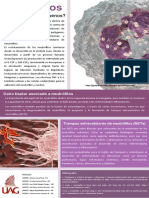
También podría gustarte
- Trampas Extracelulares de Neutrofilos Un Mecanismo de Defensa Con Dos CarasDocumento7 páginasTrampas Extracelulares de Neutrofilos Un Mecanismo de Defensa Con Dos Caraslucia mayancelaAún no hay calificaciones
- Agilizar La Creación de VacunasDocumento6 páginasAgilizar La Creación de VacunasJaviera GonzalezAún no hay calificaciones
- Cuestionario de La Práctica de ViroDocumento4 páginasCuestionario de La Práctica de ViroMaiela FrikitonaAún no hay calificaciones
- TRAPAS EXTRACELULARES DEL NEUTROFILO (NETDocumento2 páginasTRAPAS EXTRACELULARES DEL NEUTROFILO (NETSebastián Aguilera HernándezAún no hay calificaciones
- 1.4 Trampas Extracelulares Del NeutrófiloDocumento2 páginas1.4 Trampas Extracelulares Del NeutrófiloJulio Daniel González GarcíaAún no hay calificaciones
- NET'sDocumento8 páginasNET'sNOE HERMINIO VELAZQUEZ RECINOSAún no hay calificaciones
- Leer Primero. Revisión en Términos Generales EsDocumento16 páginasLeer Primero. Revisión en Términos Generales EsChina ReyesAún no hay calificaciones
- Interpretacion ClinicaDocumento12 páginasInterpretacion ClinicaMiguel De Jesus Gallegos PeraltaAún no hay calificaciones
- 6 Sim Tip Adm Unmsm SSMDocumento18 páginas6 Sim Tip Adm Unmsm SSMBrayan CqAún no hay calificaciones
- Taller Parasitologia Juan Rozo-Kelly LopezDocumento8 páginasTaller Parasitologia Juan Rozo-Kelly LopezJuan Felipe Rozo MorenoAún no hay calificaciones
- Tejido Inmunitario HistoDocumento23 páginasTejido Inmunitario HistoAlma Rosa OrtizAún no hay calificaciones
- Taller Parasitologia Juan Rozo-Kelly LopezDocumento8 páginasTaller Parasitologia Juan Rozo-Kelly LopezJuan Felipe Rozo MorenoAún no hay calificaciones
- NEUTRÓFILOS - Doménica ViñánDocumento4 páginasNEUTRÓFILOS - Doménica ViñánDomenica ViñanAún no hay calificaciones
- BM PLP1 GarciaCarrilloSofiaGuiselleDocumento6 páginasBM PLP1 GarciaCarrilloSofiaGuiselleSofiia CarrilloAún no hay calificaciones
- NeutrófilosDocumento1 páginaNeutrófilosMiguel Angel Badillo GomezAún no hay calificaciones
- EXPO Respuesta Inmune Frente A Infecciones Bacterianas UCM FinalDocumento53 páginasEXPO Respuesta Inmune Frente A Infecciones Bacterianas UCM FinalJasser Josué Martínez RostránAún no hay calificaciones
- Informe 4. Biología MolecularDocumento13 páginasInforme 4. Biología MolecularAna María Vallejo TorresAún no hay calificaciones
- Sistema inmunologico-DIANA PACHAO - Obstetricia IIIDocumento42 páginasSistema inmunologico-DIANA PACHAO - Obstetricia IIIDiana Elena Pachao MariñoAún no hay calificaciones
- Fisiología Del Sistema InmunológicoDocumento27 páginasFisiología Del Sistema InmunológicoJOSE MANUEL VIDAL VARGASAún no hay calificaciones
- 1ra Lectura. Sistema Inmune InnatoDocumento10 páginas1ra Lectura. Sistema Inmune InnatoTaniaZayuriRosalesOrtizAún no hay calificaciones
- Glosario de términos inmunológicosDocumento7 páginasGlosario de términos inmunológicosanyshaeAún no hay calificaciones
- Células DentríticasDocumento7 páginasCélulas DentríticasNicole Anahi Montes RomeroAún no hay calificaciones
- Guia de Seminarios 2023Documento19 páginasGuia de Seminarios 2023caminaiaraAún no hay calificaciones
- CLASE 3. Inflamación Aguda y Crónica Neoplasias y Metastasis.Documento42 páginasCLASE 3. Inflamación Aguda y Crónica Neoplasias y Metastasis.camilaAún no hay calificaciones
- NeutrófilosDocumento2 páginasNeutrófilosAna MoraAún no hay calificaciones
- Conocimientos Actuales en La Fisiopatología de La Sepsis: C. Ortiz Leyba Y J. Garnacho MonteroDocumento7 páginasConocimientos Actuales en La Fisiopatología de La Sepsis: C. Ortiz Leyba Y J. Garnacho MonteroLeticia ConstantinoAún no hay calificaciones
- Caracteristicas Generales de La Respuesta InmuneDocumento62 páginasCaracteristicas Generales de La Respuesta InmuneAndres RomeroAún no hay calificaciones
- Año de La UnidadDocumento5 páginasAño de La UnidadD-uanerAún no hay calificaciones
- Ensayo. La Respuesta Inmune Innata.Documento7 páginasEnsayo. La Respuesta Inmune Innata.Edgar Omar De La Garza MataAún no hay calificaciones
- Campo de Las Vacunas RecombinantesDocumento2 páginasCampo de Las Vacunas RecombinantesGERARDO SILVESTRE MELENDEZ MACALOPUAún no hay calificaciones
- Neutrofilos NetosisDocumento3 páginasNeutrofilos NetosisalejandraAún no hay calificaciones
- Ensayo Cells at WorkDocumento4 páginasEnsayo Cells at WorkShirley vanessa Vega BedoyaAún no hay calificaciones
- 9.4. Neutrophils (Neutrófilos)Documento1 página9.4. Neutrophils (Neutrófilos)katherineAún no hay calificaciones
- Pract. Calificada 4 Act.02Documento9 páginasPract. Calificada 4 Act.02EdithAún no hay calificaciones
- Trabajo Autonomo #2Documento6 páginasTrabajo Autonomo #2Deniisse Garcia100% (1)
- Generalidades Del Sistema InmunitarioDocumento5 páginasGeneralidades Del Sistema InmunitarioMaria Patricia Lopez SantanaAún no hay calificaciones
- Trabajo de InmunologiaDocumento24 páginasTrabajo de InmunologiaRamona Fidelina Castillo PetitonAún no hay calificaciones
- PreguntasDocumento18 páginasPreguntasVale Prado VargasAún no hay calificaciones
- Inmunología Del Trasplante - GrupoDocumento8 páginasInmunología Del Trasplante - GrupoFreixil S.SAún no hay calificaciones
- Analisis Del Articulo - Unidad 1, 2, y 3Documento10 páginasAnalisis Del Articulo - Unidad 1, 2, y 3Maydei Rodriguez50% (2)
- Opioides y El Sistema InmuneDocumento6 páginasOpioides y El Sistema Inmunedaniel_jose_skateAún no hay calificaciones
- Fagocitosis PDFDocumento7 páginasFagocitosis PDFAna Julia Mero KuffóAún no hay calificaciones
- Evacion de La Respuesta Inmune AntitumoralDocumento10 páginasEvacion de La Respuesta Inmune AntitumoralOlivia BriceñoAún no hay calificaciones
- Principios de FisiologiaDocumento12 páginasPrincipios de FisiologiaMassiel Sofia Sullca MedinaAún no hay calificaciones
- Enzimas+Especiales+Granzimas+y+Abzimas+ +grupo+4Documento33 páginasEnzimas+Especiales+Granzimas+y+Abzimas+ +grupo+4Anypsa Palacios RomanAún no hay calificaciones
- 337113-Texto Del Articulo-185285-2-10-20210414Documento11 páginas337113-Texto Del Articulo-185285-2-10-20210414Danny FaicanAún no hay calificaciones
- Células y Moléculas Del Sistema Inmune InnatoDocumento20 páginasCélulas y Moléculas Del Sistema Inmune InnatoFernando Aponte R.Aún no hay calificaciones
- Recrecimiento Tumoral en RatonesDocumento7 páginasRecrecimiento Tumoral en RatonesJohan Leonardo Arevalo BallesterosAún no hay calificaciones
- ViruelaDocumento12 páginasViruelaNatividadAún no hay calificaciones
- Animales TransgenicosDocumento24 páginasAnimales TransgenicosFrancisco ZúñigaAún no hay calificaciones
- Celulas SanguineasDocumento4 páginasCelulas SanguineasAngie CruzAún no hay calificaciones
- Capítulo 8Documento3 páginasCapítulo 8Laura UzumakiAún no hay calificaciones
- Sistema Inmunitario Texto HistologiaDocumento18 páginasSistema Inmunitario Texto HistologiaPaloma Mamani JacoboAún no hay calificaciones
- LECTURA 1 - Josse Et Al 2017. Staphylococcal Adhesion and Host Cell Invasion Fibronectin-Binding and Other Mechanisms - En.esDocumento8 páginasLECTURA 1 - Josse Et Al 2017. Staphylococcal Adhesion and Host Cell Invasion Fibronectin-Binding and Other Mechanisms - En.esEctor Adrian Rivadeneyra CardenasAún no hay calificaciones
- CAPÍTULO 1 - Visión General Del Sistema InmuneDocumento42 páginasCAPÍTULO 1 - Visión General Del Sistema InmuneabbyAún no hay calificaciones
- ANEL A - InmunitarioDocumento4 páginasANEL A - InmunitarioEstrella Belen Galo SantiagoAún no hay calificaciones
- Eval Diagnostica Cyt - 1ero y 2do GradoDocumento9 páginasEval Diagnostica Cyt - 1ero y 2do Gradonancy matta guevaraAún no hay calificaciones
- Deber 2 MicroDocumento10 páginasDeber 2 MicroMELANY JELITZA RONQUILLO GARCIAAún no hay calificaciones
- Señalizacion Celular PDFDocumento48 páginasSeñalizacion Celular PDFOscar Ivan0% (1)
- Programa Epidemiologia Lic 64h Ene2017Documento10 páginasPrograma Epidemiologia Lic 64h Ene2017Oscar IvanAún no hay calificaciones
- GuiaDocumento21 páginasGuiaAmelia GibajaAún no hay calificaciones
- Anemia-Inbrief Yg SPDocumento4 páginasAnemia-Inbrief Yg SPAngie RubioAún no hay calificaciones
- Programa Epidemiologia Lic 64h Ene2017Documento10 páginasPrograma Epidemiologia Lic 64h Ene2017Oscar IvanAún no hay calificaciones
- Sindrome de MarfanDocumento20 páginasSindrome de MarfanOscar IvanAún no hay calificaciones
- SolucionesDocumento22 páginasSolucionesOscar IvanAún no hay calificaciones
- Anemia-Inbrief Yg SPDocumento4 páginasAnemia-Inbrief Yg SPAngie RubioAún no hay calificaciones
- Evaluación Del Impacto Ambiental Al Aire Asociado - María T. Galé Z. - usbCTG - 2014Documento76 páginasEvaluación Del Impacto Ambiental Al Aire Asociado - María T. Galé Z. - usbCTG - 2014Oscar IvanAún no hay calificaciones
- Transversales PDFDocumento7 páginasTransversales PDFOscar IvanAún no hay calificaciones
- Practica 03 Cambio GrasoDocumento9 páginasPractica 03 Cambio GrasoOscar IvanAún no hay calificaciones
- Reanimacion Cardiopulmonar BásicaDocumento13 páginasReanimacion Cardiopulmonar BásicaCinthy Jacobo100% (1)
- Diseños de Estudios EpidemiologicosDocumento11 páginasDiseños de Estudios Epidemiologicosapi-3707228100% (1)
- Presentacion 5Documento4 páginasPresentacion 5Oscar IvanAún no hay calificaciones
- Causalidad EpidemiologiaDocumento12 páginasCausalidad EpidemiologiaOscar IvanAún no hay calificaciones
- 21 CausalidadDocumento17 páginas21 CausalidadangieAún no hay calificaciones
- Clase Micro - Corynebacterium.2013Documento25 páginasClase Micro - Corynebacterium.2013Victor Gavidia CaceresAún no hay calificaciones
- Neutrófilos: Células Clave en la Respuesta InflamatoriaDocumento34 páginasNeutrófilos: Células Clave en la Respuesta InflamatoriaDiana PachonAún no hay calificaciones
- Fisiopatología Del AsmaDocumento28 páginasFisiopatología Del AsmamarthaAún no hay calificaciones
- Extremidades BiónicasDocumento16 páginasExtremidades Biónicasasfa weAún no hay calificaciones
- Silabus de Patologia General Unjbg 2019Documento15 páginasSilabus de Patologia General Unjbg 2019Mike CalliAún no hay calificaciones
- Practica 6Documento24 páginasPractica 6victor quispe CastroAún no hay calificaciones
- Lupus Erimatoso Sistemico LES Trabajo Final..........Documento26 páginasLupus Erimatoso Sistemico LES Trabajo Final..........Angelica ZambranoAún no hay calificaciones
- Inmunidad HumoralDocumento7 páginasInmunidad HumoralMilagros FloresAún no hay calificaciones
- GL OsarioDocumento6 páginasGL OsarioCarlos PerezAún no hay calificaciones
- Vacunas Tipos de VacunasDocumento6 páginasVacunas Tipos de VacunasDelicias AL GroneAún no hay calificaciones
- Apuntes de Clases de Células de La Inflamación Aguda y Crónica PDFDocumento18 páginasApuntes de Clases de Células de La Inflamación Aguda y Crónica PDFBoris Dan IllanesAún no hay calificaciones
- 3 C EscarlatinaDocumento8 páginas3 C EscarlatinaMauricio Franco MuñozAún no hay calificaciones
- 2.marco TeoricoDocumento5 páginas2.marco TeoricoJoshimarck Escamilla Lara100% (1)
- Eritema NodosoDocumento24 páginasEritema NodosoThalia LRAún no hay calificaciones
- Conocimiento y aceptación de la vacuna Covid-19 en personal de saludDocumento103 páginasConocimiento y aceptación de la vacuna Covid-19 en personal de saludDelia ChavezAún no hay calificaciones
- Actualización en Sepsis y Choque Séptico: Nuevas Definiciones y Evaluación ClínicaDocumento27 páginasActualización en Sepsis y Choque Séptico: Nuevas Definiciones y Evaluación ClínicaPaola Botello ValenciaAún no hay calificaciones
- 0000008434paaparcana Flores Wilber AntonioDocumento2 páginas0000008434paaparcana Flores Wilber AntoniotatiAún no hay calificaciones
- Autoin Muni DadDocumento49 páginasAutoin Muni Dadanabel martinezAún no hay calificaciones
- Cuidado Del Sistema InmunitarioDocumento2 páginasCuidado Del Sistema InmunitarioMaira Alejandra Garzon MoreraAún no hay calificaciones
- Balotario para Examen Teorico Practico - Enfermeria TecnicaDocumento12 páginasBalotario para Examen Teorico Practico - Enfermeria TecnicaJorge CastillaAún no hay calificaciones
- Muestras Gratis InmunoDocumento45 páginasMuestras Gratis Inmunomicro.fcvunlpAún no hay calificaciones
- Inmunidad Innata UNMSM 2019Documento96 páginasInmunidad Innata UNMSM 2019Alejandro Favio Talavera Arias100% (1)
- Fisiologia I Exposiciones Grupo 1 Tercer Semestre 2023-2024 CiDocumento8 páginasFisiologia I Exposiciones Grupo 1 Tercer Semestre 2023-2024 CiAndreina BarragánAún no hay calificaciones
- Respuesta Inmune NormalDocumento42 páginasRespuesta Inmune NormalSandra Lisseth Ortez CastroAún no hay calificaciones
- TeoríaDocumento148 páginasTeoríalolAún no hay calificaciones
- Sida SeminarioDocumento36 páginasSida Seminario11-MH-HU-DIANA STEFANY MEZA SALCEDOAún no hay calificaciones
- DMSODocumento28 páginasDMSOdrmedrulfoAún no hay calificaciones
- Apa ParasitologiaDocumento57 páginasApa ParasitologiaDiego FigueroaAún no hay calificaciones
- Mecanismos innatos defensa hospederoDocumento2 páginasMecanismos innatos defensa hospederoKarime Avitia ReyesAún no hay calificaciones
- Silabo 2016-2-FinalDocumento17 páginasSilabo 2016-2-Finalalexander_cueva_18Aún no hay calificaciones