También podría gustarte
- Reacción de ArthusDocumento6 páginasReacción de ArthusGloria María100% (2)
- Equipo 8 Factor Estimulante de Colonias Ftp02a 20-2Documento9 páginasEquipo 8 Factor Estimulante de Colonias Ftp02a 20-2Ruth Cobo RosalesAún no hay calificaciones
- Trastornos Hemostasia Primaria y Secundaria. AlgoritmoDocumento2 páginasTrastornos Hemostasia Primaria y Secundaria. AlgoritmoAlex PazAún no hay calificaciones
- Reparación de Tejidos - RobbinsDocumento14 páginasReparación de Tejidos - RobbinsGerson OrellanaAún no hay calificaciones
- Infeccion Por ChancroideDocumento24 páginasInfeccion Por ChancroideClau TomAún no hay calificaciones
- Linfoma Folicular 1Documento11 páginasLinfoma Folicular 1SHEYLA SHARINA FUENTES PINTOAún no hay calificaciones
- Anemias Hemolíticas Extracorpusculares o ExtrínsecasDocumento7 páginasAnemias Hemolíticas Extracorpusculares o ExtrínsecasAndreaRosales0% (1)
- Cuadro Comparativo Tipos de BiopsiaDocumento1 páginaCuadro Comparativo Tipos de BiopsiaJorge Harraka100% (2)
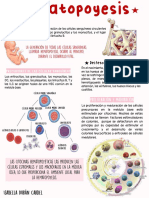
- HematopoyesisDocumento1 páginaHematopoyesisISABELLA DURAN CARDEL100% (1)
- Monografia de Valoracion PreanestesicaDocumento42 páginasMonografia de Valoracion PreanestesicaEspinoza Paola100% (2)
- EstereotiposDocumento2 páginasEstereotiposMarthaMoralesAún no hay calificaciones
- Clasificación PRRDocumento3 páginasClasificación PRRFernando Sanchez MorenoAún no hay calificaciones
- Alteraciones Benignas de Los Leucocitos 1Documento44 páginasAlteraciones Benignas de Los Leucocitos 1Paul Arturo Boza Huaroc100% (1)
- Puntos Dolorosos AbdominalesDocumento6 páginasPuntos Dolorosos AbdominalesLizet nava garciaAún no hay calificaciones
- Artritis SepticaDocumento13 páginasArtritis SepticaAldair Cantillo BarriosAún no hay calificaciones
- Tipos de TóraxDocumento2 páginasTipos de TóraxGabriela Corrales CerdeiraAún no hay calificaciones
- Sdra Volutrauma AtelectraumaDocumento29 páginasSdra Volutrauma Atelectraumaandrexcor100% (1)
- Endocarditis InfecciosaDocumento4 páginasEndocarditis InfecciosaJOSART ARATH CERNAS ORNELASAún no hay calificaciones
- TrombofiliasDocumento27 páginasTrombofiliasAlejandro Jacob IturraAún no hay calificaciones
- Enfermedades Benignas de La MamaDocumento8 páginasEnfermedades Benignas de La MamayaraAún no hay calificaciones
- Semiologia Ganglionar en PPDocumento38 páginasSemiologia Ganglionar en PPnanu01101988Aún no hay calificaciones
- Enfermedades Exantematicas CuadroDocumento3 páginasEnfermedades Exantematicas CuadroJuan Cua100% (2)
- Cascada de CoagulaciónDocumento6 páginasCascada de CoagulaciónSantiago Cruz Quispe0% (1)
- Tamiz NeonatalDocumento29 páginasTamiz NeonatalDonovan Noe Gamez Vega100% (1)
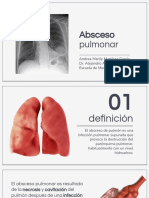
- Absceso PulmonarDocumento32 páginasAbsceso PulmonarAndrea Mtz100% (1)
- Apache 2Documento4 páginasApache 2jamesAún no hay calificaciones
- Sindrome HemorragiparoDocumento50 páginasSindrome HemorragiparoMarco MoralesAún no hay calificaciones
- Enfermedad de Still Del Adulto (ESA)Documento14 páginasEnfermedad de Still Del Adulto (ESA)José Fernando Huanca CordovaAún no hay calificaciones
- Desarrollo y Supervivencia de Linfocitos EmbriologiaDocumento56 páginasDesarrollo y Supervivencia de Linfocitos EmbriologiaSelene Leoon100% (1)
- Sabiston CicatrizaciónDocumento33 páginasSabiston CicatrizaciónEdwin Lionel Castillo MirandaAún no hay calificaciones
- Clasificación de Bethesda de La Citología Cervical.Documento15 páginasClasificación de Bethesda de La Citología Cervical.Dilbeth AlvarezAún no hay calificaciones
- Reaccion Del Centro GerminalDocumento22 páginasReaccion Del Centro GerminalMEDICINA UNIILIBRE 2016-2 GENERALAún no hay calificaciones
- IvuDocumento30 páginasIvuMaria Cristina HuertasAún no hay calificaciones
- 1.a Pancreas Exocrino. Sabiston. Tratado de Cirugia 20a EdicionDocumento12 páginas1.a Pancreas Exocrino. Sabiston. Tratado de Cirugia 20a Edicionmarcela belen100% (1)
- Caso VPHDocumento23 páginasCaso VPHjeancarlosbazaAún no hay calificaciones
- Anemias Cuadro ComparativoDocumento3 páginasAnemias Cuadro ComparativoMiguel Angel Mendoza Cruz100% (10)
- Riñón y Sistema ColectorDocumento32 páginasRiñón y Sistema ColectorIndira BonolaAún no hay calificaciones
- Trabajo de InvestigacionDocumento37 páginasTrabajo de InvestigacionLeslyAgredaNavarroAún no hay calificaciones
- Hipermutacion SomaticaDocumento2 páginasHipermutacion SomaticaIvan Moya50% (2)
- Terapéutica EmpleadaDocumento19 páginasTerapéutica EmpleadaAngelica Cg100% (2)
- Reparación de TejidosDocumento8 páginasReparación de TejidosFernando VillavicencioAún no hay calificaciones
- Resumen de Hematolinfoide de RobbinsDocumento57 páginasResumen de Hematolinfoide de RobbinsMARIA JOSE TORRES ARTEAGAAún no hay calificaciones
- Diagnostico Diferencial Dengue Chikungunya ZikaDocumento4 páginasDiagnostico Diferencial Dengue Chikungunya ZikaBere D LeonAún no hay calificaciones
- La QuimiotaxisDocumento22 páginasLa QuimiotaxisMarisella Chumán Sánchez100% (3)
- Colangitis Aguda PDFDocumento7 páginasColangitis Aguda PDFfruben19Aún no hay calificaciones
- Historia Natural de DERMATOFITOSISDocumento5 páginasHistoria Natural de DERMATOFITOSISSebastian Posadas100% (3)
- DiagnosticoDocumento8 páginasDiagnosticoContrala ApostasíaAún no hay calificaciones
- Estudios de Gabinete y LaboratorioDocumento14 páginasEstudios de Gabinete y LaboratorioDeka Badillo0% (2)
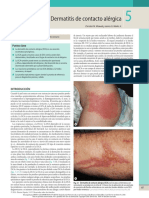
- Dermatitis Contacto AlérgicaDocumento16 páginasDermatitis Contacto Alérgicadiegorojas100% (1)
- Tipos de Inmunidad y Barreras Contra InfeccionesDocumento25 páginasTipos de Inmunidad y Barreras Contra InfeccionesjoseAún no hay calificaciones
- MielopoyesisDocumento9 páginasMielopoyesisLeidy GonzabayAún no hay calificaciones
- STUDOCU Insuficiencia-Cardiaca-Medicina-Interna-De-Harrison-Edicion-2019Documento12 páginasSTUDOCU Insuficiencia-Cardiaca-Medicina-Interna-De-Harrison-Edicion-2019Camila Cazuriaga100% (1)
- EVCDocumento36 páginasEVCCarolina Diaz Quiroz100% (1)
- HematopoyesisDocumento19 páginasHematopoyesisJocelyne Calderon PadillaAún no hay calificaciones
- Hematopoyesis MonseDocumento5 páginasHematopoyesis Monsejuan Cordova LaraAún no hay calificaciones
- Hematopoyesis ResumenDocumento4 páginasHematopoyesis ResumenErika Elizabeth Medina ToledoAún no hay calificaciones
- HEMATOPOYESISDocumento7 páginasHEMATOPOYESISAileen PresuelAún no hay calificaciones
- Celulas MadreDocumento9 páginasCelulas Madrearmanditocote94Aún no hay calificaciones
- Articulo 2 HematologiaDocumento10 páginasArticulo 2 HematologiaRene Gtz SAún no hay calificaciones
- Hematopoyesis AAyFADocumento15 páginasHematopoyesis AAyFAmarytayupegAún no hay calificaciones
- Terapia Biológica Enfermedades Reumáticas y Anti TNF TrabajoDocumento8 páginasTerapia Biológica Enfermedades Reumáticas y Anti TNF TrabajoBryan Jeremy Guzman SuxoAún no hay calificaciones
- 08 - Prem - Ciencias Naturales 0Documento110 páginas08 - Prem - Ciencias Naturales 0Briceño HbvAún no hay calificaciones
- Linfocitos T y B - 1Documento3 páginasLinfocitos T y B - 1Lourdes Tuanama JaraAún no hay calificaciones
- Inmuno Degras 1 - FinalDocumento117 páginasInmuno Degras 1 - FinaljeanpierreAún no hay calificaciones
- Actividad Ciencias Naturales 7moDocumento1 páginaActividad Ciencias Naturales 7moMia AztridAún no hay calificaciones
- 5 Antígeno e Inmunogeno 2021Documento18 páginas5 Antígeno e Inmunogeno 2021Luna GabrielaAún no hay calificaciones
- Inmunogenetica PANAMA 2020Documento48 páginasInmunogenetica PANAMA 2020Valentin MoranAún no hay calificaciones
- Cuestiones Evau Sol InmunoDocumento5 páginasCuestiones Evau Sol InmunoSilviaAún no hay calificaciones
- Inmunidad AdaptativaDocumento17 páginasInmunidad AdaptativaBianca Martina CastroAún no hay calificaciones
- Artículo de Mecanismos Efectores de La Inmunidad HumoralDocumento7 páginasArtículo de Mecanismos Efectores de La Inmunidad HumoralBryan SaltosAún no hay calificaciones
- Biologia BACHILLERATO SOLUCIONARIO SERIE OBSERVA-88-92Documento5 páginasBiologia BACHILLERATO SOLUCIONARIO SERIE OBSERVA-88-92Juber Orlando Sifuentes julcaAún no hay calificaciones
- Trivia - Sistema LinfaticoDocumento6 páginasTrivia - Sistema LinfaticoMonique SouzaAún no hay calificaciones
- MonografiaDocumento32 páginasMonografianick marinAún no hay calificaciones
- Unidad 3. Mecanismos de Defensa Del Organismo - PDF (10017)Documento70 páginasUnidad 3. Mecanismos de Defensa Del Organismo - PDF (10017)silviaguevara2004Aún no hay calificaciones
- UNID Maduraci+ N Linfocitos B PDFDocumento25 páginasUNID Maduraci+ N Linfocitos B PDFFranKlin Toledo Medina100% (1)
- 11 - Memoria InmunologicaDocumento36 páginas11 - Memoria Inmunologica25984076Aún no hay calificaciones
- NeoplasiasDocumento14 páginasNeoplasiasAngel MejiaAún no hay calificaciones
- Preguntas Resueltas. InmunologíaDocumento17 páginasPreguntas Resueltas. Inmunologíathalia100% (1)
- Anticuerpos 2017 PDFDocumento53 páginasAnticuerpos 2017 PDFAnchundia Kevin100% (1)
- Bienvenidos A La Clase #10, Sistema Sanguíneo e Inmunidad. Sistema Inmune Y La SangreDocumento31 páginasBienvenidos A La Clase #10, Sistema Sanguíneo e Inmunidad. Sistema Inmune Y La SangreJesica MachadoAún no hay calificaciones
- InmunologíaDocumento13 páginasInmunologíaJefferson Andres0% (1)
- Tema 1. InmunologíaDocumento46 páginasTema 1. InmunologíaDiana Deza DelgadoAún no hay calificaciones
- 1) Células y ÓrganosDocumento40 páginas1) Células y ÓrganosJulian BarreraAún no hay calificaciones
- Lupus Eritematoso DiseminadoDocumento28 páginasLupus Eritematoso DiseminadoENMANUEL JESUS SILVA MERCADOAún no hay calificaciones
- Resumen Tema 2 Mecanismos de Defensa Frente A Los Hongos PatógenosDocumento30 páginasResumen Tema 2 Mecanismos de Defensa Frente A Los Hongos PatógenosSteph VivasAún no hay calificaciones
- Sistema Inmunologico LinfaticoDocumento14 páginasSistema Inmunologico LinfaticoDani Angel GaAún no hay calificaciones
- Actividad 8 - Linfocitos B - Martinez Fragozo Jose RonaldoDocumento1 páginaActividad 8 - Linfocitos B - Martinez Fragozo Jose RonaldoJOSÉ RONALDO MARTÍNEZ FRAGOZOAún no hay calificaciones
- Cuestionario InmunologiìaDocumento5 páginasCuestionario Inmunologiìaanon_686516841Aún no hay calificaciones
- Fisiopatología Del Lupus Eritematoso Sistémico (2013) PDFDocumento9 páginasFisiopatología Del Lupus Eritematoso Sistémico (2013) PDFdrheayAún no hay calificaciones
- Trabajo de InvestigaciónDocumento6 páginasTrabajo de InvestigaciónLiceth Monserrate Pico ChávezAún no hay calificaciones