con anticoagulantes.
Párpados
Son dobles pliegues cutáneos capaces de abrirse y cerrarse presentes en la parte
anterior del globo ocular. Protegen córnea y esclerótica y ayudan a regular la cantidad
de luz que entra al ojo.
Antes de entrar en sus capas, cabe destacar que en los márgenes palpebrales
encontramos dos o tres hileras de pestañas. Las pestañas son pelos cortos, rígidos y
curvos, con una intensa pigmentación, de vida corta. Proceden de folículos pilosos
cortos y rectos en los que no se inserta ningún músculo piloerector.
344
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Piel
A este nivel no existen papilas
dérmicas, pero sí vello, aunque es incoloro.
Capa subcutánea
Es tejido conjuntivo laxo.
Músculo orbicular
Es un músculo esquelético que regula
el cierre del párpado, inervado por el nervio
facial (VII).
Placa tarsal
Es el “esqueleto” del párpado. No está
constituido por hueso obviamente, sino por
tejido conjuntivo denso rico en fibras
colágenas y elásticas. Tiene dos porciones:
x Inferior: Con el músculo oblicuo menor.
x Superior: Con el músculo palpebral superior, inervado por el nervio oculomotor
(III) y encargado de echar el párpado hacia atrás. La parálisis de este músculo
provoca ptosis palpebral.
Conjuntiva palpebral
Ya hemos hablado de ella.
Glándulas palpebrales
Se encuentran inervadas por las dos divisiones del SNA:
x SNPS: Las fibras parasimpáticas proceden del ganglio pterigopalatino.
x SNS: Las fibras simpáticas provienen del ganglio cervical superior.
La secreción de estas glándulas se encuentra sincronizada con la de la glándula
lacrimal principal por medio del neurotransmisor VIP.
De Meibomio
Son glándulas sebáceas tubuloacinares largas, que podemos observar en los
tarsos como estrías amarillentas verticales (25 en el párpado superior y 20 en el
inferior). No comunican con ningún folículo piloso.
Su secreción es de carácter oleoso (ya hemos dicho que son sebáceas) y se
extiende sobre la secreción lacrimal, retrasando su evaporación. La principal función de
esta secreción es impedir la unión de los párpados.
345
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
DeZeiss
Son glándulas sebáceas pequeñas
que sí se comunican con los folículos
pilosos, concretamente con los de las
pestañas.
De Moll
Son glándulas sudoríparas
apocrinas con sinuosos conductos
excretores que se encuentran en el margen
palpebral.
Glándulas lacrimales accesorias
x De Krause: en el fórnix de la conjuntiva.
x De Wolfring: sobre la placa tarsal superior.
Aparato lagrimal
Está compuesto por la glándula lacrimal principal y las dos glándulas lacrimales
accesorias que se acaban de mencionar.
Glándula lagrimal
Es una glándula bilobulada que se encuentra en el margen superoexterno de la
órbita o margen superoexterno del párpado superior, por debajo de la conjuntiva.
Sus adenómeros son tubuloalveolares, y tienen una capa de células
mioepiteliales que ayudan a expulsar la secreción. Su luz es grande y drenan por medio
de entre 6 y 12 conductos excretores.
346
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Secreción lagrimal
Es muy heterogénea, puesto que es una mezcla de las secreciones de:
x Glándula lagrimal principal.
x Glándulas lagrimales accesorias.
x Células caliciformes de la conjuntiva bulbar.
x Glándulas tarsales palpebrales.
Se trata de una secreción serosa y alcalina, cuyos componentes son:
x Proteínas, lípidos, metabolitos, electrolitos, fármacos.
x Lisozima (bactericida).
x Lactoferrina (bacteriostático).
x sIgA.
Tiene como función barrer cuerpos extraños y humedecer y lubrificar la
superficie del globo ocular y cara interna de los párpados.
La secreción lagrimal drena por los conductos lagrimales superior e inferior, que
se hallan en el ángulo interno del ojo y están revestidos por epitelio estratificado plano.
De ahí pasa al conducto lagrimal común, que lo lleva al saco lagrimal (a partir del cual,
este incluido, el sistema se encuentra tapizado por epitelio respiratorio), que drena a la
cavidad nasal a través del conducto lacrimonasal.
Clínica
x Lagrimeo excesivo (no es clínica a no ser que sea extremo): Puede ser
desencadenado por irritantes físicos y químicos, por exposición a una luz intensa
o por emociones intensas.
x Ojo seco o queratoconjuntivitis seca: Desecación de la córnea por alteración en
la producción de lágrimas. Puede provocar úlceras, perforación, pérdida del
humor acuoso y finalmente ceguera.
347
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Cristalino
Es una lente biconvexa elástica situada delante del humor vítreo y detrás del iris
y del humor acuoso. Con la vejez pierde la elasticidad y pierde la capacidad para
enfocar sobre la retina. Es una estructura poralizada, siendo el polo anterior más inferior
y el posterior más superior.
Se trata de una estructura elástica que permite enfocar la visión y acomodar la
lente que es el ojo humano. Es transparente, lo que facilita la visión de los objetos. Es
avascular, por lo que su nutrición es llevada a cabo por difusión de los humores acuosos
y vítreo.
El cristalino está suspendido en el ojo por el cuerpo ciliar, al que se conecta por
medio de unas fibras denominadas zónula de Zinn o ligamento suspensorio.
Está formado por millones de fibras transparentes organizadas de manera muy
regular y todas mediante. Está compuesto por una cápsula, un epitelio subcapsular con
células cúbicas y las fibras del cristalino.
Cápsula
Lámina basal gruesa producida por células del epitelio anterior con colágeno IV,
I y II en menor medida. Es laminillar y más gruesa en la zona ecuatorial, donde se
inserta la zónula de Zinn.
Por debajo de su parte anterior se encuentra una sola capa de células epiteliales
cúbicas, que se extienden en sentido posterior hasta la región ecuatorial. No se
encuentran células epiteliales por debajo de la superficie posterior de la cápsula.
Epitelio subcapsular
Está en la superficie anterior con células cúbicas: A y E (mayor índice mitótico).
Los núcleos son centrales redondos y poseen zónulas occludens. Se desplazan y tienen
una elevada tasa de división mitótica.
En región ecuatorial, se alargan y rotan para quedar paralelas a la superficie, a la
vez que sintetizan filensina y cristalinas. Tienen metabolismo productor de glucosa, si la
glucosa aumenta se acumula sorbitol. El exceso de sorbitol lleva a la insolubilización de
proteínas y a la pérdida de transparencia Æ cataratas (más adelante desarrollado).
Al llegar nuevas células, las anteriores se desplazan al centro (región de fibras).
Fibras del cristalino
Las células del epitelio subcapsular aumentan de altura y pierden el núcleo y los
orgánulos. Se llenan de proteínas cristalinas y migran hacia posterior. En la zona central
están comprimidas y no se distinguen unas con otras.
348
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Las cristalinas elevan el índice de refracción y persisten toda la vida. Permiten
de esta forma un crecimiento lento y continuo del cristalino. En la época senil las fibras
están retraídas y condensadas.
Refracción
Un dioptrio es la superficie de separación de dos medios en los cuales la
velocidad de la luz no es la misma.
Podemos asumir los frentes de onda luminosas a rayos, que experimentan dos
fenómenos básicos al incidir en un dioptrio: reflexión y refracción. Parte de la onda
vuelve al medio 1. Se rige por la ley de la reflexión. Otra parte de la onda atraviesa el
dioptrio, pero cambia su dirección; se rige por la ley de la refracción (ley de Snell).
349
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
El ojo en situación normal enfoca al infinito y la imagen se forma en la retina,
Su potencia es de 56 dioptrias., con una capacidad de acomodación de 4 dioptrías.
Alteraciones: presbicia, miopía e hipermetropía.
Cataratas
Se llama catarata a cualquier opacidad del cristalino, focal, difusa, pequeña o
grande. El factor etiológico más común es la edad, explicado por el aumento progresivo
de la densidad cortical del cristalino por la compactación de las fibras epiteliales.
Existen cataratas secundarias por diversos motivos (muchísmos): causas físicas,
infecciones, metabólicas, inflamatorias...
Cambios bioquímicos:
x Aumento inicial en el contenido de agua.
x Disminución del agua en la medida en que la catarata madura.
x El contenido de potasio disminuye progresivamente.
x El sodio y el cloro aumentan.
x Disminuye el consumo de oxígeno y se reduce la concentración de glutatión y
ácido ascórbico.
x Disminuye la cantidad de proteínas
x Oxidación de proteínas cristalinas.
x Depósitos proteicos de alto peso molecular.
Variación de las proteínas del cristalino por desnaturalización, dispersión
elevada de la luz por la filensina y cristalinas agreagadas. Esto afecta a la visión
correcta.
La cirugía es la única opción terapéutica. Se quita todo el cristalino y es solo
preservado el saco capsular, dentro del cual se implanta una lente con poder de
convergencia previamente calculado para lograr un resultado refractivo postoperatorio
lo más cercano posible a la emetropía. Complicaciones: rotura de la cápsula posterior Æ
glaucoma y desprendimiento de retina.
350
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
SISTEMA NERVIOSO
GENERALIDADES
El sistema nervioso está compuesto por:
x SN Central: Forma un eje de simetría bilateral. Está constituido por el encéfalo y
la médula espinal, revestidos por una caja ósea.
x SN Periférico: Conecta el eje de simetría bilateral que constituye el SNC con la
periferia del organismo. Está formado por un sistema plexiforme de ganglios y
nervios.
Además, de manera independiente, se puede realizar otra división, con un
criterio funcional y no anatómico (ambos van a estar englobados a la vez en SNC y
SNP):
x SN Autónomo o vegetativo: Sistema motor encargado de inervar las vísceras.
x SN de la vida de relación: Sistema que en esencia regula las funciones no
viscerales. Permite la interacción con el medio ambiente y se encarga de la
percepción consciente, el procesamiento de la información y la realización de
movimientos automatizados y voluntarios.
Recordemos que la unidad básica del tejido nervioso es la neurona, que se
intercala entre las divisiones sensitivas y musculares formando redes con múltiples
sinapsis. Su finalidad es procesar la información y transmitirla.
SN CENTRAL (SNC)
Forma un eje de simetría bilateral. Está constituido por el encéfalo y la médula
espinal, estructuras que se encuentran altamente protegidas, tanto por una caja ósea
(cráneo y columna vertebral) como por un almohadillado de líquido cefalorraquídeo y
distintas cubiertas protectoras. En él vamos a encontrar una barrera denominada barrera
hematoencefálica, de la que hablaremos más adelante.
Tiene una serie de cavidades, por lo que podemos decir que es hueco, y es
epineural.
Desarrollo
El sistema nervioso en su conjunto procede del neuroectodermo. Su formación
comienza con el proceso de neurulación: aparece en el feto un tubo hueco, denominado
tubo neural, sobre el que se distinguen irregularidades conocidas como crestas neurales.
El conjunto de tubo neural y crestas neurales constituyen la placa neural, darán lugar a:
351
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Crestas neurales: formarán los ganglios nerviosos, los melanocitos (recordad que
son de origen neuroectodérmico) y la médula adrenal.
x Tubo neural: originará el SNC, es decir, que es propiamente el que dará lugar al
encéfalo y la médula espinal. La porción anterior del tubo neural se dilata e
invagina, y cuando esta invaginación de independiza se forma la vesícula
encefálica, la cual a su vez se dividirá formando tres vesículas primarias:
o Anterior: dará lugar al prosencéfalo.
o Media: formará el mesencéfalo.
o Posterior: origen del rombencéfalo.
Estas darán lugar a 5 vesículas secundarias, que formarán:
o Telencéfalo Æ futura corteza cerebral.
o Diencéfalo Æ tálamo, epitálamo e hipotálamo.
o Mesencéfalo Æ tronco cerebral.
o Metencéfalo Æ protuberancia, cerebelo y pedúnculos cerebelosos.
o Mielencéfalo Æ bulbo raquídeo.
La maduración de las distintas estructuras nerviosas no ocurre de manera
simultánea, por eso se conoce como maduración heterócrona.
Como cualquier otro proceso de génesis, requiere diferenciación, multiplicación
y migración celular, y está sujeto a simetría:
x Diferenciación: Existe una doble vía de diferenciación, las células precursoras
conocidas como neuroblastos darán lugar a las neuronas y los espongioblastos
acabarán siendo células gliales.
352
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Migración: Las células migran en dirección centrífuga, es decir, alejándose de su
núcleo de origen, y lo hacen a distintas velocidades, de ahí que finalmente el
proceso de maduración en su conjunto sea heterócrono. La migración desigual
dará lugar a la existencia de una sustancia gris y una sustancia blanca con una
organización definida; en el encéfalo, la sustancia gris es periférica y en la
médula es interna. Los cuerpos celulares se agruparán después de la migración
en:
o Núcleos: Tridimensionales
o Capas, láminas o estratos: En esencia, son lo mismo que los núcleos,
pero debido a su aplanamiento son bidimensionales.
Meninges
Las meninges son envueltas conjuntivas cuya función es proteger el SNC. Tres:
x Duramadre o paquimeninge
x Leptomeninges:
o Piamadre
o Aracnoides
Duramadre
Es la meninge más externa. Está constituida por tejido conjuntivo denso rico en
vasos sanguíneos y linfáticos. Se divide en dos partes:
x Craneal: Se continúa con el periostio craneal.
353
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Espinal: No se continúa de forma directa con el periostio del canal vertebral,
sino que lo hace con el espacio epidural en su cara externa.
La superficie interna de la duramadre está constituida por un conjunto de células
limitantes fuertemente unidas, y da al espacio subdural. Este espacio es virtual, solo
aparece en situaciones patológicas, como tras un traumatismo por ejemplo.
La duramadre presenta desdoblamientos que delimitan unos espacios revestidos
por endotelio, conocidos como senos venosos de la duramadre. A ellos llega la sangre
venosa cerebral, y la envían a las venas yugulares internas. Además de estas
formaciones, la duramadre presenta extensiones laminares que penetran hacia el
parénquima nervioso, conocidas como hoces, cuya función es de sostén y protección
puesto que evitan que un traumatismo desplace estructuras nerviosas presentes en el
cráneo, y además permiten la entrada de la aracnoides en los surcos y giros cerebrales
más profundos.
Leptomeninges
Se trata de dos capas de tejido conjuntivo laxo que tienen igual origen, cuyas
células son los meningocitos. Aunque permanecen independientes en la infancia,
realmente en el adulto se fusionan formando una única cubierta a la que se denomina
piaracnoides, cuya hoja parietal es la anterior aracnoides y cuya hoja visceral es la
piamadre primitiva.
Aracnoides
Es la capa media. En su superficie externa, situada por debajo del espacio
subdural, encontramos una nueva membrana limitante, constituida por láminas de
fibroblastos con abundantes uniones intercelulares que se asientan sobre una lámina
basal.
La aracnoides emite trabéculas hacia la
piamadre, que atraviesan el espacio subaracnoideo
envolviendo arterias y venas, y están formadas por
meningocitos estrellados.
A su vez, la aracnoides emite protrusiones hacia los senos venosos de la
duramadre. Estas estructuras están recubiertas de mesotelio y se conocen como
354
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
granulaciones aracnoideas de Pacchioni. Permiten el drenaje del líquido
cefalorraquídeo, que pasa hacia la sangre venosa a favor de gradiente de presión por vía
transcelular (a través de las células mesoteliales) o paracelular (entre ellas).
Con la edad, las granulaciones aracnoideas pueden llegar incluso a penetrar en el
hueso, constituyendo fositas granulosas, e invaginarse formando venas diploicas.
Además, debido a este proceso, es frecuente su calcificación con los años.
Piamadre
Es la capa más interna, por lo que se
halla en contacto directo con la parte más
externa del tejido nervioso, que es la limitante
glial, constituyendo la barrera pío-glial. Se trata
de una membrana formada por 1 o 2 capas
discontinuas de meningocitos aplanados, más
gruesa a la altura de la médula, a la que se
encarga de fijar mediante los ligamentos
dentados y el filum terminale interno y externo.
La piamadre se encarga de envolver
formando una cubierta a modo de embudo
(embudo pial) los vasos sanguíneos que
penetran en el parénquima nervioso.
Clínica: alteraciones meníngeas
x Meningitis: Inflamación de las meninges por colonización infecciosa del líquido
cefalorraquídeo presente en el espacio subaracnoideo.
x Meningiomas: Tumores de meningocitos.
x Hemorragias: rotura de vasos:
o Subaracnoidea: en el espacio subaracnoideo
o Subdural: en el espacio subdural.
Definición
Es un sistema hueco. Los ventrículos están
ocupados por LCR, de manera que tenemos LCR en el
interior y alrededor (en el espacio subaracnoideo).
Luego está flotando entre un líquido interno y uno que
lo rodea. Esto es importante porque si no, el propio peso
del parénquima comprimiría raíces nerviosas y vasos;
además, gracias a que el SNC se encuentra flotando en
LCR, su peso es menor en flotación.
355
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Como hay LCR dentro y fuera, hay dos espacios que además se encuentran
comunicados:
x Espacio Cerebroespinal Externo: Es el espacio subaracnoideo.
x Espacio Cerebroespinal Interno: Es el sistema ventricular. Está formado por
cuatro espacios comunicados denominados ventrículos. Los ventrículos I y II
están en el telencéfalo y se comunican con el III (que está en el diencéfalo) por
los agujeros de Monro. A su vez este se comunica con el IV (del rombencéfalo)
por el acueducto de Sylvio. Por lo tanto, aunque tenemos 4 espacios, el LCR está
en continuidad y puede pasar de uno a otro.
Ambos espacios se comunican a través de dos vías:
x Vía lenta, atravesando todo el parénquima nervioso.
x Vía rápida, en ella los orificios se comunican a través de dos orificios (Luschka
y Magendie). El hecho de que ambos sistemas estén en comunicación, permite
contrarrestar cambios de presión. Los ventrículos I y II son los laterales.
Espacio subaracnoideo
El espacio cerebroespinal externo o espacio subaracnoideo está formado, en la
aracnoides entre la membrana aracnoidea y la piamadre, por un conjunto de cavidades
no siempre comunicadas entre sí pues a pesar de ser continuo existen zonas donde el
espacio se estrecha (hemisferios cerebrales a nivel del telencéfalo) o se amplía (base del
encéfalo). Las zonas donde se dilata se conocen como cisternas, formándose así la
cisterna cerebelomedular, la interpeduncular, quiasmática y la ambiente o superior. Hay
mapas con diferentes localizaciones de cisternas.
El LCR se forma en los plexos coroideos, es de color muy claro, tiene pocas
proteínas de baja densidad (sobre todo inmunoglobulinas), baja densidad celular (1-
5cél/mL), se extrae por punción lumbar y el análisis de este líquido es tremendamente
útil porque permite de entrada un diagnóstico diferencial de enfermedades como
infecciones o Parkinson, no solo avisando del problema sino del tipo de infección o
enfermedad. Recientemente se ha conseguido identificar en el LCR ADN tumoral a
través de la biopsia líquida.
El LCR, que está en dos sistemas de cavidades, está circulando continuamente.
El del sistema ventricular o interno circula generalmente desde posiciones craneales a
caudales (desde los ventrículos I y II al III y después al IV). Tras llegar a este último
ventrículo pasará al sistema cerebroespinal externo por dos mecanismos.
El drenaje del LCR se produce en las granulaciones aracnoideas de Pacchioni a
nivel del encéfalo y en los puntos de salida de los nervios espinales en la médula. Se
calcula que diariamente se drenan a sangre venosa 330-380mL. La circulación del LCR
puede ralentizarse si las células endoteliales se acumulan formando calcosferitos.
356
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Provocan exceso de LCR que lleva a un aumento de presión en las meninges que daña
el parénquima nervioso; hidrocefalia.
El sistema ventricular tiene su luz tapizada por el epéndimo que es epitelio
cúbico simple. En realidad, este epitelio está formado por células que, aunque
morfológicamente son epiteliales, no lo son: células ependimarias o ependimocitos (que
también tapizan a nivel de la médula a este epéndimo). Esta capa de ependimocitos se
apoya en una capa limitante.
Los ependimocitos son células gliales que se unen entre sí para formar una
barrera. Presentan microvellosidades en la membrana apical, algún cilio (aunque no
siempre) y bajo este epitelio tenemos una capa glial subependimaria llamada también
limitante glial interna.
En algunos puntos (sobre todo a nivel del
ventrículo III), entre los ependimocitos se
diferencian unos especiales: los tanicitos. Estas
células serían ependimocitos con prolongaciones
basales que se meten hacia el parénquima
atravesando la limitante glía interna y llegando a
parar a vasos, a capilares. Otro tipo de células que
se adosaban a los vasos sanguíneos en el SN son los
astrocitos. Los tanicitos se piensa que intervienen
en el transporte de sustancias desde la luz, entre el
LCR y la sangre de estos capilares.
357
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Plexos coroideos
Son los puntos donde se sintetiza LCR. Son estructuras replegadas que aparecen
en la pared dorsal de algunos ventrículos. Estos en esta zona, en vez de tener un
epéndimo liso, muestran un epéndimo muy replegado. Tiene forma de colgajo, con un
eje mesodérmico que procede directamente de la piamadre y es por esta zona por donde
entran los capilares fenestrados. Revistiendo el mesodermo, encontramos el epitelio
ependimario modificado. El hecho de que en esta zona el epitelio se repliegue hay que
entenderlo como un mecanismo de aumento de la superficie (200m2).
El eje mesodérmico es un conjuntivo laxo muy celular con fibroblastos y
melanocitos, fibras nerviosas, aferencias (por lo que esta zona es una zona del SNC
estimulable y sensible al dolor), y células pigmentarias. Además, pueden aparecer
concreciones de carbonato cálcico o calcáreas que en este caso reciben el nombre de
acervulus.
Por fuera hay un epitelio muy replegado. No tiene el aspecto típico, sino que hay
un gran desarrollo de vellosidades (células con ribete en cepillo). Tiene gran cantidad de
uniones (adherens y occludens) siendo muy hermético (lo que es fundamental para
sellar e impedir el paso de elementos sanguíneos al LCR) y tiene una membrana basal
tremendamente replegada en laberintos con abundantes bombas de transporte y
mitocondrias intercaladas. Estas células tienen un gran metabolismo y funciones de
transporte. Estas células toman por el polo basal (pía) donde sintetizan el LCR y lo
excretan por el polo apical a la luz ependimaria.
Vienen a sintetizar 0.5 L de LCR diario y este se renovaría varias veces al día,
pues su volumen final es de 150 mL.
Apoyados en el epitelio, en la luz, están los macrófagos de Kolmer (son
estrellados). Tienen que ver con la limpieza de esta zona, pues eliminan restos o células
de naturaleza envejecida para que no lleguen al LCR porque circularían por él.
358
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Vascularización
Esta altamente vascularizado, y las grandes arterias las encontramos en el
espacio subaracnoideo desde donde van a penetrar. Estos vasos penetran en el
parénquima nervioso y durante un tiempo siguen acompañadas por el embudo pial.
Hablamos así de un espacio denominado el espacio de Virchow-Robin para esas zonas
alrededor de los vasos en los que sigue existiendo la limitante glíal-subglial (toda la
zona del embudo es este espacio). Cuando las arterias se ramifican en vasos más
pequeños (capilares) desaparece el espacio de Virchow-Robin.
359
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Barrera hematoencefálica
El SNC tiene una barrera cuya existencia se conoce desde
la antigüedad porque cuando se inyectaban colorantes vitales en
la sangre y después se analizaba el SN, este aparecía limpio, sin
color, por lo que debía de existir algo que impidiera su entrada.
Es una barrera que se forma muy precozmente y hoy en día
sabemos que reside en los vasos, asentándose en tres estructuras:
su endotelio, su lámina basal y las prolongaciones gliales. Se
denomina barrera hematoencefálica y su desintegración produce
un edema cerebral. Los vasos del SNC son capilares continuos.
Las células endoteliales son más delgadas de lo habitual, y tienen muy
restringido el tráfico de moléculas, pues hay pocos receptores de membrana, procesos
de pinocitosis y transportadores para sustancias esenciales como aminoácidos, glucosa,
vitaminas…). Hay ZO que dan un sellado casi hermético.
Alrededor se encuentra la lámina basal y está asociada a astrocitos que forman el
pie vascular. Las células gliales son esenciales para el mantenimiento de las zónulas
occludens del endotelio pues tienen pies vasculares, que son prolongaciones en forma
de embudo de astrocitos que acaban pegados al endotelio.
En los pies hay
muchas acuaporinas,
las cuales juegan un
papel clave en el
mantenimiento de
homeostasis de agua,
para evitar los edemas.
Pero, recordemos
que, en los plexos
coroideos, el locus
ceruleus, locus niger y
neurohipófisis no hay
este tipo de barrera.
360
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Médula espinal
Está alojada en el canal vertebral, es lineal, alargado y posee dos zonas
fusiformes donde es más grueso denominados intumescencia cervical, braquial o
superior y la intumescencia lumbar o inferior.
Tiene una posición dorsal respecto de la notocorda y, como los otros órganos del
SNC, está hueco en su centro formando un conducto conocido como epéndimo o canal
ependimario que se encuentra recubierto por un epitelio ependimario, está bañado por
LCR y protegido por las meninges. Tiene una estructura simétrica y se divide en dos
mitades, las hemimédulas izquierda y derecha.
Se divide en dos regiones, la posterior o dorsal y la
anterior o ventral. Las fibras nerviosas que entran (traen
información sensitiva) lo hacen siempre por posiciones
dorsolaterales formando la raíz dorsal, sensitiva. Las que
salen lo hacen por posiciones ventrolaterales y constituyen la
raíz ventral, motora. Estas raíces son parte del sistema
nervioso periférico. Por tanto, la zona posterior de la medula
tiene naturaleza sensitiva mientras que las órdenes motoras
aferentes salen por posiciones ventrales por las raíces
anteriores o ventrales.
En la raíz posterior tenemos un engrosamiento que
está intercalado en todas las raíces posteriores (menos en la
primera cervical) que se conoce como ganglio sensitivo o
espinal.
Por otro lado, las dos raíces del mismo nivel se unen y
van a formar una única estructura que es el nervio raquídeo o
nervio espiral. Entonces, la raíz posterior que habíamos dicho
que era sensitiva y la anterior que tiene naturaleza motora
forman juntos el nervio espinal con naturaleza mixta.
361
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Tenemos 31 pares de nervios espinales (8 cervicales, 12 torácicos, 5 lumbares, 5
sacros y 1 coccígeo) que salen por los agujeros vertebrales del canal vertebral. Esta
disposición da una imagen de órgano segmentado, aunque no existe segmentación.
Durante el desarrollo embrionario la columna vertebral crece más que la médula
espinal eso significa que aunque inicialmente la médula llena todo el canal vertebral y
cada par de nervios sale del canal a su mismo nivel espinal, después en el recién nacido
la zona inferior o caudal de la medula está a nivel de la 3º vértebra lumbar (por debajo
ya no hay). Ese acortamiento se va acentuando a nivel de la infancia y al llegar al adulto
solo llega a nivel de la L1 o incluso la T12.
Como cada nervio espinal tiene que salir por su agujero, los primeros tienen el
agujero justo enfrente pero después los siguientes tienen que recorrer un espacio hasta
alcanzar el agujero. Todo este conjunto de nervios espinales del canal vertebral se
denomina cola de caballo o cauda equina.
Otra característica de la médula es que su extremo inferior tiene un aspecto
cónico que se adelgaza, el cono medular. Desde ahí sale un filamento pial delgado, el
filum terminale, que ancla el cono medular a la columna vertebral, para dar estabilidad y
que no esté flotando de manera inestable.
En la médula espinal tenemos cuatro surcos que tienen un nombre característico.
Dos de ellos delimitan las hemimédulas izquierda y derecha, la cisura mediana anterior,
por la que va la arteria nutricia, y el surco mediano posterior. Laterales encontramos el
surco anterolateral y el posterolateral, más o menos a la altura de la salida y entrada de
las raíces espinales. Por último, encontramos un tabique medio posterior, glial.
La espina bífida es una malformación congénita que se da cuando los arcos
neurales no se cierran adecuadamente (ocurre en la zona lumbar-sacra). Es asintomática
y se puede ver a través de técnicas de imagen como las radiografías. Hay muchos tipos
de espinas bífidas que sí dan síntomas, ya que la separación de los arcos da un espacio
tal que se produce una protrusión hacia fuera de las meninges. En un 10% es
meningocele; pero la mayoría de pacientes presentan mielomeningocele, donde el
problema radica en que no solo se protruye fuera del arco meninges, sino también
parénquima nervioso. Con ello se da salida de LCR, puede haber hidrocefalia, daños
neurales…
En el SNC se da una migración celular en el desarrollo, y no es a la misma
velocidad. En el caso de la médula espinal, la sustancia gris queda interna y se describe
como una sustancia gris en forma de H o alas de mariposa. Forma un asta dorsal
(sensitiva), lateral o intermedia (visceral) y el ventral o anterior (motora) en cada
hemimédula. En las astas encontramos neuronas o grupos de neuronas (núcleos)
característicos.
Desde el punto de vista funcional podemos dividir la médula en 4 regiones
funcionales. En la médula dorsal tenemos una zona somato-sensitiva (neuronas que
362
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
reciben información sensitiva que tiene que ver con órganos como el músculo) y otra
víscero-sensitiva (reciben información de las vísceras), mientras que en la ventral hay
una zona víscero-motora y otra somato-motora. Ambas regiones se separan por el surco
de Hiss (durante el proceso embrionario) que dividiría la médula en una región dorsal y
una ventral. Después este surco desaparece.
La sustancia blanca es periférica, está formada por fibras mielínicas y se
organiza en tres cordones por cada hemimédula. El cordón dorsal va del surco medio
posterior al asta posterior, el lateral está entre el asta dorsal y ventral, y el anterior entre
la cisura media anterior y el asta ventral.
Sustancia gris
En la sustancia gris las neuronas se agrupan en pequeños núcleos. Cuando tiene
forma bidimensional hablamos de capa o estrato.
Un núcleo es un conjunto de neuronas que ocupan una posición concreta y que
morfológicamente son del mismo tipo (son idénticas) y que además llevan a cabo la
misma función (llevan las mismas aferencias y están conectadas al mismo destino).
Derivan de un precursor común y migran juntas a su posición definitiva. A cada uno se
le da un nombre propio para diferenciarlos entre sí.
En el asta dorsal encontramos una base, un cuello y una cabeza con diferentes
núcleos:
x En la base, el núcleo torácico o de Clarke, que se identifica en los niveles
torácicos.
x En el cuello del asta encontramos el núcleo propio.
x En la cabeza hay varios núcleos que son la capa marginal de Waldeyer, la
sustancia esponjosa o zona marginal y la sustancia gelatinosa de Rolando.
En el asta lateral hay dos núcleos que por su posición se denominan núcleo
intermedio lateral y núcleo intermedio medial. Esta nomenclatura se fija en la posición
que ocupan y es la que se aplica también en el siguiente asta.
Finalmente, los núcleos del asta ventral son tres y se denominan lateral, central y
medial. Sin embargo, se suele hablar de seis núcleos porque cada uno se desdobla en
dos partes, uno más pequeño en posición dorsal y el resto en posición ventral. Estos
núcleos se denominarán dorsolateral, ventrolateral, dorsocentral, ventrocentral,
dorsomedial y ventromedial.
Otra manera de hablar de un determinado núcleo puede ser a partir de la
clasificación de Rexed que consiste en dividir en 10 zonas. De esta manera, los diez
territorios laterales se laminarían, disponiéndose los núcleos grises en los diferentes
territorios: Las láminas I-VI ocupan el asta dorsal, la lámina X ocupa el territorio
circunependimario (alrededor del epéndimo) y las láminas VII-IX se encuentran en el
asta ventral.
363
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Los núcleos mencionados no son fijos, hay algunos que sí, pero otros no. En
función de la sección que estemos viendo encontraremos unos núcleos u otros.
Tipos celulares
La médula espinal no es siempre igual pues las secciones y la forma de las astas
varían en tamaño. En primer lugar, tenemos células gliales de diferentes tipos dentro de
la glía: ependimocitos (epitelio cúbico simple del epéndimo), astrocitos protoplásmicos,
microglía y oligodendroglía.
Entre las neuronas encontramos neuronas radiculares y de asociación. Las
radiculares son aquellas cuyo axón sale del SNC y pasa a formar parte del SNP. No
todas las radiculares son iguales, sino que dentro de ellas hay dos grandes grupos, las
motoneuronas y las neuronas viscerales. Las motoneuronas tienen el soma en el asta
ventral y dentro de ellas se distinguen las α y γ; las motoviscerales o neuronas viscerales
motoras tienen en el soma en el asta intermedia o lateral. Las neuronas de asociación
son aquellas cuyo axón siempre permanece en el SNC. Las hay cordonales, funiculares
y Golgi II.
Motoneuronas
Como ya se ha mencionado hay dos tipos, α y γ:
364
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Motoneuronas α: “Son las reinas, las más grandes (como la Pantoja)”. Tienen
morfología estrellada por las dendritas muy ramificadas que salen de cualquier
punto de su soma. En este soma gigante (+100μm) tenemos un núcleo grande,
nucléolo marcado y un citoplasma basófilo pues abundan los gránulos de Nissl
debido a su importante actividad metabólica
El axón sale del asta ventral (pues ahí está el soma) por la raíz ventral y tiene
como destino final el músculo esquelético formando una unión característica
conocida como placa motora (la segunda sinapsis es el miocito) que regula su
contracción. Además, durante el recorrido puede dar colaterales que llegan a
cualquier parte.
Está todo muy bien colocadito, según el músculo que se va a inervar, la neurona
se coloca específicamente. Los del tronco están inervados por motoneuronas α
que están en posiciones mediales. Si queremos inervar un músculo más lateral
como una extremidad: la motoneurona α está en una posición más lateral.
También se diferencia según el tipo de músculo inervado: flexor o extensor. Las
motoneuronas colocadas más anteriormente son musculatura extensora, mientras
que si los somas de las motoneuronas del asta ventral se colocan más
posteriormente, el músculo es flexor.
x Motoneuronas γ: Son las princesas. No son tan grandes como las motoneuronas
α. El axón no forma una placa motora, sino que inerva una estructura llamada
huso neuromuscular, de manera que las órdenes motoras de estas motoneuronas
controlan el tono muscular.
Neuronas Viscerales
Se colocan en el asta lateral. Son grandes, pero son distintas a las motoneuronas,
pues tienen forma fusiforme. Antes se decía que tenían forma de pececitos.
Tienen muchas dendritas que salen de dos polos (penachos dendríticos u
opositopolares). De ellas sale el axón de posición media y abandona la médula espinal
por la raíz ventral o anterior (como en las motoneuronas) y van a inervar musculatura
visceral, nunca somática.
Estas neuronas viscerales también se pueden denominar neuronas
preganglionares del SN Autónomo o vegetativo, se encuentran en el asta lateral y se
clasifican en dos grupos, simpáticas y parasimpáticas. Las simpáticas son las que están
en el asta lateral en los niveles dorsal a lumbar (T1-L2) mientras que las parasimpáticas
son las que están por debajo de lumbar (S2-S4). Los núcleos intermedio lateral y
medial son los que contienen estas neuronas.
Son también llamadas neuronas preganglionares puesto que no se dirigen
directamente a la víscera sino a un ganglio. Su axón sale por el asta ventral y va a un
ganglio que forma parte del Sistema Nervioso Vegetativo.
365
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Neuronas cordonales
Están en el asta posterior y reciben información sensitiva. Su soma es pequeño y
su axón permanece en la sustancia blanca, teniendo forma de T, por lo que no sale de la
médula. La bifurcación en T da lugar a una rama ascendente y a otra descendente,
Neuronas funiculares
Se localizan en el asta posterior y reciben información sensitiva. Su soma es
pequeño y el axón se dirige a la sustancia blanca ramificándose en forma de T:
x Rama ascendente: Muy larga. Sale de la médula, aunque no llega al SNC,
llevando información sensitiva a regiones encefálicas.
x Rama descendente.
Neuronas Golgi II
Son pequeñas, con el axón corto y muy ramificado. No salen hacia la sustancia
gris, sino que permanecen en la sustancia blanca.
Dentro de este grupo se encuentran las neuronas de Renshaw, que se encuentran
asociadas a motoneuronas α ejerciendo sobre ellas un efecto inhibitorio (permanente).
Neuronas comisurales
El axón de estas neuronas cruza y pasa a la hemimédula opuesta, de tal forma
que sus fibras forman las comisuras grises anterior y posterior. Pueden ser cordonales,
funiculares o Golgi II.
Sustancia blanca
La sustancia blanca contiene axones mielínicos y amielínicos que transcurren de
uno a otro segmento de la médula, entre ellos hay células giales como los astrocitos
fibrosos, oligodendroglía y microglía.
Como se ha dicho anteriormente, la sustancia blanca se divide en cordones, cada
uno de los cuales muestra diferentes fascículos. Un fascículo o tracto es un conjunto de
fibras nerviosas con igual origen y destino. Pueden tener un nombre propio o compuesto
(origen-destino). Se distinguen fascículos ascendentes y descendentes.
Fascículos ascendentes
x Cordón dorsal:
o Grácil o de Goll (medial) y cuneatus, cuneiforme o de Buldarch (lateral):
Las fibras terminan en los núcleos de Goll y de Buldarch, situados en el
bulbo raquídeo. Dan lugar a colaterales descendentes.
o Haz semilunar de Schultze (cervical).
o Fascículo septomarginal o campo oval de Flechsig (torácico).
o Fascículo triangular de Philippe-Gombault (sacro).
366
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Cordón anterolateral:
o Espino-cerebeloso posterior, dorsal o de Flechsig: Las fibras salen del
núcleo de Clarke y llegan al pedúnculo cerebeloso inferior, llevando
información propioceptiva.
o Espino-cereboloso anterior, ventral o de Gowers: Las fibras salen de las
láminas V a VII, llevando información propioceptica y exteroceptiva.
o Espino-talámico lateral: Llevan información térmica y dolorosa.
o Espino-talámico anterior, ventral: Con información táctil y de presión.
o Espino-tectal: Contienen información dolorosa, táctil y térmica.
o Espino-olivar: Fibras cervicales que tienen como destino la oliva bulbar
y llevan información propioceptiva.
o Espino-vestibular: Fibras de origen cervical que llegan a los núcleos
vastibulares con información propioceptiva.
Las fibras de ambos fascículos espino-talámicos salen del núcleo propio y llegan
hasta el tálamo habiéndose cruzado previamente.
x Cordón anterior:
o Cortico-espinal directo: Anterior. Las fibras salen de las áreas 4 y 6 de la
corteza cerebral y se decusan en niveles inferiores. Van a interneuronas,
desde donde se dirigen a motoneuronas. Se encargan del control
voluntario de movimientos finos y el control de las aferencias.
La vía piramidal es la corticoespinal cruzada y se dirigen a interneuronas
de asociación.
o Vestíbulo-espinal: Control del tono muscular y del equilibrio.
o Retículo-espinal medial o póntico: Activa a la musculatura
antigravitatoria extensora e inhibe a la flexora.
o Tecto-espinal.
o Longitudinal medial.
367
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Variaciones
Las médulas espinales no son uniformes, sino que son más anchas a nivel de las
intumescencias: C4-C5 y L4-L5.
La sustancia blanca crece al ascender
por la incorporación de fibras ascendentes y la
terminación de fibras descendentes.
A nivel de sustancia gris, el asta dorsal
es más estrecha a nivel cervical, con mayor
desarrollo de la sustancia esponjosa; mientas
que a nivel lumbar y sacro es más gruesa y
hay mayor desarrollo de la sustancia
gelatinosa.
En el asta ventral la sustancia gris está
muy desarrollada a nivel cervical, lumbar y
sacra (núcleos dorsolateral, ventrolateral,
ventromedial, dorsomedial, dorsocentral y
ventrocentral).
La sustancia gris del asta lateral está
muy desarrollada a nivel torácico (núcleo
intermedio lateral).
Arcos reflejos
Las fibras aferentes llegan al asta posterior y se genera una respuesta directa sin
intervención de las áreas superiores. Esto ocurre en una situación de peligro o alerta,
cuando el organismo desarrolla una respuesta refleja más rápida.
Esto supone que además de la neurona que recibe la información sensitiva
intervenga una motoneurona (reflejo monosináptico) o también puede intervenir una
neurona de asociación, con una interneurona (reflejo multisinaptico).
368
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Espina bífida
Malformación congénita producida por un defecto en la unión de los arcos
neurales, principalmente en la región sacra. Puede ser asintomática y solo verse
mediante radiografías, es conocida como oculta. También puede ser que exista una
protrusión de material:
x Mielomeningocele: Es la forma más grave. En esta malformación un saco o
quiste de líquido sobresale en una abertura en la espalda. El saco contiene parte
de los nervios de la médula espinal. Esta forma de espina bífida causa
discapacidades de leves a graves, como problemas que afectan la manera en que
una persona va al baño, pérdida de la sensibilidad en las piernas o los pies,
discapacidad para mover las piernas, daños neuronales, pérdida de líquido
cefalorraquídeo, hidrocefalia…
x Meningocele: Saco de líquido que sobresale por una abertura en la espalda. Por
lo general, el daño en los nervios es mínimo o no hay daños.
Lesiones de la médula espinal
x A nivel torácico o lumbar: paraplejía, parálisis miembros inferiores
x A nivel cervical: tetraplejía, parálisis completa (excepto cabeza).
En los primeros días aparece anestesia generalizada, parálisis de movimiento,
miembros fláccidos (faltan reflejos tendinosos), parálisis de vejiga y recto.
369
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Se observa hiperpolarización de las neuronas espinales por debajo del nivel de la
lesión (liberación masiva de glicina). Estos síntomas desaparecen y se restauran parte de
los reflejos espinales (no se restauran con normalidad sino que los movimientos
derivados se hacen bruscos), tono muscular espástico y se reconstruye el arco reflejo de
la vejiga automática.
Buena parte de las lesiones son secundarias a cambios en las concentraciones de
electrolitos y alteraciones vasculares.
Síndrome de la motoneurona
Las enfermedades de la motoneurona (EMN) son un grupo heterogéneo de
enfermedades, algunas de ellas hereditarias, con afectación exclusiva o predominante de
las neuronas motoras de la corteza cerebral y/o de los núcleos motores del
troncoencéfalo (preservándose habitualmente los núcleos oculomotores) y/o de las astas
anteriores de la médula espinal.
La superior seria la neurona motora que elabora la orden y está en la corteza
mientras que la inferior está en el tronco encefálico o medula espinal. Se habla entonces
de enfermedad de la motoneurona superior y la inferior.
La enfermedad de la motoneurona superior es la interrupción del tracto cortico-
espinal en algún punto. Los signos se ven en el mismo lado o en el contrario en función
de que la interrupción sea antes o después de la decusación. Se observa parálisis
fláccida de la zona afectada, perdida de reflejos tendinosos, se pierde el tono muscular
normal y a las semanas se recuperar pero se hace espástico. Aparece el clonus de tarso o
carpo y el signo de Babinksi. Faltan reflejos abdominales. Tras un ataque
cerebrovascular aparece hemiplejía fláccida seguida de hemi-parestesia espástica
permanente.
En la enfermedad de la motoneurona inferior ambas motoneuronas van a acabar
degenerando. Estos signos no son a la vez sino que lo más normal es que degenere
primero la inferior. Al cabo de los años la superior se ve también afectada, por lo que
los miembros inferiores también se debilitan y acaban atrofiándose. Engloba cualquier
tipo de signo o síntoma como consecuencia de las motoneuronas inferiores hacia el final
de la mediana edad. Suele deberse a agentes infecciosos, como es el caso de la polio.
En resumen, ambas motoneuronas (superior e inferior) acaban degenerando.
Estos signos no son a la vez sino que lo más normal es que genere primero la inferior.
Los dos primeros años afecta a motoneuronas a nivel espinal y se refleja atrofia y
alteraciones de la motilidad e lo miembros superiores. Cada vez es más progresiva y al
cabo de 2 o 3 años se aumenta: los reflejos se hacen muy bruscos y aumenta el tono
muscular los miembros inferiores se debilitan. A esto se le conoce como ELA o
esclerosis lateral amiotrófica.
370
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
SISTEMA NERVIOSO PERIFÉRICO
Es aquel que no presenta simetría bilateral, sino que es plexiforme. Se denomina
plexiforme porque es un conjunto de redes o plexos formados por distintas neuronas
cuyas prolongaciones contactan entre sí, que se encargan de la transmisión lineal de la
información aferente y eferente y de mejorarla al potenciar y suprimir distintos
impulsos. Además de transmitir tiene función de control en el procesamiento de la
información. Es decir, que no nos tenemos que imaginar los ganglios y nervios como
meros conductores o relevos de la información: también le dan forma.
El SNP aparece primero en la ontogenia, y por tanto ejerce una función de
control, pero después con el desarrollo del SNC pierde sus capacidades en favor de este.
Está compuesto de nervios periféricos con terminaciones nerviosas especializadas
(fibras aferentes y eferentes) y ganglios que contienen los somas neuronales que están
fuera del SNC.
Ganglios nerviosos
Raquídeos o espinales
Contienen los somas de neuronas sensitivas o aferentes. La información
sensitiva recogida en la periferia por estas neuronas llega al soma del ganglio, y desde
ahí viaja al SNC por la terminación axónica. Por tanto, se trata de ganglios sensitivos en
los que no hay ningún tipo de sinapsis, la información simplemente es conducida por
una neurona.
Estos ganglios son engrosamientos situados en la raíz dorsal (aferente) del
nervio espinal, y se asemejan a los interpuestos en la trayectoria de los nervios
craneales.
Estroma
Está formado por la cápsula y el tejido de soporte.
x Cápsula: Se encuentra en continuidad con el
epineuro del nervio espinal, y por tanto se trata
como este de una envuelta conjuntiva
estratificada, que emite tabiques que penetran
hacia el interior del ganglio. Obviamente, los
elementos que entran y salen del ganglio, como
los vasos sanguíneos, han de atravesar la cápsula.
x Conjuntivo: Es continuo con el endoneuro, y
muestra fibras colágenas y abundantes células
conjuntivas (fibroblastos, mastocitos…). Sin
embargo, nunca hace contacto con el tejido
nervioso.
371
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Parénquima
El parénquima de los ganglios
raquídeos se encuentra segregado, de tal
modo que:
x Periferia: Podemos hablar de
corteza, en ella se encuentran los
somas neuronales (sustancia gris)
x Centro: Podemos denominarlo
médula, en ella hallamos las
fibras (sustancia blanca).
Está constituido por células nerviosas, es decir, por neuronas y glía.
x Glía: se encuentra interpuesta entre las neuronas y el conjuntivo. Hallamos:
o Células libres: Se encuentran en el conjuntivo.
o Células de Schwann: Se asocian a las fibras nerviosas de las neuronas
ganglionares.
o Células satélite o capsulares: Se asocian al soma de la neurona ganglionar.
Forman una vuelta completa alrededor del soma y del cono axónico, a modo
de corona. Las células de esta corona se encuentran unidas entre sí, y a la
neurona. Su citoplasma es delgado y con gliofilamentos PGFA+, y su núcleo
es ovalado y denso.
x Neuronas: En etapas iniciales de la ontogénesis son bipolares, pero después las
terminaciones se acaban fusionando y pasan a ser pseudomonopolares. Su
tamaño varía, de modo que pueden ser gigantes, grandes, medianas y pequeñas.
En cualquier caso, su citoplasma es rico en orgánulos (llegamos a visualizar
pequeños grumos de Nissl) y en lipofucsina, y su núcleo es grande, pálido y con
un nucléolo marcado.
A este nivel hemos de definir una estructura llamada glomérulo. Se trata de un
segmento de la prolongación de la neurona situado entre el soma y la bifurcación en T
típica de las neuronas pseudomonopolares. A este nivel, la prolongación adquiere una
trayectoria retorcida, y se halla asociada a todavía a gliocitos capsulares, mientras que
las dendritas y el axón ya se asocian a células de Schwann.
Por tanto, tras la bifurcación en T ambas fibras son mielínicas. De ellas, hay una
rama más gruesa que se dirige a la periferia y acaba en algún receptor, encargado de
transmitir la información aferente (la dendrita) y otra más fina que se dirige a la médula,
estructura del SNC, y lleva las aferencias a través de la raíz dorsal. Por tanto, el trayecto
de la información sería el siguiente:
Receptor Æ dendrita Æ soma del ganglio raquídeo Æ axón Æ asta dorsal de la
médula espinal
372
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Los ganglios existentes en el SNC son similares a los raquídeos. Se trata de los
ganglios de los pares craneales:
x V: ganglio de Gasser, semilunar o trigeminal
x VII: geniculado
x VIII: ganglio espiral o de Corti (este es diferente, aquí las neuronas son bipolares
verdaderas y no pseudomonopolares, es del nervio auditivo).
x VIII: ganglio de Scarpa o vestibular.
x IX y X: cada uno presenta dos ganglios, uno superior y uno inferior.
373
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Vegetativos
Son los ganglios del sistema nervioso autónomo: SNS y SNPS. Se trata de
ganglios en los que sí existen sinapsis y que son motores: contienen neuronas simpáticas
y parasimpáticas que responden a las eferencias enviadas por el SNC. Según
pertenezcan a una división u otra del SNA, tienen distinta localización:
x SNS: Sus ganglios forman parte de la cadena ganglionar simpática o tronco
simpático paravertebral, o bien se trata de ganglios prevertebraes (celíaco,
mesentéricos superior e inferior y aórticorrenal).
x SNPS: Sus ganglios pueden ser:
o Cefálicos: Son los ganglios de los pares craneales.
Ciliar (III)
Submaxilar (VII)
Esfenopalatino (VII)
Ótico (IX)
o Terminales
Su estructura histológica es similar a la de los anteriores.
Estroma
Está formado por la cápsula y el tejido de soporte.
x Cápsula: Se encuentra en continuidad con el epineuro
del nervio espinal, y por tanto se trata como este de
una envuelta conjuntiva estratificada, que emite
tabiques que penetran hacia el interior del ganglio.
Obviamente, los elementos que entran y salen del
ganglio, como los vasos sanguíneos, han de atravesar
la cápsula.
x Conjuntivo: Es continuo con el endoneuro, y muestra
fibras colágenas y abundantes células conjuntivas
(fibroblastos, mastocitos…). Sin embargo, nunca
hace contacto con el tejido nervioso.
Parénquima
Está también constituido por neuronas y glía, pero en esta ocasión no se
encuentra segregado, los elementos se distribuyen por igual en centro y periferia:
x Glía: Se encuentra interpuesta entre las neuronas y el conjuntivo. Hallamos:
o Células libres: Se encuentran en el conjuntivo.
o Células de Schwann: Se asocian a las fibras nerviosas de las neuronas
ganglionares.
o Células satélite o capsulares: Se asocian al soma de la neurona ganglionar.
Forman una vuelta completa alrededor del soma y del cono axónico, a modo
374
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
de corona. Las células de esta corona se encuentran unidas entre sí, y a la
neurona. Su citoplasma es delgado y con gliofilamentos PGFA+, y su núcleo
es ovalado y denso.
x Neuronas: Son neuronas estrelladas multipolares, cuyo tamaño también puede
ser grande, mediano o pequeño. Presentan múltiples dendritas, y su axón no está
mielinizado y sale por la rama comunicante gris para incorporarse al nervio.
Existen dos tipos de neuronas:
o Dogiel I: Sus dendritas son cortas y están encerradas por una corona glial.
o Dogiel II: Sus dendritas son largas y estás asociadas a gliocitos. Si todas las
dendritas de una neurona Dogiel II salen del mismo polo de la célula,
adquieren el aspecto de una cola de cometa, de ahí que llamemos a las
células en las que esto sucede neuronas en cometa.
El glomérulo en este caso es la estructura espiral que se forma cuando las fibras
que llegan al ganglio se enrollan en torno a las dendritas. Las sinapsis que se
establecen a este nivel son sinapsis en passant.
Existen otro tipo de neuronas de pequeño tamaño conocidas como SIF, que son
neuronas catecolaminérgicas que emiten por ello fluorescencia.
Fibras aferentes
Se trata de fibras preganglionares del SNA procedentes de neuronas motoras
viscerales. Viajan por el nervio raquídeo y la rama comunicante blanca, y tras ello
tienen 3 posibilidades:
x Hacer sinapsis con las multipolares.
x Pasar de largo para llegar a un ganglio de la cadena de nivel superior o inferior
donde harán sinapsis.
x Pasar de largo y salir por la rama visceral para alcanzar un ganglio fuera de la
cadena paravertebral, en el que harán sinapsis.
Fibras eferentes
Son los axones de las neuronas de los ganglios vegetativos, los cuales alcanzan
el órgano diana y establecen sinapsis noradrenérgicas (SNS) o colinérgicas (SNPS).
Nervios
Al igual que los ganglios, los nervios presentan un estroma conjuntivo asociado
en forma de envueltas:
x Epineuro: Es la envuelta externa. Está constituido por tejido conjuntivo denso no
modelado que rodea todo un nervio periférico. Puede estar asociado a tejido
adiposo, y contiene vasos cuyas ramas penetran en el perineuro.
x Perineuro: Es un conjuntivo especializado que rodea cada fascículo de fibras
nerviosas. Está constituido por 1-5 capas de células aplanadas, todas ellas
selladas con el objetivo de constituir una barrera de difusión activa que mantiene
375
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
el medio iónico de cada haz. Entre las capas de células podemos hallar fibrillas
colágenas y algún mastocito, pero nunca fibroblastos ni células linfoides. Las
células que conforman las capas se comportan como:
o Epiteliales: Dado que mantienen uniones estrechas que las sellan, el
intercambio de sustancias requiere que en sus membranas existan
abundantes receptores y transportadores. Además, tanto en la superficie
interna como en la externa encontramos láminas basales, como en todo
epitelio.
o Musculares: Aunque no tienen filamentos de tubulina, sí poseen filamentos
de actina que les otorgan capacidad contráctil.
o Fibroblastos: Sintetizan el colágeno que hay entre las capas.
x Endoneuro: Comprende el tejido conjuntivo laxo que rodea cada fibra nerviosa
individual. Está constituido por fibras colágenas (sintetizadas por las células de
Schwann), y fibroblastos y mastocitos como únicos tipos celulares. Su
vascularización es escasa, por lo que su nutrición tiene lugar a través de la vaina
perineural.
El parénquima nervioso está constituido por fibras nerviosas asociadas a
elementos gliales y a células de Schwann, que pueden ser mielínicas o amielínicas.
Nervios espinales
Esto está en el tema de médula. Aquí introducimos un concepto nuevo, que es el
de zona de Redlich-Obersteiner, que constituye el límite entre SNC y SNP. Aquí se
adelgazan las vainas de mielina.
Ramas de los nervios espinales
x Recurrente o meníngea.
x Dorsal: Eferente a la musculatura dorsal y aferente para la piel de la espalda.
x Ventral: Eferente a la musculatura anterior y lateral y aferente para la piel del
tronco y las extremidades.
x Comunicante:
o Blanca: Eferente preganglionar a la cadena paravertebral.
o Gris: Eferente ganglionar.
Organización funcional de las fibras
El criterio de clasificación es triple:
x Visceral/somático: Visceral se refiere a glándulas, vísceras y musculatura lisa.
x Aferente/Eferente: Sensitivo/motor.
x General/Especial: Especial se relaciona con los órganos de los sentidos
cefálicos.
376
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x ASG: Este componente lo generan la extero y la propiocepción. La
exterocepción se refiere a receptores de tacto, Tª, corriente eléctrica y dolor; la
propiocepción a los receptores sensitivos de temperatura y dolor muscular
presentes en tendones y articulaciones.
x AVG: Es la interocepción, y se refiere a los receptores sensitivos de temperatura
y dolor visceral, quimiorreceptores del cuerpo aórtico y el seno carotídeo y
mecanorreceptores viscerales.
x ASE: Receptores de retina y oído.
x AVE: Quimiorreceptores de olfato y gusto.
x ESG: Musculatura somática.
x EVG: Musculatura lisa.
x EVE: Musculatura branquiomérica.
ORGANIZACIÓN HISTOLÓGICA, SISTEMA NERVIOSO CENTRAL Y
PERIFÉRICO
Parte del Sistema Nervioso Autónomo se asienta en el Sistema Nervioso Central.
En este SNC tenemos:
x El hipotálamo que es un órgano integrador máximo del sistema autónomo o
vegetativo, cumpliendo funciones importantes porque tiene además que ver
mucho con el sistema endocrino hipofisario.
x Los núcleos neuronales (neuronas motoras). Los núcleos simpáticos son
neuronas preganglionares del asta lateral (nivel dorsal y lumbar).
Además, en el caso de la componente parasimpática tenemos también neuronas
en centros superiores (en el tronco cerebral) como el núcleo de E-W, salival
superior, inferior y dorsal vago.
Dentro del SNC tenemos elementos que forman parte del vegetativo. Los
núcleos neuronales tienen neuronas motoras que envían eferencias a las vísceras.
Para el SNPS encontramos los del asta lateral de la médula espinal (S2-S4) y en
377
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
el tronco cerebral el de Edinger-Westphal, mucolacrimonasal, salivales y el
dorsal del vago.
Para el SNS tenemos los núcleos del asta lateral de la médula espinal (T1-L2).
Núcleos simpáticos y parasimpáticos
Los núcleos simpáticos están en el asta lateral de la médula espinal,
concretamente en los niveles dorsal y lumbar (T1-L2). Contienen neuronas
preganglionares cuyos axones salen por la raíz ventral. Pasan al nervio espinal y se
dirigen por la rama comunicante blanca hasta el ganglio simpático con neuronas
postganglionares. De él sale la rama comunicante gris y llega a vasos, vísceras y
músculos.
Los núcleos parasimpáticos sacros están en el asta lateral (S2-S4) y contienen las
neuronas preganglionares. Salen por la raíz ventral al nervio espinal y siguen un nervio
hasta llegar al ganglio parasimpático pélvico que contiene las neuronas
postganglionares. Sus somas viajan por el nervio esplácnico e inervan las vísceras
pelvianas y el colon descendente. También hay núcleos parasimpáticos que no están a
nivel espinal.
Núcleo de Edinger-Westphal
El núcleo de Edinger-Westphal o motor ocular común tiene neuronas
parasimpáticas cuyo axón sale por el III par o nervio motor ocular común y llega al
ganglio ciliar. Se producen sinapsis y los axones de la segunda neurona salen del
ganglio ciliar por los nervios ciliares cortos, que llegan hasta el músculo ciliar y el
constrictor del iris.
Núcleo muco-lacrimo-nasal
Del núcleo muco-lacrimo-nasal salen los axones a través del VII par o nervio
facial. Toman una desviación por el nervio petroso superficial mayor y sinaptan en el
ganglio esfenopalatino. Los axones de las postganglionares parten hacia sus dianas a
través del nervio maxilar superior. Dichas dianas son las glándulas y vasos de la nariz,
órbita y paladar.
Núcleo Salival Superior
En el núcleo salival superior los axones de las neuronas parasimpáticas salen por
el VII par o nervio facial y se desvían por el nervio cuerda del tímpano. Alcanzan el
ganglio submaxilar y envían sus axones por nervios hasta las glándulas submaxilar y
sublingual y a una parte de la lengua.
Núcleo Salival Inferior
Del núcleo salival inferior tenemos neuronas motoras parasimpáticas cuyos
axones salen por el IX par o nervio glosofaríngeo y a través del nervio petroso
superficial menor alcanzan el ganglio ótico. Se produce la sinapsis y las segundas
378
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
neuronas mandan la orden por sus axones a través del nervio maxilar inferior. Inerva la
glándula parótida y toda la mucosa oral.
Núcleo Motor Dorsal del Vago
El núcleo motor dorsal del vago o cardioneumoentérico contiene neuronas
motoras parasimpáticas que mandan los axones por el X par o nervio vago y después
toman diferentes rutas puesto que inervan una amplia variedad de dianas (nervio
cardiaco, pulmonar, esofágico o vísceroabdominal). Cuando el vago se ramifica, las
neuronas alcanzan los ganglios que están en la pared de la víscera o muy cerca (son
ganglios terminales). Es decir, cada nervio se dirige a una víscera y dentro de ella se
localizan distintos ganglios terminales. Estos ganglios terminales poseen las segundas
neuronas y sus axones son muy cortos. Las fibras llegan a corazón, pulmón, esófago y
vísceras abdominales.
Divergencia
Una característica del SNA es la divergencia. Las neuronas preganglionares son
limitadas en número, pero consiguen un efecto divergente. Se refiere a que una neurona
preganglionar sinapta con muchas postganglionares de forma que esta orden afecta a
muchas células y su efecto es mucho mayor. Es propio de los dos, pero es más notable
en la porción simpática. Es la base del “efecto de masa”.
No siempre el axón que viene de la neurona del asta lateral entra sino que baja a
otro ganglio o sube a otro, es decir, que el punto de sinapsis no es siempre el mediado.
Esta divergencia permite amplificar el efecto lo que significa efecto de masa (inervando
multitud de neuronas que a su vez inervan multitud de haces). Por tanto, cada neurona
del ganglio recibe aferencias de varias neuronas.
Ganglios paravertebrales y prevertebrales
Los ganglios de la cadena paravertebral son a nivel cervical el superior, medio e
inferior. El inferior se fusiona con los torácicos (T10-T11) y forman el ganglio
estrellado. Hay cuatro lumbares, cuatro sacros y uno coccígeo. Forman la cadena
simpática a ambos lados de la y paralela a la columna vertebral.
Los ganglios prevertebrales contienen neuronas simpáticas pero no forman parte
de esa cadena. Son los celíacos, el mesentérico superior e inferior y el plexo
hipogástrico superior e inferior.
379
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
CEREBELO
Es un órgano que forma parte del encéfalo. Está situado en el IV ventrículo y se
encuentra oculto por los hemisferios cerebrales. Recubre parte del bulbo raquídeo. Se
forma a partir de la placa alar del tronco encefálico. Su cara superior está cubierta por el
cerebro y su cara anterior a su vez se expande y cubre el bulbo raquídeo. Cerebelo
significa “pequeño cerebelo”.
Esta parte tiene como función la coordinación de los movimientos corporales. Se
encarga de la sintonización fina (movimientos finos, delicados en vez de bruscos) y
además regula el tono muscular.
Morfología externa
Consta de dos hemisferios cerebelosos y una zona impar y media que es la
vermis. La superficie del cerebelo no es lisa, presenta surcos, elevaciones, que son las
circunvoluciones. En este caso son estrechas y paralelas entre sí. Es un mecanismo de
aumento de superficie.
Se estructura es en laminillas cerebelosas ramificadas o folia cerebelli. Cuando
observamos una laminilla vemos ramificación tiene una estructura arborescente en su
corte sagital que ha hecho que se creyera que aquí se asentaba el alma (árbol de la vida).
Estas laminillas tienen un eje central de sustancia blanca y una periferia la sustancia gris
cubierta por meninges. Cada una es una unidad anatomo-funcional (histona).
El cerebelo se une al tronco cerebral mediante los pedúnculos cerebelosos. Hay
tres pedúnculos cerebelosos, el inferior o cuerpo restiforme, el medio o brazo pontino, y
el superior o brazo conjuntivo.
380
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Inferior o Cuerpo Restiforme
Sale de la medula oblongada y alcanza el cerebelo. Encontramos vías
espinocerebelosas y vías que van o vienen al vestíbulo o a la oliva.
Medio o Brazo Pontino
Va al puente. Aquí encontramos las vías pontocerebelosas (son vías que
transmiten información que llega al puente desde corteza cerebral). Luego llevan
información que ha sido procesada en corteza y sale como orden, dando continuidad a
las fibras cortico-pontinas.
Brazo Conjuntivo o Superior
Contiene vías eferentes cerebelosas para el tálamo y el núcleo rojo. Une cerebelo
y mesencéfalo.
Organización interna
Inicialmente en el desarrollo el cerebelo se divide en una parte pequeña, el
lóbulo flóculo-nodular, y una más grande, el cuerpo, separadas por un pliegue
denominado surco postero-lateral. A su vez el cuerpo se divide en dos lóbulos, uno
anterior y otro posterior, separados por otro surco que es la fisura prima.
En el cerebelo no se desarrolla todo a la vez, establecemos 3 periodos. Se divide
en diferentes zonas según el momento en que aparecen y su desarrollo.
Arquicerebelo o vestíbulo-cerebelo
Es la parte más primitiva y está funcionalmente unida con los núcleos
vestibulares (por lo que está estrechamente unido al equilibrio). Lo encontramos en
peces. Está formado por el lóbulo flóculonodular y la língula (opuesta al lóbulo
flóculonodular). Los fisiólogos lo llaman vestíbulocerebelo porque une estas dos
estructuras. Estas fibras deben hacer sinapsis en un núcleo de sustancia blanca que es el
núcleo fastigial (es de sustancia gris, la cual está en sustancia blanca cerebelosa).
Paleocerebelo o Espinocerebelo
Es propio de cuadrúpedos. Además de las dos estructuras de antes, también el
lóbulo anterior del cuerpo del cerebelo y 4 segmentos de la vermis (lóbulo central,
culmen, úvula y pirámide). Para los fisiólogos se llama espinocerebelo porque aparte de
estar unido a vestibulares, está unido a la médula espinal. A este nuevo cerebelo llegan
aferencias propioceptivas de la medula espinal por las vías espinocerebelosas. Hay dos
núcleos: globoso y emboliforme. Con esta evolución se controla la postura y la marcha.
Neocerebelo o Pontocerebelo
Es sólo de mamíferos, ya que en estos animales controla la sintonización de
movimientos finos voluntarios.
381
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
En cuanto a volumen es la mayor parte del cerebelo. Aparece mucho después en
la evolución. Forma los hemisferios cerebelosos y partes del vermis y permite procesar
la información no sólo de la medula espinal y del sistema vestibular sino también desde
la corteza cerebral a través del puente por las vías corticocerebelosas. Así que aparecen
además de las vías cortococerebelosas, el tercero de los pedúnculos y el núcleo dentado
en la sustancia blanca.
Corteza
La corteza del cerebelo se estructura en tres capas, la molecular o plexiforme, la
ganglionar o de los somas de Purkinje, y la granulosa o de los granos. En esas tres capas
encontramos diferentes neuronas y células gliales, fibras y glomérulos.
Sustancia Gris
Es periférica y está organizada en tres capas:
x Capa Molecular o Plexiforme: No es
demasiado basófila a pesar de ser sustancia
gris. Es la más externa y relativamente
pobre en células. Contiene neuronas
horizontales de Cajal o estrelladas externas
y células en cesta o estrelladas internas.
Ambos tipos son inhibidoras.
x Capa Ganglionar o de los Somas de
Purkinje: Contiene las células de Purkinje.
Su axón es la única eferencia que sale de
este órgano. En esta imagen se ve arriba la
capa molecular y después unos somas
grandes, justo en la transición con la de más
abajo que es más basófila. Los somas no
están pegados entre sí, entre ellos queda un
espacio ocupado por glía.
x Capa Granulosa o de los Granos: Es muy rica en células. Las más abundantes
son los granos cerebelosos. También está la neurona Golgi II o estrelladas
grandes. Las Golgi son inhibidoras y los granos son las únicas excitadoras del
cerebelo.
Tipos neuronales
Células Horizontales
Están en la parte más superior de la capa plexiforme. Tienen un soma pequeño
estrellado con dendritas en todas direcciones y planos en las que no se aprecian muchas
espinas. Se calcula que cada célula de Cajal estaría sinaptando aproximadamente con
doce árboles dendríticos de Purkinje (doce neuronas de Purkinje). El axón de estas
células circula paralelo a la superficie y es relativamente corto y que, además de
382
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
sinaptar con las dendritas ya mencionadas, también dan colaterales que acaban en los
somas de Purkinje, y sus dendritas y axón se ramifican en un plano transversal.
Neuronas en Cesta
Son relativamente parecidas a las anteriores, pero se encuentran en la mitad
inferior de la capa molecular. Su soma es estrellado y algo más grande que las de Cajal,
y tanto él como el axón son paralelos a la superficie. El axón es bastante más largo que
el de las células de Cajal. Las dendritas salen de cualquier punto del soma en todas las
direcciones y no tienen muchas espinas, pero lo importante de estas neuronas es que las
colaterales del axón se dirigen y van a acabar ramificándose alrededor de los somas de
Purkinje (acaban abrazando el soma formando una especie de nido, cosa que no ocurre
en las horizontales), de manera que estas neuronas en cesta son capaces de sinaptar
tanto con el soma como con el cono axónico de estas neuronas de Purkinje. Las
dendritas y axones se ramifican un único plano transversal. Tanto las anteriores como
estas son inhibidoras. Así que la capa molecular se divide en dos estratos: el de las
horizontales y el de las de cesta.
Células de Purkinje
Tienen un soma con forma piriforme
muy grande (80μm) por lo que se las suele
conocer como células en botella o en pera. Se
disponen en una única hilera y están
ligeramente espaciadas entre ellas a
intervalos regulares.
El soma queda recubierto por una glia
especial que es la glía de Bergmann. Sólo
determinados puntos están sin cubrir por esta
glía y son aquellos donde se realiza la
sinapsis (con las ramas colateral de las
neuronas en cesta que recordemos que las
rodeaban), es decir, los botones sinápticos.
Su núcleo es grande, central y pálido con
mucha eucromatina, y tienen un nucléolo
marcado. La envuelta nuclear suele tener
profundas invaginaciones.
Son muy activas (citoplasma con abundantes orgánulos), tienen muchos
gránulos de Nissl, un citoesqueleto no muy marcado (neurofibrillas) en el soma aunque
sí en las dendritas y no son característicos los pigmentos de lipofucsina. Además, el
REL se acumula debajo de la membrana y constituye la formación hipolemnal o
submembranosa.
383
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
De estas células salen uno o dos troncos ascendentes (dendritas) en posición
apical y se van a ir ramificando de manera dicotómica. Todas las ramas que salen de ese
tronco dendrítico quedan en un único plano (cada una en uno), transversal, siendo la
única célula que hace esto. Estas ramas de las dendritas se extienden por toda la capa
molecular. Así, cada neurona de Purkinje tiene un árbol dendrítico muy denso que se
extiende por toda la capa plexiforme.
En las dendritas que tienen un nivel de ramificación muy superior encontramos
una superficie rugosa por su gran cantidad de espinas dendríticas. Esta cantidad de
espinas se deben a las sinapsis de las fibras paralelas, en las ramas finales. Se calcula
que una sola célula de Purkinje presenta más de 60.000 sinapsis espinosas y contacta
con 200.000 fibras paralelas.
Las dendritas se ramifican profusamente, y van adelgazando. Hay primarias u
secundarias. Entre ellas encontramos fibras que van a sinaptar. La fibra trepadora se
enrolla en cada soma del Purkinje, asciende sobre cada árbol dendrítico sobre todo por
las dendritas primarias y secundarias.
Frente al árbol dendrítico, preciosísimo, grandísimo y súper exuberante,
tenemos un axón pequeñajo, chiquitillo y cuando llega la sustancia blanca, se
mielinizará.
La sustancia gris da colaterales con dos destinos: volver a la capa de Purkinje y
sinaptar con Purkinje vecinas (luego ésta inhibe a las vecinas) o sinaptar con las
dendritas de Golgi tipo II.
Células de los Granos Cerebelosos
Son células con un soma muy pequeño (5-8μm) y redondo. El núcleo es muy
heterocromático y basófilo, denso y redondo y apenas presenta gránulos. Su citoplasma
es muy escaso y se ve como un fino halo alrededor del núcleo.
Tienen entre tres y cinco dendritas que se ramifican muy poco y terminan en
forma de garra. No se observan espinas. Su axón es muy largo y fino, y sale o de una de
las dendritas o del soma. Asciende hasta la capa molecular, donde se bifurca en T y pasa
a denominarse fibra paralela, de 3mm (es decir que estas fibras paralelas son el axón de
los granos que ya no es ascendente sino en capa molecular paralelo a la superficie).
Estas fibras son excitadoras y sinaptan con las dendritas de Purkinje y las neuronas de
la capa plexiforme (horizontales, células en cesta y ramas colaterales de Golgi II). Cada
fibra paralela, de los axones, es capaz de sinaptar con al menos 350 árboles dendríticos
de Purkinje, estimulándolos.
Células de Golgi II
Se sitúan en la capa de los granos, pero son minoritarias y están dispersas entre
las células de los granos, que las enmascaran. Las encontramos con paciencia, buscamos
un soma bastante más grande que el de los granos y el núcleo es bastante más pálido
con nucléolo (cosa que no tiene el grano) excéntrico.
384
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Su soma es mucho mayor, y hay una cierta tendencia a que se sitúen cerca de la
capa más externa, cerca de los somas de Purkinje (próximo a la capa ganglionar). Estas
neuronas presentan dendritas muy abundantes que se ramifican en todas direcciones
(tridimensionales). Estas dendritas ascienden hacia la capa molecular, ocupando buena
parte de ella y sobre ellas se producen sinapsis con las fibras colaterales del axón de las
células de Purkinje, con las fibras paralelas (el axón de los granos), con el axón de las
horizontales (inhibidas por el resto de neuronas), de las células en cesta y con los axones
de los granos. No llegan tan arriba como las fibras paralelas. Su axón es muy corto (no
sale de su capa) que se ramifica quedando muy cerca del soma y al final cada uno de
estos axones va a recorrer un trecho y forma sinaptar con un glomérulo.
Elementos Gliales
En sustancia gris, cerebelosa, en la capa molecular y granular son por una parte
microglía, por otra oligodendroglía en la capa granular acompañando a las fibras
trepadoras, musgosas y los axones de las de Purkinje. También astrocitos
protoplásmicos (astrogía protoplásmica) en la capa granular y dos glías características
de esta capa: las células de Bergmann o en horquilla se sitúan en la capa ganglionar
entre los somas de las células de Purkinje; y las células de Fañanás se encuentran en la
capa molecular.
Células de Bergmann
Son variantes de astrocitos cuyos somas se sitúan en la capa ganglionar entre los
de las células de Purkinje. Su primera función es dar sostén a las de Purkinje. Son una
horquilla ya que de su soma salen dos largas prolongaciones varicosas/tortuosas y
ramificadas, con perfil irregular, que ascienden y llegan a la superficie hasta las
meninges donde se aplanan formando la limitante glial subpial (porque recordemos que
entre las meninges y las neuronas siempre hay una limitante glial). En la superficie
forman la limitante glial del cerebelo. Cuando las células de Purkinje mueren su espacio
385
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
es ocupado por la glía de Bergmann, que prolifera abundantemente y ocupa los espacios
vacantes.
Células de Fañanás
Son específicas del cerebelo. Tienen prolongaciones mucho más cortas, que no
alcanzan nunca la superificie, con una ramificación en forma de pluma (aspecto
peniforme). Su soma se encuentra a nivel de las células de Purkinje (capa ganglionar) o
más arriba, en la capa molecular. Sus prolongaciones no alcanzan nunca la superficie ni
la limitante glial, pues aunque se adentran en la capa molecular, son muy cortas.
Fibras que encontramos en la sustancia gris
Hay dos tipos que son:
x Eferentes: Sólo sale el axón de las fibras de Purkinje. Este axón es inhibidor y se
dirige a un núcleo cerebeloso de la sustancia blanca. Por tanto, toda información
eferente del cerebelo es inhibidora.
x Aferentes: Excitadoras.
o Fibras trepadoras: Están alrededor del soma de Purkinje. Llegan desde la
oliva bulbar inferior y sus núcleos accesorios.
Se decusan y ascienden como una liana alrededor de las dendritas. Como
primarias y secundarias. Cuando llegan a la capa de los granos pierden la
mielina y continúan ascendiendo.
Cada fibra trepadora va a un único soma, se enrolla alrededor del soma y
luego alrededor de las dendritas. Luego sus colaterales sinaptan con
axones de las horizontales y en cesta alcanzan los somas de las de
Purkinje (en principio para cada trepadora tenemos una de Purkinje),
momento en el que pierden la mielina. Se enrollan alrededor del árbol
dendrítico ascendiendo y sinaptan con las dendritas más iniciales.
o Fibras musgosas: Estas fibras nerviosas proceden de la médula espinal, el
bulbo raquídeo y el puente. Van a perder la mielina en la capa granulosa
y se ramifican.
Forman rosáceas que sinaptan en el glomérulo
cerebeloso. El glomérulo cerebeloso es un grupo
eosinófilo en la capa granulosa sin gránulos (no
confundir con Golgi tipo II, no es una célula sino un
espacio).
El glomérulo tiene una envuelta glial. En su centro están
las rosáceas, engrosamientos de las fibras musgosas con
muchas vesículas sinápticas y mitocondrias. Además
dentro del glomérulo se distinguen dendritas de granos
(garras) y axones de Golgi II.
Las rosáceas sinaptan con las dendritas de los granos,
estimulándolas. Los axones Golgi sinaptan con las
dendritas de los granos, inhibiéndolos.
386
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Sustancia blanca
x Fibras: trepadoras, musgosas y eferentes.
x Núcleos: Estaciones de relevo para aferencias y eferencias. Son 4: fastigiales,
globosos, emboliformes y dentados.
Núcleo fastigial
Cerca de la línea media. Recibe fibras de la corteza del vermis, los núcleos
vestibulares y la oliva inferior. Envía fibras a los núcleos vestibulares y de la médula
oblongada.
Núcleo globoso
Recibe fibras de la corteza del vermis y envía fibras a los núcleos de la médula
oblongada.
Núcleo emboliforme
Se sitúa junto al núcleo dentado. Recibe fibras de la corteza cerebelosa y envía
fibras al tálamo.
Núcleo dentado
Cordón plegado abierto que recibe fibras de la corteza cerebelosa y las envía a
tálamo y núcleo rojo.
Células
Contienen neuronas estrelladas de axón largo y glía (microglía, oligodendroglía
y astrocitos fibrosos).
Fibras
x Pedúnculo cerebeloso inferior:
o Fibras del tracto espinocerebeloso posterior.
o Fibras del tracto cuneocerebeloso.
o Fibras del tracto olivocerebeloso.
o Fibras del tracto reticulocerebeloso.
o Fibras del tracto vestíbulocerebeloso.
387
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
o Fibras del tracto arqueocerebeloso.
o Fibras del tracto trigéminocerebeloso.
Estas fibras intervienen en el control de los movimientos oculares y el equilibrio.
x Pedúnculo cerebeloso medio.
o Fibras del tracto pontocerebeloso posterior.
x Pedúnculo cerebeloso superior.
o Fibras del tracto espinocerebeloso anterior.
o Fibras del tracto tectocerebeloso.
o Fibras del tracto vestibulocerebeloso.
o Fibras del tracto cerulocerebeloso.
Participa en el control de la postura y la locomoción, además del aprendizaje
motor.
Funciones
x Coordinación de movimientos cada hemisferio controla los movimientos de su
propio lado).
x Coordinación de la actividad muscular, mantenimiento del tono muscular y
conservación del equilibrio.
Circuitos
Las fibras trepadoras y musgosas, estas últimas a través de los granos, envían
señales activadoras a las células de Purkinje. Ambas fibras también actúan sobre las
células horizontales y en cesta, que inhiben a las Purkinje. Las Purkinje mandan
colaterales a las vecinas, inhibiéndolas, y a las Golgi tipo II, que inhiben a las dendritas
de los granos en el glomérulo.
Las fibras trepadoras y musgosas activan a las neuronas de Purkinje, que inhiben
a los núcleos en la sustancia blanca. Además, las fibras trepadoras y musgosas también
están excitando los núcleos de sustancia blanca. De esta forma, la información que llega
al cerebelo excita primero a los núcleos de sustancia blanca, que después son inhibidos
por las células de Purkinje, actúan de moduladores controlando la información que
pueden mandar los núcleos.
Los núcleos de sustancia blanca están siendo excitados de modo constante por
colaterales de las aferentes (estímulo tónico constante). Los impulsos no pueden ser
transmitidos porque están bajo control inhibidor de las Purkinje.
388
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Las células de Purkinje son inhibidas y se suprime el estado de freno, saliendo
información desde los núcleos. Los estímulos que llegan a los núcleos de sustancia
blanca se transmiten. La corteza cerebelosa regula la transmisión de impulsos por parte
de los núcleos cerebelosos mediante inhibiciones y desinhibiciones muy bien
coordinadas.
La excitación de una fila de Purkinjes se acompaña de la inhibición de todas las
Purkinjes vecinas, produciéndose una delimitación del estímulo o contraste.
La excitación de los núcleos de la sustancia blanca es modulada por la inhibición
selectiva de unas zonas y la excitación de otras como resultado del procesamiento de la
información por la corteza cerebelosa.
Por tanto, los núcleos cerebelosos descargan un patrón organizado de impulsos
que actúan sobre vías motoras pudiendo:
x Coordinar músculos antagonistas y sinergistas de movimientos involuntarios.
x Mantener el equilibrio y la postura.
x Ser responsable de movimientos finos y voluntarios.
Clínica
Malformación de Arnold-Chiari
El cerebelo se mete en el conducto vertebral en el desarrollo embrionario al
crecer un tumor o un hematoma, obstruyéndose los agujeros de Luscka y Magendie. La
cabeza es enorme y los hemisferios cerebrales muy finos.
Lesiones en la línea media
Se producen meduloblastomas en el techo del 4ª ventrículo. Aumento de presión
intracreaneal, cefaleas, vómitos, somnolecia... Además se produce ataxia troncal,
incapacidad de estas de pie sin apoyo.
389
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Lesiones en el lóbulo anterior (alcohólicos crónicos)
Ataxia postural y de la marcha, se produce tambaleo incluso sobrios y con los
ojos abiertos. Diminución en un 10% de los granos, un 20% de Purkinjes y un 30% de
la capa molecular.
Lesiones neocerebelosas
Se produce una descoordinación de movimientos voluntarios, sobre todo en
miembro superior. Hay temblor intencional o de acción. Se pierde la trayectoria suave
de los movimientos. Los movimientos rápidos alternantes se hacen torpes incluso con
los ojos abiertos. La fonación es temblorosa y hay mala articulación (descoordinación
de nervios de labios, lengua, mandíbula y paladar).
Síndrome afectivo cognitivo cerebeloso
Hay importantes déficits cognitivos (parece que es en el cerebro pero es en el
cerebelo), menor capacidad de razonamiento, desatención, falta de sentido espacial,
errores gramaticales en el habla y falta de afectividad. Déficits funcionales del cerebro
debido a lesión cerebelosa.
CEREBRO
Órgano complejo que agrupa formaciones procedentes de 2 vesículas
encefálicas. Los hemisferios cerebrales derivan del telencéfalo; pos su parte el
epitálamo, el tálamo y el hipotálamo derivan del diencéfalo.
Su peso está entre 1250 y 1600 g, con función directa del peso corporal. La
media de una persona está en 1350g si es varón y 1250g si es mujer. En los ancianos se
ve reducido por atrofia. Cabe destacar que no hay correlación alguna con el nivel de
inteligencia y el peso del cerebro.
La sustancia gris es periférica y la blanca central, con núcleos grises. Sufre una
enorme expansión en el desarrollo, junto con el paso de liséncefalo a girencéfalo (surcos
y circunvoluciones). La superficie aumenta hasta 2600 cm2. Solo 1/3 de corteza está en
la superficie.
La expansión no es uniforme en el desarrollo, sino mayor en sentido caudal y
basal. Ciertas estructuras cambian de posición, debido a un movimiento circular. El
putamen y la ínsula no giran. Las estructuras adquieren forma de arco y se modifican las
posiciones relativas.
En el telencéfalo embrionario encontramos:
x Cuerpo estriado (suelo engrosado): De él derivarán los núcleos o ganglios
basales.
390
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Palio (techo fino y arqueado): De él derivará la corteza cerebral.
Cuerpo estriado
Centro sináptico superior subcortical del sistema motor extrapiramidal. Dividido
en núcleo caudado y núcleo del putamen.
La necrosis de las neuronas pequeñas del núcleo estriado produce Baile de San
Vito.
Palio
Según el grado evolutivo tenemos: paleopalio, arquipalio y neopalio o neocórtex
(en él aparecen surcos y circunvoluciones).
Paleopalio
Es la parte más antigua, por lo que en mamíferos primitivos constituye la mayor
parte de los hemisferios cerebrales. Conforma la base del hemisferio. En el hombre el
paleopalio queda desplazado a la profundidad, sobre él se desarrolla el cuerpo estriado.
Contiene áreas olfativas. El rinencéfalo está formado por el paleopalio junto al
bulbo olfatorio y los tractos olfativos. Nuestra especie ha perdido gran parte del sentido
del olfato, por lo que somos animales microsmáticos (bulbo olfatorio involutivo.
Se distinguen 4 capas: glomerular, plexiforme, mitral y granulosa. Hay 4 tipos
neuronales distintos: mitrales, empenechadas, periglomerulares y granos (amacrinas).
Contiene glomérulos olfativos con axones de neuronas olfatorios que sinaptan
con dendritas de mitrales y empenechadas y periglomerulares, activándolas. Además,
hay sinapsis dendrodendríticas entre las mitrales y empechadas con respecto a las
periglomerulares. Las dos primeras activan a las segundas, y éstas inhiben a las
primeras.
Hay sinapsis de aferentes vías proyecciones centrífugas procedentes de distintas
localizaciones, como el locus ceruleus, los núcleos del rafe…
391
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Las neuronas periglomerulares tienen axón corto y conectan glomérulos entre sí
(5), con dendritas arborescentes en glomérulo. Las mitrales y empanachadas son
eferentes. Los granos tienen sinapsis dendrodendríticas sobre mitrales y empenachadas
con efecto inhibitorio, carecen de axón y son un tipo de interneurona. Además
encontramos glía formando una cápsula periglomerular.
Archipalio
Parte medial de cada hemisferio cerebral. En el desarrollo la rotación obliga al
archipalio a enrollarse, formándose de esta forma el asta de Ammón. Constituye el
392
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
hipocampo y la corteza parahipocampal. Recibe aferencias ópticas, acústicas, táctiles,
viscerales y olfativas.
Es un órgano de integración en conexión con el hipotálamo, los núcleos septales
y el giro del cíngulo. Influye en la vida emocional y en las funciones endocrinas y
viscerales, además de encargarse del aprendizaje y consolidación de la memoria
reciente.
La resección bilateral del hipocampo en el tratamiento de epilepsias graves
provoca la pérdida de memoria. Los recuerdos antiguos se conservan pero los sucesos
recientes solo se recuerdan unos segundos.
Neopalio (continuación)
Cuando se empieza a desarrollar empuja al paleopalio y al arquipalio hacia
posiciones más basales. Finalmente, en algunos animales como los mamíferos y, por
ende, el ser humano, aparecen una serie de plegamientos que dan lugar a los surcos y
giros de los que dispone el neopalio. Pese a que ocupa la mayor parte de los hemisferios
cerebrales, tan solo vemos a simple vista 1/3 de la superficie del neopalio, puesto que
los 2/3 restantes quedan ocultos en zonas más profundas.
Estructura del palio
Isocórtex
Presenta un máximo desarrollo, de manera que en él distinguimos seis capas. Se
corresponde con las regiones evolutivamente más modernas, que constituyen el
neocórtex. Sin embargo, existen variaciones zonales del neocórtex, que han permitido
delimitar mapas de áreas corticales, el más famoso el de Brodmann en los años 50 del
siglo pasado.
A grandes rasgos, un isocórtex puede ser:
x Homotípico: Las 6 capas que distinguimos presentan igual grado de desarrollo.
x Heterotípico: Una de las capas adquiere más predominancia. Según la que sea, lo
clasificamos en:
o Coniocórtex o córtex heterotípico granular: Es el propio de las áreas
sensitivas y, especialmente, de la acústica (41 y 42 de Brodmann). Como
su propio nombre indica, en él predominan las capas granulares. Se trata
de áreas en las que finalizan numerosas aferencias, y que por ello
disponen de neuronas de axones cortos, las cuales participan en la
asociación y la elaboración de la información.
o Córtex heterotípico agranular o corteza motora: Es propio de las áreas 4
y 6, motoras. En él hay un mayor desarrollo de las capas piramidales,
cuyas neuronas tienen largos axones.
393
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Allocórtex
No distinguimos las 6 capas, sino que por lo general vemos 3 o 4. Comprende
las regiones de arquipalio y paleopalio, más antiguas, pero el hecho de que sean
histológicamente menos complejas no implica nada, sigue tratándose de regiones con
una alta especialización.
Para analizar las capas del palio, lo más lógico es emplear un isocórtex
homotípico, pues presenta un desarrollo equitativo de todas las capas. Sin embargo, la
apariencia de las mismas depende del tipo de tinción que empleemos.
Así, con impregnación argéntica y tinción de Nissl, que es la técnica con la que
se suelen describir los estratos, estos son:
x I, molecular, plexiforme o marginal
x II, granular o granulosa externa
x III, piramidal o piramidal externa
x IV, granular o granulosa interna
x V, ganglionar o piramidal interna
x VI, polimorfa o multiforme
Con tinción de mielina, las clasificamos de un modo distinto:
x I, tangencial
x II, disfibrosa
x III, supraestriada
x IV, estría de Baillarger externa
x V, estría de Baillarger interna
x VI, subestriada
Además, con esta tinción hallamos fibras verticales, denominadas radiales.
394
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Entre unas capas y otras hay zonas limítrofes,
que son las estrías:
x De Kaes-Bechterew: entre II y III.
x De Baillarger externa: entre IV y V.
x De Baillarger interna: entre V y VI.
Células del isocórtex
Las células del isocórtex son las gliales típicas (astrocitos protoplásmicas,
oligodendroglía y microglía) y neuronas Golgi de tipos I (axón largo) y II (axón corto).
Neuronas Golgi de tipo I
Son neuronas piramidales. Constituyen el tipo celular principal del isocórtex, y
se encuentran en todas las capas menos en la I, siendo por tanto mayoritarias y
constituyendo un 70% del isocórtex. Son las únicas neuronas eferentes, y se trata de
células excitadores.
x Soma: Su soma es piramidal. En las capas II y IV son pequeños, en la III
medianos y grandes, y en la V hallamos las neuronas gigantopiramidales de
Betz. En la VI son irregulares.
x Dendritas: Existen dos tipos de dendritas, ambos con numerosas espinas:
o Basilares
o De tallo apical: Es un tallo ascendente que, a medida que se dirige hacia
arriba, da ramas oblicuas, y al llegar a la capa I se divide en ramas
horizontales.
x Axón: Es largo. Sale de la parte basal del soma o de las dendritas basilares, y se
dirige descendientemente hacia la sustancia blanca. En su camino, emite ramas
colaterales recurrentes que sinaptan con neuronas Golgi de tipo II.
Neuronas Golgi de tipo II
Existen 6 tipos de neuronas Golgi de tipo II. Todas, excepto las estrelladas
espinosas, son inhibidoras y presentan pocas espinas. Su función es modular la acción
de las células piramidales mediante sinapsis con sus colaterales recurrentes (como
hemos dicho, en su mayoría inhibitorias).
Células horizontales
Se encuentran en la capa I. Su soma es fusiforme y paralelo a la superficie de la
capa I. Sus dendritas se sitúan en el plano horizontal, y se agrupan en dos ramilletes
dendríticos en los extremos del soma. Su axón también se encuentra en el mismo plano,
y tras salir de la parte basal se bifurca en forma de T. Estas prolongaciones emiten
colaterales ascendentes.
Este tipo de células sinapta de manera inhibitoria con células piramidales.
395
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Células bipenachadas
Se encuentran en las capas II, III y IV. Su soma es fusiforme y perpendicular a la
superficie de la capa I. Sus dendritas se sitúan en el plano vertical, y se agrupan en dos
ramilletes dendríticos en los extremos del soma. Su axón también se encuentra en el
mismo plano, y tras salir de la cara lateral se bifurca en forma de T. Estas
prolongaciones emiten colaterales ascendentes.
Este tipo de células sinapta también de manera inhibitoria con células
piramidales.
Células en candelabro
Se encuentran en las capas II, III, IV y V, pero son especialmente abundantes en
las dos primeras. Su soma es estrellado, y tienen muchas dendritas. Su axón es
descendente y también se bifurca en T, pero emiten ramas ascendentes que también se
bifurcan.
Estas neuronas sinaptan con el cono axónico de las piramidales, inhibiéndolas.
Células en cesta
Se hallan en las capas III y V. Su soma es estrellado, y tienen muchas dendritas.
Su axón es descendente y emite ramas horizontales.
Células de Martinotti.
Se encuentran en las capas III, IV, V y VI. Su soma es estrellado, y tienen
muchas dendritas. Su axón asciende hasta la capa I, y sinaptan con el tallo dendrítico
apical de las células piramidales.
Células estrelladas espinosas
Se encuentran en la capa IV y en áreas sensoriales y, como su propio nombre
indica, tienen el soma estrellado. Como ya hemos dicho, tienen muchas dendritas y son
las únicas neuronas Golgi II excitatorias.
396
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Fibras del isocórtex
Pese a que analicemos la corteza por capas, los tipos celulares se organizan
funcionalmente formando columnas verticales. Hay 4 millones de columnas, unas 2500
neuronas por columna y de ellas 100 piramidales.
Mountcastle fue quien delimitó estas unidades funcionales, que no son visibles a
simple vista y que seguramente dependan de los distintos individuos y el momento vital
de los mismos, no teniendo una localización fija. A ellas llegan aferencias como fibras
de asociación o como aferentes sensoriales de la periferia, y distribuyen eferencias
(recordemos que las únicas son los axones de las células piramidales) que terminan
sobre piramidales de otras columnas o en áreas de asociación:
x Aferencias como fibras de asociación: Proceden de otras columnas, y dentro de
una misma columna ofrecen ramas a todas las capas, acabando finalmente
sinaptando con las espinas de piramidales.
x Aferencias como fibras sensitivas específicas: Acaban en las células de la capa
IV, especialmente en las horizontales y bipenachadas. Establecen cadenas de
sinapsis con piramidales.
De proyección (con corona radiada)
Sirven para conectar la corteza cerebral con centros subcorticales y pueden ser:
x Eferentes: Son excitadoras, y según la capa de la que salgan tienen destinos
diferentes:
o V: Van a la médula espinal, el cuerpo estriado, el tronco del encéfalo y el
cerebelo, pasando previamente por la protuberancia.
o VI: Van al tálamo.
Las eferentes de distintas áreas se acaban uniendo para formar la cápsula interna.
x Aferencias:
o Tálamocorticales: Se trata de aferencias excitadoras procedentes del
tálamo que acaban en su mayoría en la capa IV y, en menor medida, en
la V y la VI.
o Extratalámicas: Son moduladoras, y emplean diferentes
neurotransmisores según su lugar de procedencia. Así, las del locus
ceruleus usan noradrenalina, las de los núcleos del rafe serotonina, las de
la sustancia negra dopamina y las del núcleo basal de Meynert Ach.
De asociación
Conectan distintas áreas de un mismo hemisferio, y pueden ser:
x Cortas: arqueadas o arcuatas. Tiene forma de U, y conectan regiones situadas en
el mismo lóbulo, o giros adyacentes.
x Largas: Conectan lóbulos, formando haces que se pueden distinguir a simple
vista.
397
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Fibras comisurales
Conectan las cortezas de los distintos hemisferios. La comisura principal es la
que constituye el cuerpo calloso, pero también están la comisura anterior, la posterior y
el fórnix. Si conectan las mismas áreas de los dos hemisferios se denominan
homotípicas, si conectan distintas áreas de los mismos se llaman heterotípicas.
En caso de seccionamiento del cuerpo calloso, aparece una patología que se
denomina cerebro hendido. No conlleva alteraciones de la personalidad ni de la
inteligencia, con lo que permite al paciente llevar una vida normal, pese a que presenta
pequeñas deficiencias táctiles y visuales.
Hemos de recordar que la corteza es el punto final de las vías sensoriales, donde
percibimos el estímulo, es decir, es el sustrato anatómico de la conciencia, de manera
que si existiese una interrupción o desconexión entre el córtex y los centros inferiores se
pierde esta conciencia. A esto se le denomina síndrome aplásico, y la persona que lo
padece está despierta pero no es consciente de los estímulos.
En las distintas áreas corticales reside también una capacidad exclusivamente
humana, que es la del lenguaje. Distinguimos:
x Lenguaje interno: Es el pensamiento.
x Lenguaje hablado: Es la comunicación verbal.
x Lenguaje escrito: Permite transmitir la información mediante un tipo de
comunicación que perdura en el tiempo.
Las áreas corticales del lenguaje se integran, y se localizan en el hemisferio
dominante, de manera que:
x Diestros: Su hemisferio dominante suele ser el izquierdo.
x Zurdos: Su hemisferio dominante puede ser cualquiera o los dos.
Aun así, esto no es ciencia exacta, que sea diestro no significa necesariamente
que domine tu hemisferio izquierdo. También hemos de destacar que el hemisferio
derecho se relaciona con el lenguaje musical. Las principales regiones hemisféricas que
participan en el lenguaje hablado y escrito son, en cualquier caso:
x Centro de Wernicke: Se sitúa en la zona posterior del giro temporal superior. Se
trata de un centro integrador que sirve para interpretar el lenguaje cuando lo
escuchamos, y para conservar patrones verbales aprendidos. Su lesión conlleva
afasia, es decir, no permite entender el lenguaje ni expresarse con sentido.
x Circunvolución angular: Es un centro integrador que sirve para conservar
patrones escritos aprendidos. Su lesión conlleva agrafia, es decir, no permite
expresarse con sentido por escrito, y alexia, esto es, incapacidad de leer.
x Campo de Broca: Coordina los músculos del habla con lo que queremos decir.
Si falla, tenemos una afasia motora sin parálisis de los músculos que intervienen
en la fonación. Aun así, el paciente puede comprender y expresarse por escrito.
x Circunvolución supramarginal.
398
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Cuando hay lesión en el derecho, lo que originan son síndromes aprosodias.
Cuando se daña el área de Broca la persona se sigue expresando correctamente, pero de
manera fría, monótona, sin emoción. Si se daña el centro de Wernicke hay deficiencias
en escucha y habla; cuando se le habla, no detecta en qué contexto se le habla (ironías,
sarcasmo, segundas intenciones), no sabe la forma del lenguaje utilizada…
PATOLOGÍAS
Lóbulo Parietal
Presenta asimetrías. En el hemisferio derecho está relacionado con la apreciación
de las formas, la localización de la persona en el espacio y los objetos de alrededor,
objetos a través del tacto y regulación de la personalidad. Este mismo lóbulo, pero en el
hemisferio izquierdo está relacionado con el movimiento, reconocimiento numérico,
tareas de escritura…
Si se daña el lóbulo en el derecho aparece la Agnosia Táctil Apreciativa. Cuando
la persona busca las llaves, es incapaz de encontrarlas porque no se reconocen al tacto.
El paciente no podrá distinguir con la mano izquierda los objetos que no ve
(astereognosia)
Si se daña en el hemisferio izquierdo, se produce Agnosia Táctil Asociativa. El
paciente puede distinguir los objetos, pero tiene dificultades para enunciar la función de
los mismos. Mete la mano y saca las llaves, pero no sabe para qué sirven.
Una parte de este lóbulo está relacionado con el giro supramarginal. A la lesión
en este giro o área 40, se le llama Heminegligencia Personal. Ignora la persona el lado
contralateral. Si tiene dañado el lóbulo en un hemisferio, el lado contrario es como si no
existiera.
Corteza Prefrontal
Presenta conexiones con todo el neo-córtex, excepto en las áreas motoras y
sensitivas primarias.
Se ocupa de funciones cerebrales superiores, pensamiento abstracto, toma de
decisiones, anticipación de efectos y conducta social.
El hemisferio izquierdo desarrolla funciones como la conversación interna y el
pensamiento. Por otro lado, el hemisferio derecho se activa en el contexto del miedo
(nuestra especie es la única que puede pasar miedo en una situación que no es de
miedo).
La zona dorso-lateral, está implicada en el aprendizaje consciente (supervisor de
vigilancia). La zona media, con asociaciones auditivas y verbales.
399
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Las lesiones del lóbulo frontal producen cambios sutiles en la personalidad y
disminución del rendimiento cognitivo: falta de previsión, distracción y dificultades en
conmutar en procesos cognitivos. En la marcha se producen pasos cortos, arrastrando
los pies y con tendencia a caer.
x Si afecta al lóbulo derecho lateral se produce enlentecimiento del proceso
mental, apatía e indiferencia e hipocinesia
x Si afecta al orbito-frontal se produce conducta infantil, hipercinesia con aumento
de impulsos instintivos.
Enfermedades Neurodegenerativas
Se caracterizan porque en la sustancia gris del encéfalo hay una pérdida
progresiva de neuronas y puede ir acompañada de cambios en la sustancia blanca.
La pérdida de neuronas no es aleatoria, es muy selectiva. Se observa una
presencia de agregados proteicos que son resistentes a la degradación a través del
sistema ubicuitina-proteasoma. La causa de la agregación es variable. Además, las
inclusiones citoplasmáticas constituyen marcadores diagnósticos. Se clasifican según
criterios sintomáticos/anatómicos o anatomopatológico.
Enfermedades en la Corteza Cerebral
Demencia es la pérdida grave de la función cognitiva sin que la persona deje de
estar consciente. La demencia hace referencia en cuestión a un concepto patológico, no
a la senectud. En muchas enfermedades la demencia es precoz. La más conocida es la
de Alzheimer.
Alzhéimer
Aumenta su incidencia en la población envejecida. Consiste en un deterioro
cognitivo de las funciones superiores, en el ánimo, conducta y personalidad. También se
ve afectada la memoria y las personas que padecen esta enfermedad se desorientan.
Sufren afasia progresiva. El avance de la enfermedad es distinto en cada persona. En 5-
10 años puede aparecer profundamente discapacitado inmóvil y mudo.
El diagnóstico definitivo requiere estudio anatomopatológico. Una persona con
Alzheimer presenta:
x Grave atrofia de la corteza cerebral (lóbulos frontal, temporal y parietal).
x Ensanchamiento de surcos.
x Engrosamiento del sistema ventricular.
x Perdida de parénquima.
x Reducción del volumen encefálico.
x La parte medial del lóbulo temporal es la más afectada.
x Pérdida masiva de piramidales.
x Pérdida de neuronas colinérgicas del núcleo basal de Meynert y área septal.
400
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Mientras, en el cuerpo amigdalino, en el hipocampo y en el neo-córtex aparecen:
x Placas neuríticas, seniles, amiloide. Son depósitos esféricos focales de
prolongaciones neuríticas dilatadas y tortuosas alrededor de un núcleo central
amiloide, rojo Congo+, rodeados de un halo claro. Alrededor hay gliosis
reactiva.
El Núcleo amiloide contiene A-beta, componentes del complemento, citocinas
proinflamatorias, apolipoproteínas y alfa 1-antiquimotripsina. Cuando hay
depósitos de amiloide sin reacción neuríticas se habla de placas difusas, parecen
un estadio precoz. Son más abundantes en el hipocampo y en neo córtex.
Los depósitos de A-beta se deben al mal procesamiento de la APP (proteína
transmembranal), Rc transmembrana. Esta proteína es escindida por unas
encimas que son secretasas alfa beta y gamma.
La secretasa alfa escinde la porción extracelular y hay dos posibilidades: la alfa
secretasa, que tenemos una ruta que no produce el amiloide, es la vía no
amiloidogénica, mientras que cuando activa la beta secretasa el punto de
escisión es diferente.
Hay una segunda escisión intramembranal. Como el primer punto de sección es
distinto en la vía amiloidogénica se produce un péptido diferente con
propiedades de agregarse. Inicialmente lo que se forman son pequeños
agregados con propiedades neurotóxicas que dañan la neurona y luego forman
grandes placas.
También tienen la apolipoproteínas E. Esta codificada por el gen de las
apolipoproteínas E del cromosoma 19 tiene tres alelos épsilon 2 3 y 4 según
polimorfos de tres aminoácidos. La dosificación de épsilon 4 incrementa el
riesgo de EA y disminuye la edad de inicio.
x Ovillos neurofibrilares: Se deben a la alteración de la proteína Tau, que está
hiperfosforilada. Esta proteína se relaciona con los microtúbulos y el transporte
axónico.
El caso es que la aparición de las placas parece previa y con posterioridad la
proteína TAU deja de estar asociada a los microtúbulos en los axones, va a
desplazarse hacia el soma y dendritas.
Se hace insoluble y con ello carece de afinidad por los microtúbulos, que no
están estabilizados, y por tanto afecta al transporte axónico. Además, la proteína
Tau comienza a agregarse formando filamentos helicoidales. Hay relación entre
el número de ovillos que aparecen y el grado de demencia. Una mutación en Tau
provoca demencia fronto-temporal.
x Degeneración granulo vacuolar: Formación de vacuolas citoplasmáticas claras
con un granulo argirófilo.
x Cuerpos de Hirano: Cuerpos eosinófilos vítreos alargados para cristalinos
formados por actina. Están en las neuronas.
401
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Todos estos cambios provocan alteración en la función del hipocampo, lo que
produce amnesia retrógrada y déficit de atención. Se produce una pérdida de memoria
pasada muy marcada.
Además, hay una disminución de la función occipito-temporal (afecta diferente
según el paciente): si se lesiona el área 37 incapacidades para leer y escribir, si se afecta
la temporal polar incapacidad para reconocer familiares y si es el gris supra marginal
incapacidad para escribir. Si se afecta el lóbulo frontal el paciente está ausente
aparentemente sin enterarse. En algunos pacientes, las lesiones van con no poder
reconocer caras familiares y la persona se mete en su mundo particular, perdiendo
conexiones con el resto del mundo.
Alzheimer Familiar
Es la instauración precoz (40-50años). Mutaciones puntuales (puntos de corte de
las secretasas aumento de la propensión a la agregación etc.) y duplicaciones (efecto de
dosificación) den gen APP del cromosoma 21 (cerca de la región Down, por lo que los
Down cuando llegan pueden tener síntomas). Cuantas más largas son las duplicaciones
antes aparece la enfermedad. .
402
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Demencia fronto-temporal.
x Enfermedad vascular.
x Demencia con cuerpo de Lewy.
x Neurosífilis.
x Enfermedad de Creutzfeldt-jacobs.
Enfermedades Neurodegenerativas de Ganglios y Tronco Encefálico
Disminución de la expresión facial, postura encorvada, enlentecimiento del
movimiento voluntario, mayor rigidez en la cabeza y temblor de pildorero. Esto son
lesiones en el sistema dopaminérgico nigro-estriata.
Enfermedad de Parkinson
Se debe a la pérdida masiva de neuronas del sistema dopaminérgico. Antes de
que aparezcan los síntomas, la persona que va a sufrir la enfermedad muestra aparición
en las neuronas de las neuritas de Lewy (neurofilamentos típicos) y unos depósitos de
alfa sinucleina (cuerpos de Lewy). Como consecuencia de estas alteraciones lo que se
produce es una pérdida de pigmentación de sustancia negra y locus ceruleus y muerte de
un número importante de neuronas nígricas y reacción de la glía.
La falta de estas neuronas lleva a un descenso en los niveles de dopamina, lo que
conlleva una neuropatía por degeneración retrógrada.
En esta enfermedad, los pacientes presentan el siguiente cuadro clínico:
x Temblores en los miembros, primero superior después inferior homolateral y a
los 3 años afecta al lado contrario.
x Es un temblor característico y se llama temblor de reposo porque solo afecta a
los músculos que están en reposo y desaparece en los movimientos voluntarios.
x Ligero temblor de acción que se manifiesta en escritura temblorosa.
x Temblor de lengua y labios.
x Movimiento característico de índice y medio sobre pulgar.
x Rigidez muscular progresiva sobre todo de flexores.
x Caminar peculiar.
x Mayor dificultad para hablar, escribir (cuesta más, la letra es más pequeña e
irregular y los movimientos son más lentos y torpes debido a la rigidez
muscular).
La mayor parte tienen herencia autosómica dominante. Hay dos genes descritos
la cinasa2 y la alfa sinucleina (implicada en botones sinápticos). En ambos casos hay un
aumento en el número de copias del gen. Pero no todas las personas con Parkinson
presentan alteraciones en estos genes.
El gen LRRK2 al expandirse, causa un mayor riesgo de Parkinson y una
aparición del mismo más temprana.
403
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Se han descritos casos de herencia autosómica recesiva que es el Parkinsonismo
Juvenil. Varios genes, uno implicado en la función mitocondrial, pero el más conocido
es el de la Parkina, una ubiquitina que parece que cuando está mutada lo que hace es
afectar a la eliminación, a la metabolización de la alfa sinucleina.
Otros genes implicados son el DJ-1 y PNK1.
Corea de Huntington
Es una enfermedad neurodegenerativa hereditaria. Lo que tenemos es la
afectación y muerte de las neuronas del estriadas, primero el caudado y luego el
putamen, mueren primero las pequeñas y luego las más grandes.
Las personas con Huntington presentan trastornos a nivel cognitivo, tienen
movimientos espasmódicos y llamativos por el cuerpo. Tienen trastornos en la corteza
que producen trastornos afectivos de conducta y son cohibidos. Presentan una pérdida
importante de memoria y se ve afectada la parte afectiva de la personalidad.
Los pacientes muestran expansión del gen HD (repeticiones CAG) que es la
huntingtina (región de la poliglutamina) interviene en el transporte vesicular y mediado
por microtúbulos. Más de 35 repeticiones producen la enfermedad de Huntington. Hay
una relación inversa entre el número de repeticiones y la edad de inicio. Expansiones en
espermatogénesis, anticipación.
Cuando aparecen las repeticiones aumentan determinadas enzimas
fundamentalmente la caspasa 1 y esto activa a la 3 que se dedican a eliminar la
huntingtina salvaje. Esto activa las vías apoptóticas a través de una familia de proteínas
que es la Bid (familia BCL2) lleva a la formación y liberación masiva del citocromo c y
se une a apaf-1 y se forma el apoptosoma. Esto activa a la caspasa 9 lo que provoca
muerte neuronal.
404
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
GLÁNDULA PITUITARIA O HIPÓFISIS
Es la principal glándula endocrina en conexión con el hipotálamo por el tallo
hipofisario y red vascular. Está alojada en la silla turca, una depresión del esfenoides.
Pesa 0,5 gramos en varones y 1,5 en mujeres multíparas. Entre el periostio y la cápsula
hay un plexo venoso.
Los órganos endocrinos son: hipófisis, glándula pineal, tiroides, paratiroides y
glándulas suprarrenales.
El hipotálamo y la hipófisis (denominada también pituitaria) forman una red
neuroendocrina integrada que se llama sistema hipotalamohipofisario. El sistema
hipotalamohipofisario comprende dos componentes: 1) el sistema hipotalámico
adenohipofisario, que conecta el hipotálamo con la hipófisis anterior, y 2) el sistema
hipotalámico neurohipofisario, que une el hipotálamo con la hipófisis posterior.
El hipotálamo, que corresponde al suelo del diencéfalo y que forma parte de las
paredes del tercer ventrículo, está constituido por agregados de neuronas, llamados
núcleos, algunos de los cuales secretan hormonas. Estas células neuroendocrinas se
localizan por detrás de la barrera hematoencefálica, pero sus productos de secreción se
liberan al exterior de la misma.
ESTRUCTURA
Anatómica
Se divide en dos lóbulos:
x Anterior o pars distales: Contiene cordones celulares ramificados, folículos
ocasionales y una red de sinusoides.
x Posterior, que incluye pars intermedia y pars nervosa.
x El infundíbulo conecta con el hipotálamo.
Histológica
La adenohipófisis es grande,
glandular y rojiza, de carácter
epitelial; incluye la pars distalis, la
pars intermedia y a pars tuberalis.
La nurohipófisis es pequeña,
blanca y firme, de carácter neural;
incluye la pars nervosa, el tallo o eje
infundibular o neural y la eminencia
media.
405
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Neurohipófisis, tallo neural: Eje del tallo hipofisiaria.
x Adenohipófisis, pars tuberalis: Lengüeta que rodea el tallo neural. No secretora
x Adenohipófisis, pars intermedia: Banda por delante de la pars nervosa.
ORIGEN EMBRIOLÓGICO
406
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Su origen es doble: la adenohipófisis deriva de la bolsa de Rathke, una
evaginación del techo del estomodeo (ectodermo), y la neurohipófisis deriva de una
evaginación del suelo del diencéfalo, el infundíbulo (neuroectodermo).
El desarrollo de la bolsa de Rathke está controlado por BMP-4 (proteína ósea
morfogenética 4). El crecimiento de la bolsa es controlado por el FGF-8 (factor de
crecimiento fibroblástico 8), que activa los genes Lhx3 y Lhx4.
Ambos divertículos contactan y se pierde la conexión con el estomodeo. La
conexión con el infundíbulo persiste y forma el tallo neural. La porción inferior
prolifera y se diferencia en pars nervosa (pituicitos). Hay desarrollo de fibras nerviosas
hacia el tallo neural y la pars nervosa.
La porción de la bolsa de Rathke en contacto con la pars nervosa se transforma
en pars intermedia y crece poco. En el hombre solo en época fetal, no fisiológica. El
resto crece mucho y forma la pars distalis, cerca de la cual crecen dos lengüetas
alrededor del tallo neural y se fusionan para formar la pars tuberalis.
Pueden quedar restos de la luz de la bolsa en forma de pequeñas vesículas con
coloide junto a la neurohipófisis. La hipófisis faríngea está formada por restos de la
comunicación de la bolsa con estomodeo, puede derivar en tumores productores de
hormonas.
VASCULARIZACIÓN
Es clave en la regulación de la función hipofisiaria. La arteria carótida interna
origina las arterias hipofisarias:
x Superior: Irrigan la eminencia media, tallo hipofisario y pars distalis.
x Inferior: Irrigan la pars nervosa.
Ambas arterias conectan a través de la arteria trabecular. La mayor parte del
lóbulo anterior no tiene irrigación arterial directa.
Plexo capilar primario
Está formado por capilares fenestrados en la parte superior del tallo neuronal, a
partir de la arteria hipofisaria superior. Recoge la secreción de los núcleos
hipofisotropos hipotalámicos (hormonas liberadoras/inhibidoras). Drena por venas porta
hipofisarias que van hacia la pars tuberalis.
Hormonas hipofisótropas:
x GHR, GHRH: Hormona liberadora de hormona de crecimiento.
x GIH, somatostatina: Hormona inhibidora de hormona de crecimiento.
x PRH: Hormona liberadora de prolactina.
407
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x TRH: Hormona liberadora de tirotrofina.
x GnRH: Hormona liberadora de gonadotrofinas.
x CRH: Hormona liberadora de corticotrofina.
Plexo capilar secundario
Está formado por capilares fenestrados en la pars distalis, formado a partir de
venas porta hipofisiarias.
Sistema porta hipotálamo-hipofisario
Formado por los plexos capilares primario y secundario. Se logra la integración
funcional del hipotálamo con la adenohipófisis.
x Llevar hormonas liberadoras o inhibidoras a células de la adenohipófisis.
x Recoge las secreciones de la pars distalis, que pasarán a la circulación sistémica.
La mayor parte de la sangre de la adenohipófisis es drenada por venas
hipofisiarias al seno cavernoso, que se comunica con la circulación sistémica.
Venas porta breves se dirigen desde la pars distalis a la pars nervosa y de ahí al
hipotálamo. Se forma un sistema de retroalimentación de la pars distalis sobre el
hipotálamo.
Plexo capilar terciario
Se encuentra en la pars nervosa. Procede de la arteria hipofisiaria inferior y
recoge las secreciones de la neurohipófisis.
408
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
PARS DISTALIS
Ocupa el 75% y es la principal glándula endocrina, encargada de regular al resto
de glándulas endocrinas. Además, regula algunos tejidos no endocrinos.
El parénquima contiene células glandulares (productoras de GH, TDH, ACTH,
LTH, FSH y LH) y células folículoestrelladas. El estroma está constituido por una red
de fibras reticulares. Existe una red capilar fenestrada (“sinusoides”).
Células foliculoestrelladas
Son epiteliales y estrelladas, con largas prolongaciones. Están unidas por nexos
y tienen función de sostén, delimitando folículos junto a la pars nervosa. Exhiben MHC
y producen IL6.
Las células glandulares se dividen en cromófobas, acidófilas y basófilas (no
división funcional, sino histológicas).
409
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
En los cordones existe predominio de células acidófilas y de células cromófobas.
Células cromófobas
Representan un 50% y se acepta que pueden ser cualquier tipo de célula que han
descargado ya las hormonas, perdiendo sus características tintoriales. Son pequeñas.
Células acidófilas
Son un 40% y más abundantes en regiones laterales. Tienen morfología
redondeada, no muy grandes, con gránulos eosinófilos. Hay dos tipos:
x Somatotróficas: Producen hormona de crecimiento.
Son un 50% del total hipofisiario. Se sitúan sobre
todo en regiones laterales y tienen RER y Golgi
abundantes, además de gránulos de secreción
densos.
410
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Son estimuladas por GHRH y son inhibidas por somatostatina (GIH) y por
niveles altos de glucemia.
x Lactotróficas o mamotróficas: Producen prolactina.
Constituyen un 15-20% del total hipofisario. Su distribución es uniforme y su
forma es poliédrica y grande. En gestación y lactancia aumenta en número y
tamaño. También aumenta el RER, Golgi y los gránulos. Los gránulos densos
miden entre 600 y 900 nm. Tras la lactancia se eliminan por autofagia.
Las células lactotróficas responden a dopamina, que tiene un efecto inhibitorio.
La PRL (hormona liberadora de prolactina) estimula la secreción y se
retroinhibe y la TRH también tiene un efecto estimulante.
La GH se libera de forma pulsátil, máximo al inicio del sueño. Estimula los
hepatocitos para segregar IGF1 o somatomedina C y estimula la zona de reserva de
cartílago metafisiario para crecimiento en longitud de los huesos largos y estimula la
diferenciación de células satélites para el desarrollo de la musculatura esquelética.
Clínica:
x Adenoma: Tumor benigno por exceso de GH.
x Gigantismo: Tumor secretor en la infancia o pubertad que hace que los
cartílagos epifisiarios crezcan en exceso.
x Acromegalia: Tumor secretor de GH en adultos provoca hipertrofia de manos,
pies, mandíbula y tejidos blandos. Provoca deformidades macroscópicas (nariz,
orejas, mandíbula y cráneo). Huesos largos no aumentan de longitud.
x Enanismo hipofisiario: Déficit en GH en niños. Se trata administrando GH
humana sintetizada mediante tecnología genética.
La prolactina se secreta de forma pursátil, con cada succión. Estimula el inicio y
mantenimiento de la lactancia (mamogénesis, lactogénesis y galactopoyesis).
La hiperprolactinemia se produce por un tumor secretor de prolactina que
modifica el eje hipotalámiohipofisiario ocasionando una deficiencia de gonadotropinas.
411
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
En mujeres puede llevar a galactorrea y a infertilidad por falta de ovulación y
amenorrea. En hombres reduce la fertilidad y la líbido, y puede provocar galactorrea.
Células basófilas
Forman un 10%. Son redondeadas y más grandes, con gránulos ligeramente
basófilos, PAS+. Tipos:
x Células tirotróficas (5%): Producen tirotrofina, tirotropina o TSH (hormona
estimulante del tiroides).
Más abundantes en la región anterior. Son grandes, poliédricas, con un núcleo
redondeado y excéntrico. Es abundante en orgánulos, especialmente Golgi, con
gránulos muy pequeños y densos (100-150 nm).
Son estimuladas por la TRH e inhibidas por aumento de las hormonas tiroideas
T3 y T4.
La TSH o tirotropina regula la función tiroidea y el crecimiento, además de
estimular la liberación de prolactina.
412
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
El hipotiroidismo producido por un déficit en la secreción de la TSH ocasiona
una reducción del metabolismo celular, temperatura y metabolismo basal,
además de lentitud mental.
x Células gonadotróficas (10%): Producen FSH y LH.
Distribución uniforme. Cada célula sintetiza tanto FSH como LH. Se
empaquetan juntas en gránulos pequeños (200-250 nm).
Son estimuladas por GnRH, producido en el núcleo arciforme del hipotálamo.
La secreción de GnRH es pulsátil, cada 60-90 minutos.
La liberación de FSH es potenciada por activina, segregada por células
foliculares y de Sertoli; mientras que es inhibida, junto a la GnRH, por inhibina,
segregada por células foliculares, de Sertoli y de la pars distalis, y estradiol. Sus
funciones son:
o Estimula el desarrollo de folículos ováricos.
o Estimula a células foliculares para síntesis y secreción de activina,
inhibina y estradiol.
o Estimula la producción de estrógenos y de ABP en células de Sertoli.
Las funciones de la LH son:
o Estimula la maduración folicular, la ovulación y la formación del cuerpo
lúteo.
o Estimula la estereidogénesis (progesterona) en el folículo ovárico y en el
cuerpo lúteo.
o Estimula la síntesis de testosterona en células de Leydig.
Déficits en FSH y LH pueden llevar a reducción de la fertilidad y de las
funciones reproductoras. En mujeres provoca trastornos menstruales. En varones
causa hipogonadismo hipogonadotropo (testículos pequeños e infertilidad).
La ooforectomía y orquiectomía (castración) determina aumento significativo de
la síntesis de FSH y LH. Las células gonadotróficas se hipertrofian y muestran
aspecto vacuolado (células de la castración).
413
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Células corticotróficas (20%): Producen corticotrofina o ACTH.
Son más abundantes en la región media y tienen morfología poligonal o
estrellada. Su núcleo es excéntrico, el citoplasma claro, hay abundantes
mitocondrias, cisternas cortas de RER y gránulos de secreción pequeños (200-
250 nm) escasos, en la periferia.
Están estimuladas por CRH, sintetizada en núcleos paraventriculares
hipotalámicos, estrés y vasopresina. Inhibidas por cortisol.
Sintetizan precursor POMC (proopiomelanocortina), también se cree que en
hipotálamo hígado, corazón, riñones, tejidos digestivos y órganos reproductores.
POMC genera:
o N-terminal: Función desconocida.
o Hormona β-lipotrófica (β-LPH): Rinde en γ-LPH y β-endorfina.
β-LPH y γ-LPH son hormonas liposolubles. γ-LPH contiene la secuencia
de β-MSH, en el hombre no secretada.
β-endorfina contiene secuencias met-enk, aunque en el hombre no hay
evidencias de que se genere.
o ACTH: Síntesis circadiana, con picos matutinos. Estimula las zonas
fascicular y reticular de la corteza suprarrenal, síntesis de
glucocorticoides y hormonas esteroides sexuales. Además, aumenta la
pigmentación de la piel y estimula la lipólisis.
En vertebrados inferiores se degrada y rinde α-MSH y CLIP (péptido
parecido a corticotropina).
414
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
α-MSH y β-MSH actúan regulando la dispersión de gránulos de
melanina en melanóforos y cambios de coloración de la piel (peces,
anfibios y reptiles).
La enfermedad de Cushing es un trastorno debido a un adenoma
adenohipofisario secretor de ACTH que conduce a un aumento de
cortisol, obesidad, osteoporosis y atrofia muscular.
Déficit en ACTH reduce la secreción suprarrenal de andrógenos. En
mujeres ocasiona pérdida del vello axilar y púbico.
PARS INTERMEDIA (adenohipófisis)
La pars intermedia rodea una serie de pequeñas
cavidades quísticas que son los restos de la bolsa de Rathke.
En otras especies se encuentra mejor desarrollada, pero en
el hombre solo tiene una actividad similar en la etapa fetal y
en un corto periodo de tiempo posnatal.
Las células de la pars intermedia son basófilas y
cromófobas, y se organizan en cordones que invaden la pars
nervosa, lo cual se conoce como infiltración basófila. Las
células parenquimatosas pueden formar folículos, los cuales
muestran un epitelio estratificado en la parte anterior y
cúbico en posterior.
En poiquilotermos, como las ranas, las células basófilas producen alfa y beta
MSH, que estimula la producción de pigmento en los melanocitos y la dispersión del
pigmento en los melanóforos. En los seres humanos la MSH no es una hormona
415
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
funcional bien definida, sino que se trata de un subproducto del procesamiento
postraduccional de la beta-LPH. Dado que en la pars intermedia humana hay una
cantidad reducida de MSH, salvo en el periodo fetal (alfa MSH abundante), las células
basófilas se consideran corticotrofas, ya que tanto ACTH como MSH tienen secuencias
similares, puesto que derivan del POMC.
PARS TUBERALIS (adenohipósis)
La pars tuberalis es una extensión del lóbulo anterior a lo largo del tallo
pituitario, que constituye una capa celular delgada alrededor del mismo. Es una región
muy vascularizada, pues contiene las venas del sistema porta hipotálamo hipofisario.
Las células predominantes son las basófilas, y están distribuidas en cúmulos y folículos
y en cordones pequeños relacionados con los vasos sanguíneos. Aunque poseen
receptores para melatonina, su función endocrina no está determinada.
NEUROHIPÓFISIS
Está constituida el lóbulo posterior, que es una extensión del SNC, el tallo
neural, que es el eje del tallo hipofisario, y la eminencia media.
Pars nervosa
Es una extensión del SNC, por tanto, se trata de una extensión del HPT que
almacena y libera los productos almacenados en él. La pars nervosa está constituida por
axones procedentes del HPT y pituicitos, y en ella encontramos una importante red de
capilares fenestrados.
Los pituicitos son células gliales especializadas asociadas a los capilares
fenestrados. Estas células son de forma irregular y tienen muchas ramificaciones, por lo
que se parecen a los astrocitos y, como ellos, tienen filamentos intermedios específicos
de GFAP. Presentan en su citoplasma vesículas lipídicas llenas de pigmento. Sus
prolongaciones largas contactan tanto con los axones como con el endotelio y la lámina
basal capilar, con lo que al retraerse permiten la liberación de NT a la sangre. En
definitiva, el pituicito simple cumple una función similar a la de los astrocitos en el
resto del SNC.
416
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Fibras nerviosas
Las fibras nerviosas proceden de los somas de neuronas neurosecretoras situados
en los núcleos supraóptico y paraventricular hipotalámicos, y que acaban vertiendo su
secreción a los capilares en vez de liberar NT en una sinapsis.
x Tracto hipotálamohipofisario: Constituido por unos 100 000 axones amielínicos
que podemos observar a la altura del infundíbulo. Está a su vez constituido por:
1. Tracto tuberoinfundibular: Contiene axones procedentes de los núcleos
ventromedial, dorsomedial e infundibular, que van hasta la eminencia
media para liberar hormonas estimuladoras e inhibidoras al plexo capilar
primario.
417
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
A la eminencia media acaban llegando tanto estos axones como axones
procedentes de neuronas parvocelulares de los núcleos supraóptico y
paraventricular, que se encargan de la secreción de ADH y oxitocina. Por
tanto, finalmente la sangre va a contener hormonas estimuladoras e
inhibidoras de la secreción de otras hormonas adenohipofisarias, ADH y
oxitocina, y se dirigirá a la pars distalis.
2. Tracto supraópticohipofisario: Contiene axones también procedentes de
neuronas de los núcleos supraóptico y paraventricular, pero esta vez
magnocelulares, que van por el tallo neural hasta la pars nervosa. Las
procedentes del núcleo supraóptico secretan ADH, las del núcleo
paraventricular secretan oxitocina. En las terminaciones axónicas se
acumulan vesículas de secreción, dilatando los segmentos axónicos. Estas
dilataciones, denominadas cuerpos de Herring, sirven para almacenar
temporalmente las hormonas, se tiñen con sales Cr/Al y se observan al
microscopio óptico.
Transporte de las hormonas a lo largo de los axones de ambos tractos
De él se encarga la proteína neurofisina, que forma un complejo por medio de
enlaces no covalentes con la hormona a transportar. La ADH, la oxitocina y la
neurofisina derivan de un precursor común.
x Oxitocina -neurofisina I.
x ADH - neurofisina II.
Regulación hormonal
Oxitocina
Promotor de la contracción muscular lisa. Promueve la contracción de:
x Músculo liso uterino durante el orgasmo, la menstruación y el parto, como
consecuencia de la distensión de la vagina y el cérvix.
x Células mioepiteliales de los alveolos secretores y los conductos excretores de la
glándula mamaria, como consecuencia del reflejo de succión.
x Conducto deferente en hombres, se especula con ello, pero no está claro.
ADH
Su liberación se haya estimulada por una disminución del volumen sanguíneo o
aumento de la presión osmótica plasmática, y en menor medida por dolor,
traumatismos, tensión emocional o nicotina. Al contrario, una retención de agua, que
conlleva un aumento del volumen sanguíneo, inhibe la secreción de ADH.
El efecto fisiológico primario de la ADH es aumentar la permeabilidad de las
porciones distales en la nefrona, lo cual contribuye a concentrar la orina. Asimismo, las
grandes dosis no fisiológicas aumentan la tensión arterial, porque promueven la
contracción del músculo liso en las arterias pequeñas y las arteriolas.
418
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
EPÍFISIS O GLÁNDULA PINEAL
El nombre de pineal se toma de la palabra “piña” (sí, como en la que vive Bob
Esponja) puesto que es un órgano con forma de pequeña piña.
Esta estructura se forma a partir de una evaginación en el techo del tercer
ventrículo, concretamente en la línea media. Tiene un tamaño pequeño en torno a los
5mm de diámetro y un peso alrededor de los 150 g.
DESARROLLO EMBRIONARIO
En el desarrollo embrionario se constata la aparición de una pequeña
evaginación en forma de saco, en el techo de lo que será posteriormente el diencéfalo,
concretamente en la línea media del III ventrículo.
Esta pequeña evaginación sacular avanza, crece y se pliega, creando finalmente
una masa sólida de células que está envuelta por un tejido conjuntivo derivado de las
meninges.
ESTRUCTURA HISTOLÓGICA
Histológicamente, esta glándula pineal está envuelta por una cápsula fina que
deriva de la piamadre; de la cual van a partir finos tabiques que permiten lobular esta
glándula. Se trata de una estructura en la que encontramos un estroma con abundantes
vasos y también fibras nerviosas.
419
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Tenemos también un parénquima en el que se distinguen los pinealocitos y unas
células de carácter glial que se denominan células intersticiales. Además, como
recordaréis del apartado del Sistema Nervioso, la glándula pineal es una de las
excepciones del SN en cuanto a barrera hematoencefálica se refiere, puesto que no hay
barrera HE.
Pinealocitos
Son células grandes, muy abundantes y de aspecto pálido claro, que contienen en
su interior un núcleo redondeado generalmente centrado, en el que destaca un nucléolo
muy prominente.
Bajo la membrana plasmática es fácil encontrar abundantes cisternas de REL, en
las cuales se almacena Ca2+. También se observa un desarrollo considerable del RER y
del AG. Es también posible encontrar restos de un cilio presente durante la etapa
embrionaria.
Además, en estas células son muy abundantes las mitocondrias. Es frecuente que
también haya un almacén de lípidos, siendo lo más característico unos gránulos de
secreción pequeños en torno a los 200 nm y que son muy electrodensos.
También hay Sinapsis en Barra o en Cinta. Son característicos de los
pinealocitos, pero también ya se comentaron en retina y oído interno.
Los pinealocitos son células que presentan algunas prolongaciones, no muchas,
generalmente 1 o 2. En algunas ocasiones son cortas, en otras son más largas. Terminan
expandiéndose, generalmente adosadas a capilares. En estas prolongaciones lo más
destacado son los haces de microtúbulos y las mitocondrias.
Los pinealocitos suelen aparecer agrupados, a veces formando cordones. Entre
ellos es frecuente la aparición de uniones de tipo GAP (uniones en hendidura). Estos
pinealocitos se apoyan en la lámina basal y son las células que se encargan de sintetizar
y secretar melatonina.
420
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Células Intersticiales
Las células intersticiales son células de tipo glial. Aparecen en bajo número
(5%) y suelen estar o bien rodeando los pinealocitos o bien situados entre estos.
Estas células son mucho más pequeñas, presentan núcleos más pequeños y
oscuros y son positivas a la GFPA. Estas células gliales tienen como función dar soporte
a los pinealocitos.
Cuerpos Arenáceos
El conjunto de los cuerpos arenáceos se conoce como arenilla cerebral. Son
masas muy basófilas que destacan en el parénquima.
Estos cuerpos arenáceos son concreciones de fosfato cálcico. Estas concreciones
comienzan a encontrarse inicialmente en el interior del pinealocito, en su citoplasma.
Posteriormente, van a ser expulsadas. Una vez fuera, en la membrana plasmática, van a
precipitar formando cristales.
Hay que decir que la formación de estos cristales, esta calcificación de la
glándula pineal, no afecta a su función. Por lo tanto, hay que contemplarlo como un
proceso natural y fisiológico y no como un proceso degenerativo.
La aparición de la arenilla cerebral comienza a partir de la primera década.
Posteriormente, estos cuerpos arenáceos van a ir aumentando.
Esta calcificación de la glándula pineal tiene un valor clínico, puesto que hace
dar positivo en las radiografías. Por lo tanto, dado que la glándula pineal se sitúa en la
línea media, nos sirve como punto de referencia. Así pues, un desplazamiento de la
pineal en referencia a esa línea media, nos indica una posible presencia de una masa
tumoral o un proceso como una hemorragia cerebral.
INERVACIÓN
La inervación de la glándula pineal corre a cargo fundamentalmente de fibras
simpáticas. Estas fibras simpáticas postganglionares proceden del ganglio cervical
superior y alcanzan la pineal a través del nervio coronario.
En el interior de la pineal producen descargas sinápticas a través de dos
neurotransmisores: la noradrenalina y el neuropéptido Y.
Esta inervación simpática es la que lleva a estimular a los pinealocitos y, por lo
tanto, es la responsable de la síntesis y secreción de melatonina. Esto se pone de
manifiesto en experimentación animal. En animales en los que se produce la destrucción
bilateral del ganglio cervical superior cesa de inmediato la secreción de melatonina y no
pueden recuperar esta función.
Más recientemente se han descrito otras fibras nerviosas que inervan la pineal:
por una parte, fibras parasimpáticas y fibras que alcanzan la pineal directamente desde
el SNC. Estas últimas se tratarían de fibras procedentes de la cisura posterior y que
legarían a la pineal atravesando la cintilla óptica.
421
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
FUNCIÓN
La glándula pineal es una glándula muy antigua que aparece evolutivamente ya
en vertebrados inferiores. En estos animales se trata de un órgano sensible a la luz, por
lo cual en muchos textos se ve descrita como ojo parietal o tercer ojo.
Se trata por lo tanto de una estructura fotosensible y esto es posible porque en
estos animales, la glándula pineal se encuentra inmediatamente por debajo de la piel; es
decir, fuera del cráneo.
Así que, esta glándula actúa como fotorreceptor, capta información lumínica y la
envía al cerebro a través del nervio pineal.
Sin embargo, en los mamíferos, esta función de fotorrecepción se ha perdido,
puesto que la glándula pineal está dentro de la caja craneal.
Igualmente, la luz va a seguir teniendo un efecto indirecto sobre la epífisis. Esta
epífisis en los mamíferos tiene un carácter de glándula endocrina. Esta glándula
endocrina tiene como función básica la síntesis y secreción de melatonina a partir de
triptófano, concretamente a partir de la serotonina.
Hay que decir que se trata de una hormona que no se puede almacenar, por lo
tanto, según se sintetiza, se debe excretar. Su síntesis máxima se va a producir durante
el periodo nocturno; es decir, en ausencia de luz. Cuando se produce la exposición a la
luz, se produce la inhibición instantánea de la síntesis de melatonina.
422
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Como se observa en el esquema, la información es captada a nivel de la retina,
pasa hasta el núcleo supraquiasmático y, a partir de aquí, viajando por el tracto
hipotalámico espinal, llega hasta la médula espinal. Desde aquí se va a producir el
trasbase de información hasta el ganglio cervical superior. Desde el ganglio cervical
superior se va produce la llegada de información por las fibras postganglionares
simpáticas hasta la glándula pineal.
Melatonina
La melatonina es una hormona que está relacionada directamente con la
inducción del sueño. Cuando se comienza a sintetizar melatonina es cuando empezamos
a notar esa sensación de somnolencia.
Por lo tanto, podemos decir que la melatonina tiene un papel clave en la
regulación de los ritmos circadianos; es decir, en la regulación del ritmo que estipula la
duración de la noche y el día, el ritmo de 24 horas aproximadamente.
La información lumínica es captada en la retina por las células ganglionares, las
cuales van a enviar esa información lumínica al cerebro. Además, estas células
sintetizan una molécula, la melanopsina, que va a ser transferida hasta el núcleo
supraquiasmático. La información también llega al ganglio cervical superior y, por lo
tanto, esta información alcanza la pineal.
El núcleo supraquiasmático es el órgano que va a desencadenar los ritmos
circadianos. Por ello, podemos denominarlo como el marcapasos endógeno de los
ritmos circadianos. Estos ritmos circadianos creados por el núcleo supraquiasmático van
a ser regulados por la melatonina que ha sido sintetizada por los pinealocitos.
La melatonina se ha puesto de manifiesto como una molécula que tiene que ver
con la adaptación que sufre el organismo cuando viajamos varios husos horarios de una
manera rápida (es lo que se conoce como adaptación al jet-lag).
Es una molécula que tiene que permitir una adaptación rápida cuando se produce
un cambio súbito en los ciclos día-noche.
También se ha puesto de manifiesto su papel en personas que tienen cambios
crónicos en la duración del ciclo día-noche. Por ejemplo, los trabajadores que tienen
turnos laborales que van rotando, trabajando unos días por la mañana, otros por la tarde
y otros por la noche. En estas personas se produce una afectación de los ciclos
circadianos y en ellos la melatonina juega un papel fundamental para conseguir una
buena adaptación.
En diferentes trabajos, se ha visto el papel que juega esta hormona en los
cambios que sufren las respuestas emocionales según la estación del año. Es lo que se
conoce como trastornos afectivos estacionales.
La melatonina es una hormona que va a controlar los ritmos corporales diarios.
Esto es así porque actúa sobre el eje hipotálamo-hipofisario. Esta hormona, es la que
controla que las distintas hormonas que se segregan en el organismo, fundamentalmente
las hipotalámicas, pero no solo ellas (también los neurotransmisores) lo hagan conforme
423
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
a un patrón característico que es un patrón circadiano propio de cada hormona o de cada
neurotransmisor.
Hay que señalar que este control de los ritmos circadianos se pone de manifiesto
no sólo en la etapa postnatal, sino también se detecta ya un ritmo circadiano en la etapa
fetal. Esto es posible porque la melatonina materna atraviesa la membrana placentaria y
desencadenan ya la regulación de los ritmos a nivel del feto.
En otras especies, no en el caso del hombre, la melatonina también tiene que ver
con la regulación de la actividad sexual estacionaria. Hay especies que no son fértiles a
lo largo del año, sino que tienen periodos fértiles.
La melatonina tendría que ver con la inhibición de las gonadotropinas, por lo
tanto, con la inhibición de las gónadas y los órganos anejos. Esto causaría la involución
temporal de estos órganos, lo que causaría que hubiera periodos de no fertilidad.
Una de las últimas funciones descubiertas en la melatonina tiene que ver con su
función protectora, presente en humanos. La melatonina protege frente a radicales libres
y a esto se le conoce como función antioxidante.
CLÍNICA
Los pinealomas son tumores muy destructivos que destruyen completamente la
pineal y suelen aparecer en personas, en niños, que tienen una pubertad muy precoz
(varones con espermatogénesis antes de los nueve años o niñas que tienen una secreción
estrogénica y una primera menstruación antes de los ocho años).
En estas personas es muy frecuente la aparición de un síndrome neurológico
conocido como Síndrome de Parinaud. Este síndrome es un trastorno neurológico muy
característico que muchas veces es lo que ayuda al diagnóstico del pinealoma.
424
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
TIROIDES
La glándula tiroides está situada en la región anterior de la base del cuello
delante de la tráquea y por debajo de la laringe. Se trata de una glándula endocrina que
consiste en dos lóbulos laterales grandes unidos por un itsmo, que es una delgada banda
de tejido tiroideso. Con frecuencia, desde el itsmo se extiende hacia arriba un lóbulo
piramidal.
ORIGEN EMBRIOLÓGICO
La glándula tiroides comienza a desarrollarse durante la 4ª semana de gestación
a partir de un primordio originado como un engrosamiento endodérmico del piso de la
faringe primitiva. El primordio crece caudalmente y forma una invaginación canalicular
conocida como conducto tirogloso, que desciende a través del tejido del cuello hasta su
destino final frente a la tráquea, donde se divide en dos lóbulos. Durante esta migración
caudal el conducto tirogloso se atrofia, se convierte en un tubo ciego, y deja restos en el
40% de las personas (también consideramos el lóbulo piramidal como un resto del
conducto tirogloso), que pueden ser origen de quistes.
Alrededor de la 9ª semana de gestación, las células endodérmicas se diferencian
en láminas de células foliculares que se organizan en folículos. Para la 14ª semana los
folículos bien desarrollados con su revestimiento de epitelio folicular contienen coloide
en su luz. A partir de este momento, la glándula pasa a ser funcional.
Durante la 7ª semana cúmulos de células epiteliales que tapizan la invaginación
de la cuarta bolsa faríngea, región a veces denominada quinta bolsa faríngea, conocidos
como cuerpos ultimobranquiales, inician su migración a la glándula tiroides en
desarrollo, y se incorporan a los lóbulos laterales. Tras fusionarse con la glándula
tiroides, estas células se dispersan entre los folículos y dan lugar a las células
parafoliculares, que quedan incorporadas al epitelio folicular.
FUNCIONES DE LA GLÁNDULA TIROIDES
Se trata de una glándula endocrina capaz de sintetizar y segregar:
x Tiroxina (T4).
x Triyodotironina (T3).
x Calcitonina.
Responde a la TSH de la pars distalis de la hipófisis a partir de la semana 22 del
desarrollo.
425
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
ESTRUCTURA HISTOLÓGICA
La glándula está rodeada por una fina
cápsula de tejido conjuntivo que emite tabiques
hacia el interior del parénquima para delimitar
parcialmente lobulillos irregulares. Esta cápsula
tiene dos capas: una externa laxa y una interna
densa que contacta con el parénquima y desde
donde parten los mencionados tabiques. En
conjunto, ambas capas contribuyen a aislar la
glándula, lo cual facilita su extirpación
quirúrgica.
Las unidades funcionales y estructurales del parénquima de la glándula son los
folículos tiroideos, alrededor de los cuales encontramos tejido interfolicular con fibras
reticulares y capilares fenestrados.
FOLÍCULOS
Un folículo tiroideo es un compartimento quístico, más o menos esferoidal, que
tiene una pared formada por un epitelio cúbico simple o cilíndrico, el epitelio folicular
(que cambia de altura según su actividad), y cuyo tamaño varía entre 50 µm y 1 mm.
Los folículos contienen un material gelatinoso llamado coloide, que contacta con la
superficie apical de las células. La superficie basal está apoyada en una lámina basal
típica.
Células foliculares
Las células foliculares o células principales exhiben un citoplasma basal
basófilo, y un núcleo esferoidal con un nucléolo prominente. El RER y el aparato de
Golgi son de ubicación supranuclear. Hallamos en ellas tres tipos de gránulos o
vesículas:
x Gránulos subapicales, de 200 nm: Contienen tiroglobulina, sintetizada en el
RER y en el AG, que se expulsa por exocitosis.
x Gotas de coloide, de hasta 4 µm: Se trata de coloide que se ha introducido en la
célula por endocitosis.
x Vesículas muy densas similares a las que se hallan asociadas al AG: Se trata de
lisosomas fusionados con gotas de coloide.
Entre las células foliculares hallamos una gran cantidad de complejos de unión,
y su superficie apical, en contacto con el coloide, presenta microvellosidades cortas (no
se observa chapa estriada) y pseudópodos.
Los pseudópodos se extienden desde el dominio apical de las células foliculares
tiroideas y, tras rodear una porción de coloide (tiroglobulina), organizan un fagosoma
426
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
intracelular. Los lisosomas se fusionan con el fagosoma e inician la degradación
proteolítica de la tiroglobulina mientras se desplazan hacia el dominio basal de la célula
folicular.
Coloide
Se trata de una secreción de naturaleza exocrina, que expulsan las células
foliculares. Se tiñe con colorantes ácidos y básicos, y con PAS, pero suele artefactarse
en las preparaciones.
El componente principal del coloide es la tiroglobulina, una glucoproteína
yodada de gran tamaño, aunque también existen en el coloide otras enzimas y
glucoproteínas. La tiroglobulina no es una hormona, sino la forma inactiva de
almacenamiento de las hormonas tiroideas.
427
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Fase exocrina de las células foliculares
Las células foliculares captan de la sangre ion yoduro por su membrana basal.
Este es transportado al interior de la célula activamente, por medio de una ATPasa (que
puede ser inhibida con perclorato), lo cual permite concentrar en el LIC grandes
cantidades de yoduro (hasta 200 veces más de lo normal), que puede así difundir y salir
por la membrana plasmática apical hacia el coloide.
Paralelamente, en el RER y el AG tiene lugar la síntesis de tiroglobulina y de
peroxidasa tiroidea, que se acumulan en vesículas que posteriormente serán exocitadas
por la membrana apical. En este espacio, la peroxidasa entrará en contacto con el
yoduro, transformándolo en yodo, de manera que en la periferia el yodo se unirá a la
tiroglobulina y se producirá la yodación de la misma. El propiltiouracilo y el metil
mercaptoimidazol (MMI) inhiben a la peroxidasa y con ello el proceso de yodación, con
lo que se usan para el tratamiento de las glándulas hiperactivas.
Fase endocrina de las células foliculares
La TSH hipofisaria estimula la endocitosis de tiroyodoglobulina por parte de las
células foliculares, que se realiza por emisión de pseudópodos hacia el coloide y
captación de gotas del mismo que viajan por el citoesqueleto hasta los lisosomas y se
fusionan con ellos.
La degradación lisosomal de la tiroyodoglobulina genera T3 y T4, así como
metabolitos intermedios (MIT, DIT). La cantidad de T4 generada es 20 veces superior a
la de T3, pero en cualquier caso ambas hormonas son liberadas a los capilares
fenestrados del espacio interfolicular a través de la membrana basal, y se transportan en
sangre unidas a proteínas séricas. El propiltiouracilo es también capaz de bloquear la
conversión de T4 en T3 en los tejidos periféricos.
Células parafoliculares o células C
Se trata de células procedentes de la cresta neural, que forman parte del sistema
neuroendocrino difuso. No están expuestas a la luz folicular, luego no contactan con el
coloide, y secretan calcitonina, hormona que regula el metabolismo del calcio.
Estas células solo constituyen un 0,1% de las células de la glándula. Las
podemos hallar aisladas o en grupos, en cuyo caso se unen entre sí mediante
desmosomas, aunque son difíciles de distinguir (a no ser que sea por Ag o Ac
específicos). Son más grandes y claras que las foliculares, presentan núcleos mayores y
más claros. De igual modo, tienen un RER y un AG prominentes. Presentan vesículas
electrodensas pequeñas, de unos 60 a 150 nm.
La calcitonina, sintetizada por las células C es un antagonista fisiológico de la
PTH. Su gen se expresa también en HPT e hipófisis, y aunque se sabe que CGRP es un
neurotransmisor y tiene efectos vasodilatadores, el déficit parcial o la carencia absoluta
de calcitonina tras una tiroidectomía no tiene efectos clínicos.
428
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
FUNCIONES DE LAS HORMONAS
T3 y T4
Aunque las células foliculares producen ambas hormonas, T4 es exclusivamente
sintetizada a este nivel, mientras que T3 se produce por reconversión de T4 en hígado,
pulmones y corazón mayoritariamente.
Ambas hormonas actúan uniéndose al ADN (TRE), y aumentan el consumo de
oxígeno y por ende el metabolismo basal de las células. También aumentan la
producción de calor, la actividad cardiaca y las secreciones endocrinas.
Una importante acción de estas hormonas es el estímulo de la actividad
osteoblástica y, con ello, del crecimiento longitudinal de los huesos, con independencia
del consumo de oxígeno: de ahí que un déficit de hormonas tiroideas en la infancia
conlleve un retraso en el crecimiento.
Además, tanto T3 como T4 atraviesan la membrana placentaria, por lo que son
esenciales en el desarrollo inicial del SNC (aunque a partir de la 14ª semana la glándula
fetal ya es activa y capaz de producir hormonas).
Calcitonina
Su secreción por parte de las células C es independiente de las hormonas
hipofisarias. Antagoniza los efectos de la PTH, ya que disminuye la calcemia en sangre
al suprimir la acción resortiva de los osteoclastos y promover el depósito de calcio en
los huesos acrecentando el ritmo de calcificación del osteoide. Se regula por
retroalimentación: una calcemia elevada estimula la secreción, mientras que una
calcemia disminuida la inhibe.
CLÍNICA
Déficit de T4 y T3 durante el desarrollo fetal
Lleva a lesiones irreversibles del SNC y retraso mental por disminución del
número de neuronas, defectos de mielinización. Si hay déficit de las hormonas tiroideas
maternas antes del desarrollo del tiroides fetal, el retraso mental es más grave.
Hipotiroidismo congénito o cretinismo
Trastorno que se produce por la ausencia congénita de tiroides. Existe una falta
de T4 y T3, lo cual conlleva a su vez la falta de estimulación de las células somatotropas.
Por ello además de las lesiones neurológicas irreversibles se produce enanismo.
Bocio
Hipertrofia tiroidea provocada por hipo o hipertiroidismo. Puede ser:
429
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
x Endémico o por deficiencia de yodo (por falta de I en la dieta).
x Tiroiditis de Hashimoto (enfermedad autoinmune).
x Tóxico o enfermedad de Graves (hipertiroidismo).
Hipotiroidismo
Conlleva una reducción del metabolismo basal, hipotermia e intolerancia al frío.
Se produce asimismo una reducción de la sudoración, y vasoconstricción cutánea. Si no
se trata aparece mixedema, con trastornos cutáneos, enlentecimiento de la actividad
cerebral y somnolencia.
Un caso especial de hipotiroidismo debido a una reacción autoinmune frente a
la tiroglobulina y la peroxidasa tiroidea (Ac microsomales) es la enfermedad de
Hashimoto, que cursa con reducción del número de folículos y su funcionalidad.
Hipertiroidismo/ enfermedad de Graves
Enfermedad autoinmunitaria en la que el tiroides es hiperfuncional. Aumenta el
número y tamaño de los folículos, pero en ellos hay poco coloide. Se produce una
secreción muy elevada de H tiroideas, llegando a ser tóxicas, pese a que los niveles de
TSH son normales.
El mecanismo etiopatológico implica a los linfocitos T, los cuales segregan IL1,
TNF e interferón γ, que estimulan las células foliculares para producir más citocinas. A
la par, las células plasmáticas sintetizan Ac contra los receptores de TSH presentes en la
membrana basal de las células foliculares. Al imitar los Ac los efectos de la TSH, se
producen grandes cantidades de hormonas tiroideas de forma incontrolada.
Encontramos infiltrados linfocitarios en los músculos extraoculares y el tejido
orbitarioÆ diferenciación a adipocitosÆ edemaÆ exoftalmos.
Además, T3 estimula la bomba fosfolambano, implicada en la liberación y
captación de calcio en el retículo sarcoplásmico de los miocitos cardiacosÆ taquicardia
y temblores.
Adenoma tiroideo (neoplasia benigna): Nódulos hiperactivos (puntos “tibios o calientes”).
Carcinomas tiroideos: Son muy malignos, aunque en ellos la función tiroidea es normal.
Carcinoma medular de tiroides: Se asocia a las células C y a una producción excesiva de
calcitonina. Los niveles de calcio son normales y no hay lesiones óseas.
Enfermedad ósea de Paget
Conlleva un incremento de la actividad de los osteoclastosÆ amplia resorción
óseaÆ neoformación de tejido óseo mal organizado
Î Administración de calcitonina: en tratamiento de dolores relacionado con
metástasis esqueléticas (efectos a través del SNC).
430
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
PARATIROIDES
Las glándulas paratiroides son glándulas pequeñas en asociación con la parte
posterior del tiroides. Son ovoides, tienen pocos milímetros de diámetro y están
distribuidas en dos pares (4) que forman las paratiroides superiores e inferiores. Suelen
estar ubicadas en el conjuntivo de la parte posterior de los lóbulos laterales de la
glándula tiroides. Aun así, la cantidad y ubicación pueden variar: en el 2-10% hay
glándulas adicionales o accesorias en el cuello o mediastino. En total, la glándula pesa
unos 130 g, y tiene un color pardo amarillento.
Desde el punto de vista estructural, cada glándula paratiroides está rodeada por
una cápsula de tejido conjuntivo delgada que emite tabiques incompletos que dividen la
glándula en lobulillos mal definidos. El parénquima presenta adipocitos infiltrados a
partir del periodo puberal, que aumentan con el paso de los años, constituyendo en el
adulto la mitad de la masa glandular. El estroma es reticular y presenta sinusoides.
431
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
PARÉNQUIMA
Se organiza en cordones anastomosados con dos tipos celulares (puede que haya
folículos pero no está claro).
Células principales
Características
Son las más abundantes. Se trata de células poliédricas pequeñas, que poseen un
núcleo de ubicación central y claro. El citoplasma eosinófilo pálido contiene
acumulaciones de glucógeno extensas y vesículas densas lipídicas que se cree que son la
forma de almacenamiento de la PTH. Este tipo celular presenta asimismo un gran
desarrollo de RER y AG.
Funciones
Tienen a su cargo la síntesis, almacén y secreción de grandes cantidades de
PTH: la sintetizan a partir de un precursor que es procesado por proteólisis en el AG.
Estas células también presentan en su membrana plasmática un receptor sensor
de calcio (CaSR), con lo que se relacionan con el mantenimiento de la calcemia en los
tejidos y líquidos corporales. El ion Ca, al unirse a CaSR, activa una cascada de señales
que inhibe la secreción de PTH, de manera que si los niveles de ion Ca disminuyen se
activa la liberación de PTH. Esto se debe a que la PTH actúa regulando los niveles de
ion Ca y fosfato: presenta una acción lenta y sostenida que permite elevar los niveles de
Ca por estimulación de la reabsorción del hueso mineralizado mediante la activación de
la osteolisis osteoclástica, con lo que los efectos son opuestos a los de la calcitonina, la
cual actúa de manera rápida y efímera.
El efecto de la PTH no es directo sobre los osteoclastos: actúa sobre los
osteoblastos. Cuando estamos en situación de hipocalcemia, se segrega PTH, la cual
actúa sobre receptores presentes en los osteoblastos, provocando la síntesis de:
1. M-CSF: Activa el den RANK e induce la diferenciación de monocitos a
osteoclastos inmaduros
2. RANK-L: La unión de RANK a RANK-L provoca la diferenciación del
precursor del osteoclasto a un osteoclasto inmaduro. Sin embargo, este no
madura definitivamente, puesto que la acción de la osteoprotegerina lo impide.
En adición, PTH estimula la reabsorción renal de Ca y su absorción intestinal
(ya sea por vía directa o a través de vitamina D activa, cuya producción renal a partir de
25-dihidroxicolicalciferol también estimula) para elevar la calcemia, y la excreción
renal de fosfato.
La regulación de la PTH se realiza por retroinhibición: la hipercalcemia
retroinhibe la PTH.
432
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Células oxífilas
Aparecen solas o en cúmulos dispersos en los que las células están unidas entre
sí y a las principales por desmosomas. Pese a que comienzan a observarse a los 6-7 años
de edad y su número va en aumento, constituyen la porción menor de las células
paratiroideas, y son mucho más redondeadas y grandes que las células principales
Presentan un citoplasma muy eosinófilo con un núcleo pequeño muy basófilo. Sus
orgánulos son escasos, y no presentan vesículas de secreción, pero sí poseen un elevado
número de mitocondrias.
Su función es desconocida, se especula con que sean células de transición o
células principales en degeneración, pero esto Mari Pili no se lo cree porque una célula
que la vaya a palmar no tiene tantas mitocondrias ni de coña.
433
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
CLÍNICA
Hipoparatiroidismo
Puede ser:
x Por extirpación accidental del paratiroides en una cirugía tiroidea. A las 24-48h
se produce hipocalcemia y excitabilidad nerviosa con parestesias y crisis de
tetania y epilepsia. Causa asfixia y muerte si no se suministra PTH en un plazo
de tiempo breve.
x Idiopático: Incapacidad paratiroidea para responder a los niveles de calcio.
Existe una mutación de CaSR que condiciona que el paratiroides funcione como
si hubiera hipercalcemia y por tanto no segrega PTH.
Hiperparatiroidismo
Puede deberse a:
x Hipercalcemia benigna familiar: Una mutación en CaSR impide responder a los
niveles de Ca en sangre, de modo que aumenta la secreción de PTH. Si el
individuo es homocigoto para el alelo mutado se le ha de extirpar la glándula.
El hiperparatiroidismo produce un tumor benigno funcional o adenoma. Un
exceso de PTH lleva a fosfaturia, hipercalciuria, hipercalcemia por desmineralización
ósea y formación de quistes y cálculos renales.
434
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
GLÁNDULAS SUPRARRENALES
Órgano par situado sobre el polo superior de cada riñón, en el tejido adiposo
perirrenal. Su peso es de aproximadamente 10g. Está formada por una cápsula gruesa de
la que salen tabiques, una corteza externa y una médula interna (ambas con origen y
función diferentes).
x Corteza (80-90%): Tiene un color pardo amarillento, con origen mesodérmico.
Produce hormonas esteroides (derivadas del colesterol o corticoesteroides).
Esencial para la vida.
x Médula (10-20%): Tiene un color pardo rojizo y su origen es neuroectodérmico
(crestas neurales). Produce catecolaminas. No es esencial para la vida.
ORIGEN EMBRIOLÓGICO
En la sexta semana el epitelio del mesotelio
invade el mesonefros y forman la corteza fetal o
provisional. En la octava semana hay una segunda
invasión que forman la corteza permanente o definitiva.
El 80% es corteza fetal, acidófila, durante este periodo.
La corteza definitiva recuerda a la zona glomerular del
adulto, más basófila.
La corteza fetal se organiza en cordones con
células con orgánulos para la síntesis de lípidos. La
corteza definitiva tiene células más pequeñas y está
regulado por ACTH fetal y la placenta.
La unidad fetoplacentaria está formada por las
glándulas suprarrenales y la placenta, que interaccionan
para intercambiar moléculas y así poder sintetizar
glucocorticoides, aldosterona, andrógenos y estrógenos.
Durante la etapa fetal no se distingue médula. En
la séptima semana las células neuroectodérmicas emigran
435
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
más allá de los ganglios celiacos y alcanzan la glándula suprarrenal. Se van agrupando
en posiciones centrales hasta el nacimiento.
La masa máxima relativa es al final del cuarto mes. El tamaño es casi similar al
del riñón. Al final de la gestación se producen 100-200 mg de esteroides por día (el
doble que en el adulto).
La corteza fetal involuciona tras el nacimiento, se pierde el 50% del peso. La
corteza definitiva se desarrolla y forma 3 zonas. La diferenciación acaba a los 4 años.
Las células cromafines se aglomeran tras la desaparición de la corteza fetal y forman la
médula.
Durante el periodo fetal sus funciones son
x Síntesis de DHEA (dehidroepiandrosterona), pasa al SCT que lo transforma en
estrógenos y se eliminan por orina de la madre. En caso de muerte fetal se
produce una caída brusca de estrógenos en orina, hecho que tiene valor
diagnóstico.
x Síntesis de glucocorticoides esenciales en el feto para la producción de
surfactante por neumocitos II (octava semana), el desarrollo del eje hipotálamo
hipofisario y la inducción de la involución tímica.
436
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
CORTEZA
Tiene tres zonas concéntricas:
x Glomarular: mineralocorticoides
x Fascicular glucocorticoides y
x Reticular hormonas sexuales
No hay gránulos de secreción. No
hay límites definidos entre las zonas.
437
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Zona glomerular
Bajo la cápsula, ocupando un 10-15% de la corteza. Sus células están ordenadas
en grupos redondeados o arcos con capilares fenestrados (“sinusoides”) alrededor.
Es una zona hipercromática con núcleos apiñados. El núcleo es redondo y muy
basófilo y el citoplasma tiene grumos basófilos de RER. El REL es abundante, como el
Golgi, y tiene mitocondrias con crestas transversales. Hay escasas gotas pequeñas de
lípidos.
Carecen de 17α-hidroxilasa. No pueden sintetizar cortisol ni esteroides, pero sí
mineralcorticoides como la aldosterona.
Tiene capacidad regenerativa, por lo que puede reemplazar a las zonas fascicular
y reticular tras la extirpación quirúrgica (se desdiferencias para luego diferenciarse
siguiendo una vía distinta).
Zona fascicular
Forma un 75% de la corteza, con células ordenadas en cordones rectos y largos.
Hay capilares fenestrados (“sinusoides”) entre los cordones.
Sus células son grandes y cúbicas, con un núcleo grande, central y claro,
pudiendo ser, binucleado. El citoplasma tiene aspecto espumoso (espongiocitos) y es
acidófilo. Está lleno de gotas lipídicas grandes. El REL es abundante y las mitocondrias
tienen crestas tubulares.
No pueden sintetizar aldosterona. Contienen 17α-hidroxilasa y sintetizan
glucocorticoides (cortisol). Contienen 17,20 hidroxilasa y sintetizan esteroides sexuales.
438
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Zona reticular
Ocupa un 10-15% de la corteza con células formando una red anastomosada de
cordones cortos. Hay capilares fenestrados (“sinusoides”) entre los cordones.
Sus células son cilíndricas y acidófilas, con abundantes lisosomas. Hay escasas
gotas de lípidos, pero sí gránulos de lipofucsina. Menor desarrollo de REL y
mitocondrias con crestas tubulares.
Su secreción de glucocorticoides es escasa, principalmente es productora de
esteroides sexuales: DHEA y andostendriona.
Regulación
x Zona glomerular: Angiotensina II.
Las células glomarulares no responden al cortisol porque se convierten en
cortisona (hepatocitos).
La disminución de la tensión arterial o en niveles de sodio, aumenta la secreción
de renina, formación de angiotensina I y II y estimula la síntesis de aldosterona.
Alteraciones:
o Aldosteronismo primario: Producción excesiva de aldosterona que
suprime el sistema renina-angiotensina y disminuye la actividad de la
renina plasmática. Debido a: hiperplasia nodular (hipertensión más
ligera), tumor en la zona glomerular (síndrome de Conn) y gen
quimérico producido por la fusión de dos encimas (11 beta hidroxilasa y
aldosterona sintasa).
o Aldosterino secundario: Secreción excesiva de aldosterona debido a
aumento en la secreción de renina por un descenso de la perfusión renal,
embarazo o hipertensión arterial.
x Zona fascicular y reticular: ACTH.
o Enfermedad de Cushing: Aumento en la secreción de cortisol y
andrógenos suprarrenales debido a un tumor en la adenohipófisis que
produce un exceso de ACTH. Cambio hialino de Crooke, el citoplasma
de las células corticotrófica se hace más homogéneo y pálido.
Puede haber hipercortisolismo dependiente de ACTH por secreción...
o Síndrome de Cushing, hipercortisolismo no dependiente de ACTH:
Hiperproducción de cortisol y andrógenos suprarrenales por tumor en la
corteza suprarrenal o administración de glucocorticoides exógenos (s.
Cushing yatrogéno).
Funciones
Aldosterona
La aldosterona, sobre todo en el túbulo contorneado distal y en el túbulo
colector, aumenta la reabsorción de sodio y la excreción de agua. Va a aumentar la
secreción renal de potasio e hidrogeniones y la retención de agua.
439
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Glucocorticoides
Aumentan la glucemia (inverso a la insulina). En hígado promueve la captación
de aminoácidos y glucosa, la síntesis de glucosa a partir de aminoácidos, la
gluconeogénesis y la glucogenolisis.
Estimula la degradación proteica y la movilización de ácidos grasos en
adipocitos.
Tiene efecto antiestrés, contribuye a combatir situaciones físicas y psíquicas de
estrés (infecciones, temperaturas extremas, cirugías...).
Además, tiene efecto antinflamatorio suprime las respuestas tisulares a la lesión
y reduce la inmunidad celular y humoral. Inhibe la colagenasa, la síntesis de
leucotrienos y la proliferación linfocitaria. Se utiliza como tratamiento en la artritis
reumatoide, el asma, alergias y en caso de reacciones inmunológicas en trasplantes.
Hormonas sexuales (DHEA y androstendiona)
440
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
MÉDULA
No se observa un límite nítido de la médula con la corteza. Está constituida por
cordones de células cromafines entre los que encontramos células ganglionares,
capilares fenestrados y vénulas. No tiene capacidad de regeneración.
Células ganglionares
Las células ganglionares son en realidad neuronas simpáticas cuyos axones se
extienden hacia la corteza suprarrenal y modulan ésta y los vasos sanguíneos mediante
la secreción de diversas sustancias. Sin embargo, su bajo número y dispersión hacen
difícil su observación a microscopía.
Células cromafines
Las células cromafines, organizadas en cordones celulares, son grandes y
pálidas. También son nerviosas, puesto que se trata de neuronas postganglionares
simpáticas, pero se diferencian de las ganglionares en que no presentan prolongaciones.
Si se cultivan in vitro, sí las presentan, pero in vivo su aparición se haya inhibida por los
glucocorticoides segregados por la corteza suprarrenal, de modo que si querremos
realizar el experimento con un cultivo in vitro de inyectar glucocorticoides, el
crecimiento de las prolongaciones se inhibirá.
Las células cromafines se encargan de la síntesis, el almacenamiento y la
secreción de catecolaminas (que van unidas a cromograninas). Su RER y su Golgi están
muy desarrollados y presenten vesículas citoplásmicas y gránulos de dos tipos:
x Con norepinefrina (NA/NE): De núcleo denso y excéntrico.
x Con epinefrina (E/A): De núcleo menos denso, más pequeños.
La noradrenalina se sintetiza en el interior del gránulo gracias a la dopamina-
beta-hirdoxilasa, y tras ello sale al citosol, donde puede ser convertida en adrenalina
gracias a la feniletanoamina N-metiltransferasa o PNMT, cuya síntesis es activada por
los glucocorticoides sanguíneos procedentes de la corteza suprarrenal. Una vez
sintetizada, la adrenalina vuelve a entrar en los gránulos en los que se almacena. Este
proceso se puede inhibir mediante reserpina, un fármaco que impide el transporte de
catecolaminas.
Del total de células cromafines, un 80% son ricas en adrenalina, y un 20% en
noradrenalina, aunque esta última actúe como precursora de la primera. Las
catecolaminas se segregan a la sangre, y también se produce a este nivel secreción de
dopamina y se cree que de encefalinas.
Cuando adrenalina y nordrenalina se degradan, se forma ácido vanilmandélico y
metanefrina, que se excretan por la orina: cuanto mayor sea la producción de
catecolaminas mayor será su degradación y, por tanto, estos metabolitos tienen
importancia clínica.
441
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Clínica: feocromitoma
Es una hipertensión mantenida o episódica por tumor en la médula suprarrenal,
que conlleva un aumento de ácido vanilmandélico en la orina. Si está asociado a otro
tumor endocrino hablamos de síndrome de neoplasia endocrina múltiple.
442
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Regulación
De ella se encarga el SNS, de manera que si se seccionan sus fibras cesa la
actividad secretora medular. Las fibras preganglionares simpáticas sinaptan con las
cromafines, que son las posganglionares, e inducen la liberación de catecolaminas.
Funciones de las catecolaminas
Capacitan al organismo para afrontar situaciones catastróficas, emergencias
generadas por estrés físico o psíquico. Es decir, crean un ambiente para que la energía
se pueda usar al máximo para afrontar un esfuerzo físico máximo.
Se considera que el verdadero producto de la médula suprarrenal es adrenalina, y
noradrenalina sería solo su precursor. Si extirpásemos la glándula los niveles de
adrenalina se reducirían a cero, pero los de noradrenalina no bajarían, puesto que se
origina también en las terminales axónicas adrenérgicas del sistema vascular.
La secreción basal de adrenalina es limitada, pero aumenta en hipoglucemia,
traumatismo, ejercicio físico, tras intervención quirúrgica…
x Estimula la gluconeogénesis hepática y la lipolisis, aumentando la glucemia y la
concentración plasmática de ácidos grasos.
x Receptores alfa adrenérgicos de miocitos lisos vasculares: vasoconstricción.
x Receptores beta-adrenérgicos: vasodilatación (miocitos estriados) y
broncodilatación (miocitos lisos de vías aéreas).
x Aumenta la tensión arterial, la frecuencia respiratoria, la frecuencia cardiaca y
Q.
x Vasodilatación coronaria y esquelética.
x Vasoconstricción en piel y tubo digestivo.
Vascularización
Del plexo capsular compuesto por arterias suprarrenales superiores (de frénica
inferior), medias (de aorta) e inferiores (de renal) salen arteriolas:
x Que irrigan cápsula.
x Corticales: Dan capilares fenestrados rectos o sinusoides que forman una red en
la zona reticular y drenan por venas colectoras situadas en el límite
corticomedular (pues en corteza no hay venas ni linfáticos).
x Medulares: Forman una red capilar fenestrada que frena por las venas central y
suprarrenal, pero no irrigan la corteza. En la médula, las venas presentan
engrosamientos conocidos como almohadillas y provocados por el desarrollo de
miocitos lisos en disposición circular y longitudinal.
La médula presenta una doble irrigación, puesto que así le llega sangre con
hormonas corticales como el cortisol que inducen la producción de catecolaminas en
respuesta al estrés.
443
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
444
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
PÁNCREAS ENDOCRINO
Es una glándula aneja al aparato digestivo. Posee unas pequeñas zonas dispersas
que corresponden al páncreas endocrino (2% del páncreas), más abundantes en la zona
de la cola.
Las células endocrinas se desarrollan aquí entre las semanas 9-12. Cada una de
las masas se denomina islotes de Langerhans, siendo de tamaño y cantidad de células
muy variables. Forman cordones muy ramificados entre los cuales hay vasos. Además,
cada islote se delimita por finas fibras reticulares que también penetran hacia el interior.
Estas células responden a estímulos nerviosos (fibras simpáticas y parasimpáticas).
Así, en cada islote podemos reconocer fibras del Sistema Nervioso Autónomo,
tanto simpáticas como parasimpáticas. Además, como se ha mencionado anteriormente,
entre la masa exocrina hay no sólo vasos, sino también grupos celulares como células
endocrinas.
445
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
SISTEMA PORTA ÍNSULO-ACINAR
Consiste en una red capilar fenestrada. En cada islote tenemos una arteriola
aferente (entra en el islote) que se capilariza, con capilares fenestrados. La sangre que
entra llega primero a la zona periférica del islote, y luego llega a la central. En la zona
central hay células B, y cuando la sangre las alcanza, ya ha recogido las hormonas
segregadas por las células de la periferia (principalmente A y D). La sangre se drena por
una vénula, que sale del islote y se mete en la porción exocrina, donde se vuelve a
capilarizar (la vénula es un vaso corto).
La porción exocrina (donde se
encuentra el segundo plexo capilar)
tiene doble vascularización, con una red
capilar que irriga directamente a los
acinos, y otra que proviene del islote.
Así, hay un aporte sanguíneo mayor, y
el sistema porta asegura que las
hormonas segregadas en los islotes
actúen en el propio páncreas (local).
Hay unas hormonas (insulina,
VIP y CCK), que estimulan la secreción
exocrina; mientras que glucagón, PP
(polipéptido pancreático) y
somatostatina inhiben dicha secreción y
proceden de Langerhans.
TIPOS CELULARES
Hay cuatro tipos de células:
x Las células A o α producen
glucagón.
x Las células B o β producen
insulina.
x Las células D o δ producen
gastrina y somatostatina.
x Las células F producen PP.
Los tipos celulares se reconocen al MET o mediante inmunocitoquímica. Con el
tricrómico de Mallory-Azan se distinguen los tres primeros tipos. Con esta tinción:
x Las A se tiñen de rojo, y son periféricas. Sus gránulos y vesículas son visibles a
MET. Sus gránulos son los más pequeños que vamos a poder ver.
x Las B se tiñen de color más pardo/ocre, y son centrales. Sus gránulos presentan
un tamaño mayor que los de glucagón.
x Las D se tiñen en azul, y son periféricas. Los gránulos de las D son los más
grandes.
446
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
Los gránulos son diferentes: los más grandes son los de las D y su matriz es
homogénea, mientras que los de las A y las B son más pequeñas con un centro denso y
un halo periférico claro. En el caso de las A, esta zona densa está en un polo excéntrico,
mientras que en las B tenemos un núcleo más centrado con una estructura cristalina o
paracristalina. Hay otras células en el islote, pero son minoritarias.
Todas estas células del islote existen también fuera del páncreas, excepto las B,
que solo existen en los islotes pancreáticos. Así, las células insulares (excepto las B),
son equivalentes a las células endocrinas de la mucosa del tubo digestivo. Y, si se daña
el páncreas o las células B, no hay células similares que puedan suplir dicho daño.
Todas las células del islote se unen entre sí por desmosomas, Zonula Occludens
o uniones tipo GAP. Lo islotes se generan a lo largo de la vida y por ello se van
formando nuevas células a partir de los conductos excretores, que son de la porción
exocrina. Esos conductos forman un epitelio de revestimiento, a partir de cuyas células
se pueden formar nuevas células endocrinas que migran al islote.
Células A o α
Las células alfa es una población que supone cerca del 20% de las células
endocrinas de los islotes, y se sitúan en la periferia de los mismos; aunque no todas las
células de la periferia son A. Tienen gránulos de tamaño no muy pequeño pero tampoco
grande, con un centro denso homogéneo y un halo periférico claro. Son muy
abundantes.
Producen fundamentalmente glucagón pero también dicolecistoquinina (CCK),
GIP y ACTH-endorfina. La hormona principal que sintetizan es el glucagón a partir de
un precursor (preproglucagón, sintetizado en el RER). Las vesículas que contienen el
glucagón se fusionan con la membrana plasmática y lo liberan. Son estimuladas por los
niveles bajos de glucemia, o los altos de arginina y alanina, y la inervación simpática.
No todo el glucagón es sintetizado por estas células, también lo hace el tubo
digestivo y el encéfalo. El glucagón circulante ejerce sus efectos a todos los órganos. El
principal órgano donde se degrada es el hígado. Éste provoca la glucógenolisis,
conduciendo a elevar los niveles de glucemia (hiperglucemia).
Células B o β
Son las mayoritarias de las células endocrinas (70%). Ocupan la zona central de
los islotes. Poseen gránulos muy abundantes y ligeramente mayores que los de las
células A, con un núcleo denso. Tienen halo periférico claro pero el centro no es denso
homogéneo, sino que se observa la insulina cristalizada. La cantidad de células B se
ajusta al 3% diariamente por mitosis para ajustar su masa a los requerimientos según la
ingesta y la metabolización de la glucosa, habiendo un ajuste diario.
Los niveles elevados de glucemia (hiperglucemia) provocan que las células B no
solo sinteticen insulina, sino que también la secreten. Así, el glucagón también estimula
a estas células B.
447
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
La insulina es sintetizada por un precursor y a partir de ahí se metabolizará. Esta
hormona presenta dos cadenas que están unidas por el péptido C. Al producirse la
escisión del péptido C se quedan libres las dos cadenas de insulina.
Hay que recalcar el hecho de que dentro de la vesícula que va a descargar
insulina, tenemos un núcleo central denso que se le denomina cristaloide de insulina.
Para formar dicho cristaloide son necesarios niveles elevados de Zinc.
Para que se dé la descarga de estas vesículas hay que estimular las células B. En
este caso necesitamos altos niveles de glucosa.
Cuando estas células son estimuladas, una molécula que actúa como canal
(GLUT2) comienza a captar glucosa. Como consecuencia, los niveles de glucosa bajan
porque están entrando en la célula. Los canales de potasio se cierran, por tanto, también
se acumula K+. Esta cascada acaba con la apertura de canales de calcio voltaje-
dependientes. Los gránulos que contienen insulina, después de todo esto, se vierten por
exocitosis.
Las células B también son estimuladas por el GLP-1 o el GIP (péptido inhibidor
gástrico, secretado por las células K del tubo digestivo), pero no son estímulos tan
potentes como los gránulos de Nissl de glucosa. La inervación parasimpática es otro
estimulador. Por el contrario, las catecolaminas (E y NE) inhiben la liberación de
insulina.
448
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
La insulina actúa sobre determinadas células, haciendo que el GLUT4 pase a
formar parte de su membrana plasmática (la tienen internalizada en vesículas del Golgi).
Con el estímulo, GLUT4 pasa a estar en la membrana plasmática, actuando como un
canal que permite la entrada de glucosa. Ocurre en adipocitos, miocitos cardiacos y
esqueléticos, fibroblastos y hepatocitos. Induce a hipoglucemia. En los adipocitos, la
glucosa se desvía hacia la síntesis de triglicéridos. En hepatocitos y miocitos, está
implicada en la formación de glucógeno (glucogenogénesis).
Clínica: Diabetes Mellitus
La diabetes mellitus es un
término que agrupa diferentes
trastornos metabólicos, en los que
el paciente tiene niveles elevados
de glucosa. Diferentes situaciones
pueden llevar a esta hiperglucemia
(problemas de secreción, de
insulina...) y producir también
glucosuria. Como consecuencia, las
paredes arteriales están afectadas,
comienza un depósito de ateroma.
También se ve afectado el sistema
microvascular, habiendo una mayor
permeabilidad en la zona capilar.
Las láminas basales (vasculares o
no) están más engrosadas. Los
órganos más comprometidos en
personas diabéticas son la retina,
los riñones y los miembros
inferiores.
Hay dos grandes grupos, diabetes I y II.
x La diabetes mellitus tipo I se debe a un proceso autoinmune.
x En la diabetes mellitus tipo II tenemos agotamiento de células B (desaparecen).
449
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
En el caso de las mujeres diabéticas, sus hijos tienen mayor posibilidad de
padecer sobrepeso y presentar una hiperplasia de estos islotes (mayor tamaño y número
de islotes). La diabetes gravídica se produce cuando en la mujer gestante la masa de
células B aumenta, siendo la mujer no diabética, pero durante la gestación lo es.
Tenemos también diabetes en las que hay mutaciones en el ADN mitocondrial,
aunque es poco frecuente. Hay un mayor número de pacientes en los que estos defectos
génicos son dominantes, aunque no implica necesariamente que tengan que padecer
obesidad.
Hay diabetes que afectan a recién nacidos, la diabetes mellitus neonatal, y la
hipoglucemia hiperinsulinémica neonatal. En ambos casos las mutaciones están en los
mismos genes, SUR 1 y KIR 6.2.
En la diabetes mellitus neonatal, los genes, al estar mutados, provocan que los
canales de K estén permanentemente abiertos, por lo que se da una reducción drástica
en la secreción de insulina, y por tanto padecen hiperglucemia.
En la hipoglucemia hiperinsulinémica neonatal, estos mismos genes están
mutados, pero tiene como consecuencia que los canales de K estén permanentemente
cerrados. En esta situación, tenemos hipersecreción de insulina no regulada.
También podemos tener situaciones de hiperinsulinema por tumores de células B
(insulinoma).
Células D o δ
Son menos abundantes (10%), están en la periferia del islote, presentan gránulos
de secreción de mayor calibre y tienen matriz homogénea, sin halo periférico claro. Los
mismos factores que regulan las B regulan a estas células D. Sintetizan y segregan
somatostatina y gastrina.
La somatostatina es inhibidora y actúa en los propios islotes inhibiendo al resto
de células (alfa, beta...). La somatostatina pancreática es la misma que la gástrica. Así,
inhibe las células oxínticas, las endocrinas del estómago (y por tanto la secreción de
gastrina), y las exocrinas del páncreas. También inhibe la secreción de GH hipofisaria.
Células F
Las células F son sólo un 2%, sintetizan y segregan el polipéptido P (PP), que es
inhibidor, inhibe la secreción de enzimas pancreáticas. Inhibe la acción de la
somatostatina y la contracción de la vesícula biliar y del páncreas.
450
Descargado por alberto gallego (aryal020202@gmail.com)
lOMoARcPSD|4316946
451
Descargado por alberto gallego (aryal020202@gmail.com)
También podría gustarte
- Los Senos ParanasalesDocumento2 páginasLos Senos ParanasalesJohan David SalazarAún no hay calificaciones
- Anatomia de Las Vias AereasDocumento5 páginasAnatomia de Las Vias Aereasperla batista carrascoAún no hay calificaciones
- Semiologia y NeumologiaDocumento37 páginasSemiologia y NeumologiaOmar ZapataAún no hay calificaciones
- Ruta de La Vida 4 DemorasDocumento44 páginasRuta de La Vida 4 DemorasMario Fernando Villota Esparza0% (1)
- Embriologia Piel Cabeza y CuelloDocumento45 páginasEmbriologia Piel Cabeza y CuelloGabriel Alejandro FloresAún no hay calificaciones
- Embriología Orientada A Sistemas Musculo Esqueletico 2Documento77 páginasEmbriología Orientada A Sistemas Musculo Esqueletico 2Samuel Fernando Velásco ChacónAún no hay calificaciones
- Médula ósea y desarrollo del esqueleto humanoDocumento23 páginasMédula ósea y desarrollo del esqueleto humanoEdu Hernandez Ramirez100% (1)
- Onfalocele WordDocumento11 páginasOnfalocele WordmariajoseAún no hay calificaciones
- Cristalino y Luxacion-2Documento17 páginasCristalino y Luxacion-2gabriela lucasAún no hay calificaciones
- CLASE #1 ORL ANATOMIA DE OIDO, NARIZ, FARINGE y LARINGEDocumento7 páginasCLASE #1 ORL ANATOMIA DE OIDO, NARIZ, FARINGE y LARINGEluce netAún no hay calificaciones
- Enfermedad Pulmonar Intersticial DisfusaDocumento10 páginasEnfermedad Pulmonar Intersticial DisfusadrheayAún no hay calificaciones
- Cataratas: Causas, síntomas y tratamiento de la opacidad del cristalinoDocumento12 páginasCataratas: Causas, síntomas y tratamiento de la opacidad del cristalinoGenesis Orozco100% (1)
- Características y Funciones de Los Ganglios BasalesDocumento1 páginaCaracterísticas y Funciones de Los Ganglios BasalesMaydi Jorleny Alvizures BarahonaAún no hay calificaciones
- PrepucioDocumento12 páginasPrepucioGreismarly AguanaAún no hay calificaciones
- Caso Clinico OtorrinoDocumento9 páginasCaso Clinico OtorrinoMaria Del Cisne Solorzano100% (1)
- L.C. CRÓNICA: CAUSAS, SÍNTOMAS Y TRATAMIENTODocumento15 páginasL.C. CRÓNICA: CAUSAS, SÍNTOMAS Y TRATAMIENTOJhulissa arias cruzAún no hay calificaciones
- Tema 3 Meninges PDFDocumento8 páginasTema 3 Meninges PDFCristian Camilo HerreraAún no hay calificaciones
- Tetralogía de Fallot & Coartación de La AortaDocumento19 páginasTetralogía de Fallot & Coartación de La AortaRodrigo Nuñez MendezAún no hay calificaciones
- Sesion 48 Enfermedades Infecciosas Del Aparato DigestivoDocumento29 páginasSesion 48 Enfermedades Infecciosas Del Aparato DigestivoPacko Guerrero CastroAún no hay calificaciones
- Embriologia de Aparato Genital y GenitourinarioDocumento102 páginasEmbriologia de Aparato Genital y GenitourinarioNain Oliva LozanoAún no hay calificaciones
- Banqueo Extremo-HistoDocumento18 páginasBanqueo Extremo-HistoNicole GarcíaAún no hay calificaciones
- Pruebas de Weber y Rinne evalúan hipoacusiasDocumento14 páginasPruebas de Weber y Rinne evalúan hipoacusiasDon PendejazoAún no hay calificaciones
- Patologías de Los Ganglios LinfáticosDocumento19 páginasPatologías de Los Ganglios LinfáticosPatricia ArteagaAún no hay calificaciones
- Defectos del tubo neural: causas, diagnóstico y tratamientoDocumento38 páginasDefectos del tubo neural: causas, diagnóstico y tratamientoBeatriz Kazuko DelaHerránAún no hay calificaciones
- Anillo de Waldeyer PDFDocumento3 páginasAnillo de Waldeyer PDFGuadalupe García FigueroaAún no hay calificaciones
- Semiologia de Los OidosDocumento17 páginasSemiologia de Los Oidosana caren carrasco guereca100% (1)
- Desarrollo Anatómico Del OjoDocumento38 páginasDesarrollo Anatómico Del OjoClaudi CallejoAún no hay calificaciones
- Intertrigo Gineco-ObstetriciaDocumento33 páginasIntertrigo Gineco-Obstetriciaangelica_10695882Aún no hay calificaciones
- Patología de Válvulas Auriculo VentricularesDocumento47 páginasPatología de Válvulas Auriculo VentricularesGabriela MoralesAún no hay calificaciones
- Vías LagrimalesDocumento21 páginasVías LagrimalesAngie EncarnacionAún no hay calificaciones
- 1 - Fisiopatologia Del Trauma HuancayoDocumento51 páginas1 - Fisiopatologia Del Trauma HuancayoJoel Alderete CamposAún no hay calificaciones
- Malformaciones Del TimoDocumento6 páginasMalformaciones Del TimoNaomi CarreraAún no hay calificaciones
- Caso01 Embriologia 1Documento11 páginasCaso01 Embriologia 1Nicolle Aquino ThorneAún no hay calificaciones
- Malformaciones de Arcos FaringeosDocumento5 páginasMalformaciones de Arcos FaringeosZeak & TiankyAún no hay calificaciones
- J ctvtxw3sz 21Documento23 páginasJ ctvtxw3sz 21Oli QuiñonesAún no hay calificaciones
- Semiologia de Cabeza y CuelloDocumento46 páginasSemiologia de Cabeza y CuelloAndres MarquezAún no hay calificaciones
- Histologia Expo NarizDocumento17 páginasHistologia Expo NarizSeba Deza Bellido100% (1)
- 04 Esqueleto AxialDocumento123 páginas04 Esqueleto AxialMario Salini100% (1)
- Embriologia Del Sistema CardiovascularDocumento25 páginasEmbriologia Del Sistema CardiovascularLeyla PerezAún no hay calificaciones
- Simulacro Parte ADocumento23 páginasSimulacro Parte AEussk AdyAún no hay calificaciones
- Malformaciones Congénitas Del OjoDocumento3 páginasMalformaciones Congénitas Del OjoPAULINA DE LOS SANTOS ZAMBRANOAún no hay calificaciones
- Anatomía Oído InternoDocumento14 páginasAnatomía Oído InternoipfonoaudiologiaAún no hay calificaciones
- Manifestaciones Clinicas - Homocistinuria (Actualizado)Documento3 páginasManifestaciones Clinicas - Homocistinuria (Actualizado)Maria Almanza SarmientoAún no hay calificaciones
- Apuntes 1 - OtorrinolaringologíaDocumento70 páginasApuntes 1 - OtorrinolaringologíaElizabeth PeñaAún no hay calificaciones
- RinosinusitisDocumento12 páginasRinosinusitisana trocelAún no hay calificaciones
- Irrigacion ApendiceDocumento7 páginasIrrigacion ApendiceValenty VillanuevaAún no hay calificaciones
- Tromboembolia PulmonarDocumento24 páginasTromboembolia PulmonarAnnia Valdivia100% (1)
- Síndromes Parenquimatosos PulmonaresDocumento11 páginasSíndromes Parenquimatosos PulmonaresRicardo Patricio Acuña JaramilloAún no hay calificaciones
- Desarrollo Embrionario de La VisionDocumento22 páginasDesarrollo Embrionario de La VisionWinny Culqui ManosalvaAún no hay calificaciones
- Síndrome de Costen: causas, síntomas y tratamientoDocumento15 páginasSíndrome de Costen: causas, síntomas y tratamientoJosé Alvaro Rivera AvitiaAún no hay calificaciones
- Guía esplenectomíaDocumento43 páginasGuía esplenectomíaGaby UzcateguiAún no hay calificaciones
- Clase 12 Fisiologia (Ppt12)Documento12 páginasClase 12 Fisiologia (Ppt12)manuAún no hay calificaciones
- PDF-Enfermedades de La Esclera y Conjuntiva-WordDocumento20 páginasPDF-Enfermedades de La Esclera y Conjuntiva-WordKabuki SamAún no hay calificaciones
- SD de CondensacionDocumento5 páginasSD de CondensacionHugo Arancibia EspinozaAún no hay calificaciones
- Neurofisiología laríngea: receptores sensoriales y funcionesDocumento34 páginasNeurofisiología laríngea: receptores sensoriales y funcionesnefimdAún no hay calificaciones
- Resumen Cabeza y CuelloDocumento73 páginasResumen Cabeza y CuelloDaniela Gonzales100% (1)
- Tejido Oseo y Cartilaginoso PDFDocumento63 páginasTejido Oseo y Cartilaginoso PDFAna RomeroAún no hay calificaciones
- Síndrome de UsherDocumento15 páginasSíndrome de UsherGalith BarrientosAún no hay calificaciones
- 7.0 Clase Sist RespiratorioDocumento47 páginas7.0 Clase Sist RespiratorioDaniel Loyola AburtoAún no hay calificaciones
- ANESTESIADocumento60 páginasANESTESIAALBERTO GALLEGOAún no hay calificaciones
- Leccion 2Documento15 páginasLeccion 2ALBERTO GALLEGOAún no hay calificaciones
- Hipospadia FinalDocumento8 páginasHipospadia FinalALBERTO GALLEGOAún no hay calificaciones
- Lecciones de TeoríaDocumento2 páginasLecciones de TeoríaALBERTO GALLEGOAún no hay calificaciones
- Sindrome de BalintDocumento4 páginasSindrome de BalintALBERTO GALLEGOAún no hay calificaciones
- ANATOMÍA II (2014-2015) ConsensoDocumento9 páginasANATOMÍA II (2014-2015) ConsensoALBERTO GALLEGOAún no hay calificaciones
- Desarrollo Embrionario Del Tendon Cabeza Larga Biceps BraquialDocumento16 páginasDesarrollo Embrionario Del Tendon Cabeza Larga Biceps BraquialALBERTO GALLEGOAún no hay calificaciones
- Sindrome de Wallenberg2Documento5 páginasSindrome de Wallenberg2ALBERTO GALLEGOAún no hay calificaciones
- The Cerebellum and Cognitive Function 25 AñosDocumento5 páginasThe Cerebellum and Cognitive Function 25 AñosALBERTO GALLEGOAún no hay calificaciones
- Fractura Diáfisis Humeral Maria Jesus, Ana, Emilia, Laura y CarlosDocumento16 páginasFractura Diáfisis Humeral Maria Jesus, Ana, Emilia, Laura y CarlosALBERTO GALLEGOAún no hay calificaciones
- ANATOMÍA II (2014-2015) ConsensoDocumento9 páginasANATOMÍA II (2014-2015) ConsensoALBERTO GALLEGOAún no hay calificaciones
- El HermunculusDocumento16 páginasEl HermunculusALBERTO GALLEGOAún no hay calificaciones
- Trabajo Realizado Por: Laura Mazano Lozano Laura Peñate Pérez Natalia Naranjo GuerreroDocumento19 páginasTrabajo Realizado Por: Laura Mazano Lozano Laura Peñate Pérez Natalia Naranjo GuerreroALBERTO GALLEGOAún no hay calificaciones
- Tunel CarpianoDocumento18 páginasTunel CarpianoALBERTO GALLEGOAún no hay calificaciones
- ROTURA DEL TENDÓN DE AQUILES JmavDocumento8 páginasROTURA DEL TENDÓN DE AQUILES JmavALBERTO GALLEGOAún no hay calificaciones
- LUMBALGIADocumento22 páginasLUMBALGIAALBERTO GALLEGOAún no hay calificaciones
- Caso Clínico Anatomia - TRIADA DESGRACIADADocumento10 páginasCaso Clínico Anatomia - TRIADA DESGRACIADAALBERTO GALLEGOAún no hay calificaciones
- 4 Practica VB EspecilizDocumento3 páginas4 Practica VB EspecilizALBERTO GALLEGOAún no hay calificaciones
- Fractura de La Diáfisis Humeral - Maria, Olga, Ana, Carmen, Sole.Documento16 páginasFractura de La Diáfisis Humeral - Maria, Olga, Ana, Carmen, Sole.ALBERTO GALLEGOAún no hay calificaciones
- Practica 17Documento2 páginasPractica 17ALBERTO GALLEGOAún no hay calificaciones
- Practica 20Documento2 páginasPractica 20ALBERTO GALLEGOAún no hay calificaciones
- Arteria - Poplitea Guillermo Jaime y CarlosDocumento13 páginasArteria - Poplitea Guillermo Jaime y CarlosALBERTO GALLEGOAún no hay calificaciones
- Practica 6bDocumento1 páginaPractica 6bALBERTO GALLEGOAún no hay calificaciones
- Practica 18Documento3 páginasPractica 18ALBERTO GALLEGOAún no hay calificaciones
- 5 Practicacostillas EsternonDocumento2 páginas5 Practicacostillas EsternonALBERTO GALLEGOAún no hay calificaciones
- 3 Practica VB GnralesDocumento2 páginas3 Practica VB GnralesALBERTO GALLEGOAún no hay calificaciones
- Practica 21Documento3 páginasPractica 21ALBERTO GALLEGOAún no hay calificaciones
- 1 Caracteristicas VBDocumento1 página1 Caracteristicas VBALBERTO GALLEGOAún no hay calificaciones
- ClavículaDocumento1 páginaClavículaALBERTO GALLEGOAún no hay calificaciones
- Practica 1bDocumento3 páginasPractica 1bALBERTO GALLEGOAún no hay calificaciones
- Histamina - Funciones y Trastornos AsociadosDocumento13 páginasHistamina - Funciones y Trastornos AsociadosYenifer Yslas VillegasAún no hay calificaciones
- Ciclo CelularDocumento14 páginasCiclo CelularAlvaro AceroAún no hay calificaciones
- Síndrome Psicopático BasicoDocumento10 páginasSíndrome Psicopático BasicoFiorella SantosAún no hay calificaciones
- Costos de Los Accidentes de TrabajoDocumento12 páginasCostos de Los Accidentes de TrabajoRosa Marselle Sanchez BeltreAún no hay calificaciones
- Exposición FármacoDocumento5 páginasExposición FármacoCarolina DiazAún no hay calificaciones
- Cuadro ComparativoDocumento3 páginasCuadro Comparativooscar patioAún no hay calificaciones
- Manual 2012 Parte 1Documento87 páginasManual 2012 Parte 1Adriana Beatriz MirandaAún no hay calificaciones
- Aceptacion Consentimiento LigaDocumento1 páginaAceptacion Consentimiento LigaGustavo Adolfo Torres CastilloAún no hay calificaciones
- Dengue 2Documento64 páginasDengue 2Maria Alejandra GrisalesAún no hay calificaciones
- El Cuaderno de Porque Biotecnologia Nro 35 Cultivo in VitroDocumento13 páginasEl Cuaderno de Porque Biotecnologia Nro 35 Cultivo in VitrogunzhotAún no hay calificaciones
- EgosintónicoDocumento2 páginasEgosintónicoCarmela PerezAún no hay calificaciones
- ZoonosisDocumento44 páginasZoonosisWalther Lazarus CH FAún no hay calificaciones
- Triptico VbilDocumento18 páginasTriptico VbilVil Aguilar100% (1)
- Taller de Ventilacion Fisiologia RespiratoriaDocumento57 páginasTaller de Ventilacion Fisiologia RespiratoriavalenzuelacastilloAún no hay calificaciones
- Mapa Conceptual Intestino Medio y PosteriorDocumento4 páginasMapa Conceptual Intestino Medio y PosteriorMabel Ramirez0% (1)
- Semiología de CuelloDocumento7 páginasSemiología de CuellodaniAún no hay calificaciones
- Guia Paciente Renal PDFDocumento24 páginasGuia Paciente Renal PDFRomeo Lima BravoAún no hay calificaciones
- Fisiopatología de La DislipemiaDocumento5 páginasFisiopatología de La DislipemiaKarla Quiñones RufinoAún no hay calificaciones
- Protocolos MMSDocumento9 páginasProtocolos MMSm0ra0Aún no hay calificaciones
- Diagnostico de SaludDocumento33 páginasDiagnostico de SaludMario Avila100% (1)
- La Gatera de La Villa 1Documento20 páginasLa Gatera de La Villa 1Vicente Regulez FernandezAún no hay calificaciones
- Descripción de Algunas de Las Flores de BachDocumento1 páginaDescripción de Algunas de Las Flores de BachEduardo Chavez DuqueAún no hay calificaciones
- Tarea BioeticaDocumento5 páginasTarea BioeticaEduardo EstrellaAún no hay calificaciones
- Libro Microbg 1Documento83 páginasLibro Microbg 1Cristhian0% (1)
- El Psicodiagnóstico Clínico en La ActualidadDocumento10 páginasEl Psicodiagnóstico Clínico en La ActualidadThanny AlvarezAún no hay calificaciones
- Borderline Personalidad Al Borde de La LocuraDocumento3 páginasBorderline Personalidad Al Borde de La LocuraConsuelo GarciaAún no hay calificaciones
- Metodos Diagnosticos CardioDocumento1 páginaMetodos Diagnosticos Cardioapi-472537857Aún no hay calificaciones
- Sacha InchiDocumento18 páginasSacha InchiJOEAún no hay calificaciones
- Aclaraciones Coronariografia y Angioplastia CoronariaDocumento3 páginasAclaraciones Coronariografia y Angioplastia Coronariatop10uandes100% (2)
- Una Vida Saludable - EssayDocumento1 páginaUna Vida Saludable - EssayPrinss GonzalezAún no hay calificaciones