También podría gustarte
- Inmunología 5ta ClaseDocumento5 páginasInmunología 5ta ClaseKatherine VasquezAún no hay calificaciones
- Sistema de Complemento (RESUMEN)Documento16 páginasSistema de Complemento (RESUMEN)Edgar D Sanchez OliverosAún no hay calificaciones
- Sistema de Complemento (Resumen)Documento16 páginasSistema de Complemento (Resumen)Edgar D Sanchez OliverosAún no hay calificaciones
- Sistema de Complemento - IfDocumento9 páginasSistema de Complemento - IfManuel Alejandro Lengua CuliAún no hay calificaciones
- Ensayo Del Sistema de ComplementoDocumento2 páginasEnsayo Del Sistema de ComplementoKevin Denis ChoqueAún no hay calificaciones
- .Sistema de Complemento .Documento11 páginas.Sistema de Complemento .Janacua Figueroa DianaAún no hay calificaciones
- 3 SII Inflamacion y ComplementoDocumento4 páginas3 SII Inflamacion y ComplementoGuille OliveraAún no hay calificaciones
- Resumen Capitulo 4Documento8 páginasResumen Capitulo 4wig in a boxAún no hay calificaciones
- Mediadores Quimicos de La Inflamacion ConvertedwwsDocumento14 páginasMediadores Quimicos de La Inflamacion ConvertedwwsYarima MoralesAún no hay calificaciones
- Cascada de ComplementoDocumento3 páginasCascada de ComplementoCamzAún no hay calificaciones
- Ensayo Del Sistema de ComplementoDocumento2 páginasEnsayo Del Sistema de ComplementoKevin Denis ChoqueAún no hay calificaciones
- REACCIONES INMUNOLÓGICAS Sistema de Complemento RESUMENDocumento11 páginasREACCIONES INMUNOLÓGICAS Sistema de Complemento RESUMENIngrid Tatiana LozanoAún no hay calificaciones
- Sistema de ComplementoDocumento9 páginasSistema de ComplementoLauraCarolinaRestrepoAún no hay calificaciones
- Guía Práctica de Complemento Septiembre 2011Documento7 páginasGuía Práctica de Complemento Septiembre 2011InmunoBlog100% (1)
- Cuestionario Del Sistema Del ComplementoDocumento6 páginasCuestionario Del Sistema Del ComplementoKarla BastidaAún no hay calificaciones
- Repaso Tema 4 InmunologiaDocumento8 páginasRepaso Tema 4 InmunologiaorlynmagdalysAún no hay calificaciones
- Tema 4Documento23 páginasTema 4yelenabatista2Aún no hay calificaciones
- Sistema de Complemento /cascada de CoagulaciónDocumento3 páginasSistema de Complemento /cascada de CoagulaciónStephanie Miranda100% (2)
- 5 Sistema de ComplementoDocumento40 páginas5 Sistema de ComplementoLuan NettoAún no hay calificaciones
- Sistema Del ComplementoDocumento14 páginasSistema Del ComplementoDeysi LévanoAún no hay calificaciones
- Inmunología Tema 2: La Respuesta Inmunitaria CelularDocumento10 páginasInmunología Tema 2: La Respuesta Inmunitaria CelularanaAún no hay calificaciones
- Patologia QuimiocinasDocumento4 páginasPatologia QuimiocinasSegundo BAún no hay calificaciones
- Cetina de Anda Vivian Fernanda - Segundo ExamenDocumento12 páginasCetina de Anda Vivian Fernanda - Segundo ExamenViv De AndaAún no hay calificaciones
- Inflamacion, Fagocitosis y ComplementoDocumento49 páginasInflamacion, Fagocitosis y ComplementoNayeli Espinosa0% (1)
- Tema 10 Sistema de ComplementoDocumento53 páginasTema 10 Sistema de ComplementoMaria elena Diego floresAún no hay calificaciones
- Sistema de Complemento ModificadoDocumento14 páginasSistema de Complemento ModificadoYoshiro ReañoAún no hay calificaciones
- Mediadores Químicos de La InflamaciónDocumento14 páginasMediadores Químicos de La InflamaciónmichippolitiAún no hay calificaciones
- EL Sistema DEL ComplementoDocumento5 páginasEL Sistema DEL ComplementoMiranda LeonettiAún no hay calificaciones
- Sistema de ComplementoDocumento19 páginasSistema de ComplementoAlfonso Dávila LuleAún no hay calificaciones
- + Sistema Del ComplementoDocumento6 páginas+ Sistema Del ComplementoAntonella DuarteAún no hay calificaciones
- InmunoDocumento8 páginasInmunoAndres ErasoAún no hay calificaciones
- iNMUNOLOGÍA TAREAADocumento5 páginasiNMUNOLOGÍA TAREAADiana TrejoAún no hay calificaciones
- SistemadelComplemento CREDocumento5 páginasSistemadelComplemento CRERec 379Aún no hay calificaciones
- Complemento y AutoinmunidadDocumento4 páginasComplemento y Autoinmunidadwilder_meléndez_1Aún no hay calificaciones
- Sistema ComplementoDocumento8 páginasSistema ComplementoAlberto TorrecillasAún no hay calificaciones
- Resumen Via AlternaDocumento8 páginasResumen Via AlternaPRISGPEAún no hay calificaciones
- Sistema Del ComplementoDocumento35 páginasSistema Del ComplementoRomina ArgumedoAún no hay calificaciones
- Inmunología - T7.Abril - GómezDocumento4 páginasInmunología - T7.Abril - Gómezlgomez04Aún no hay calificaciones
- Sistema Del ComplementoDocumento12 páginasSistema Del ComplementoSewa FloresAún no hay calificaciones
- El Sistema Del ComplementoDocumento5 páginasEl Sistema Del Complementovictor quispe CastroAún no hay calificaciones
- Unidad Ii Reaccion AntigenoDocumento19 páginasUnidad Ii Reaccion AntigenoJavier SevillaAún no hay calificaciones
- Chopita - Inmunidad InnataDocumento9 páginasChopita - Inmunidad InnataMartita JuárezAún no hay calificaciones
- Documento Sin TítuloDocumento3 páginasDocumento Sin TítuloSarah DdelgadoAún no hay calificaciones
- El Sistema Del ComplementoDocumento5 páginasEl Sistema Del ComplementoPAOLA ARCILAAún no hay calificaciones
- Factores Quimiotacticos EndogenosDocumento9 páginasFactores Quimiotacticos EndogenosDIEGO ALEJANDRO GARCIA RINCONAún no hay calificaciones
- FAGOCITOSISDocumento55 páginasFAGOCITOSISGabbo BPAún no hay calificaciones
- Clase 002b INMUNOLOGÍA Sistema de Complemento 2022-1Documento12 páginasClase 002b INMUNOLOGÍA Sistema de Complemento 2022-1RUTH GINNA GUERRERO ALTAMIRANOAún no hay calificaciones
- 6-Sistema de ComplementoDocumento9 páginas6-Sistema de Complementogeobannys cedeñoAún no hay calificaciones
- Sistema de ComplementoDocumento17 páginasSistema de ComplementoCarlosErazoAún no hay calificaciones
- Inflamacion-LAS BASES MOLECULARES DE LA INFLAMACIÓNDocumento22 páginasInflamacion-LAS BASES MOLECULARES DE LA INFLAMACIÓNDeidara07Aún no hay calificaciones
- Resistencia Del Organismo A La Infeccion IIDocumento8 páginasResistencia Del Organismo A La Infeccion IIMayra SchafheutleAún no hay calificaciones
- Inmunologia: Funciones Mediadas Por El Sistema de Complemento y NomenclaturaDocumento11 páginasInmunologia: Funciones Mediadas Por El Sistema de Complemento y NomenclaturaAna Jose Viveros HigueraAún no hay calificaciones
- OpsoninasDocumento7 páginasOpsoninasGiancarlo Baldeón CastroAún no hay calificaciones
- Teoria 6 InmunoDocumento16 páginasTeoria 6 Inmunoandrea lyaAún no hay calificaciones
- Inmunidad y AntibioticosDocumento13 páginasInmunidad y AntibioticosKrystel Volgger GarciaAún no hay calificaciones
- Sistema Del ComplementoDocumento6 páginasSistema Del ComplementoDiego JimenezAún no hay calificaciones
- Sistema de ComplementoDocumento11 páginasSistema de ComplementodavidAún no hay calificaciones
- Correccion Exposicion Sistema Nervioso TiburonesDocumento23 páginasCorreccion Exposicion Sistema Nervioso TiburonesSantiago VasquezAún no hay calificaciones
- Ncoronelf@unjbg - Edu.pe: Prof.: Nelver Coronel FloresDocumento49 páginasNcoronelf@unjbg - Edu.pe: Prof.: Nelver Coronel FloresDagne mirella Yarleque fernandezAún no hay calificaciones
- Silabo 2021-1 Bioquimica de Organismos AcuaticosDocumento5 páginasSilabo 2021-1 Bioquimica de Organismos AcuaticosMartin PonceAún no hay calificaciones
- Enzimas - ¿Qué Son y Cómo FuncionanDocumento1 páginaEnzimas - ¿Qué Son y Cómo FuncionanRenata Quintero SuárezAún no hay calificaciones
- Apuntes de Esporulación - ZeniaPuigZDocumento2 páginasApuntes de Esporulación - ZeniaPuigZPuig Zamarroni ZeniaAún no hay calificaciones
- Hoja de Trabajo para El EstudianteDocumento10 páginasHoja de Trabajo para El EstudianteLesly OrtizAún no hay calificaciones
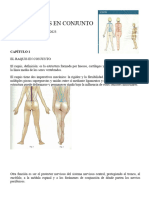
- Raquios en ConjuntoDocumento17 páginasRaquios en ConjuntocarlosAún no hay calificaciones
- Nom 087 Ecol 2002Documento21 páginasNom 087 Ecol 2002JAIR ADRIAN FLORES AGUAYOAún no hay calificaciones
- Examen Diagnóstico 4to GradoDocumento8 páginasExamen Diagnóstico 4to GradoYenyy Elisabeeth ViillamixarAún no hay calificaciones
- Apuntes Biologia Del DesarrolloDocumento6 páginasApuntes Biologia Del DesarrolloanonimousAún no hay calificaciones
- Ficha Evaluación Diagnóstica 30marzo 4º CytDocumento4 páginasFicha Evaluación Diagnóstica 30marzo 4º CytBerita TapullimaAún no hay calificaciones
- Induccion A La Herencia Cuantitativa y Genetica de PoblacionesDocumento8 páginasInduccion A La Herencia Cuantitativa y Genetica de PoblacionesJuan RiosAún no hay calificaciones
- Sistema RespiratorioDocumento2 páginasSistema RespiratorioEmilia Isabel Palma CernaAún no hay calificaciones
- Clasificación de LaboratoriosDocumento22 páginasClasificación de LaboratoriosGabriel HerreraAún no hay calificaciones
- Diversidad y EvoluciónDocumento12 páginasDiversidad y EvoluciónRossy FuentesAún no hay calificaciones
- Nivel de Daño Económico en PlagasDocumento9 páginasNivel de Daño Económico en PlagasMilagro Maria Granados AmayaAún no hay calificaciones
- INTERACCIONES EN LA VIDA DE LA MADRE TIERRA Erwin y LizhetDocumento6 páginasINTERACCIONES EN LA VIDA DE LA MADRE TIERRA Erwin y Lizhetherlanalanoca096Aún no hay calificaciones
- RV 1Documento4 páginasRV 1Mario HiyaneAún no hay calificaciones
- MembranaDocumento15 páginasMembranaANA SIBAJAAún no hay calificaciones
- Animal Mimics SpanishDocumento4 páginasAnimal Mimics SpanishElena FloresAún no hay calificaciones
- Genetica 2023-1 PDFDocumento77 páginasGenetica 2023-1 PDFBIOLOGIA UNIPAMPLONAAún no hay calificaciones
- TRAQUEADocumento23 páginasTRAQUEAJhamile Herrera BlasAún no hay calificaciones
- Tema 2Documento3 páginasTema 2pwhr67vsy9Aún no hay calificaciones
- Nivelación NaturalesDocumento13 páginasNivelación NaturalesDaniel GonzalezAún no hay calificaciones
- Tecnica de Extraccion de AdnDocumento6 páginasTecnica de Extraccion de AdnAmani Nohemi Nieto MuñizAún no hay calificaciones
- Ensayo Importancia de Los InsectosDocumento3 páginasEnsayo Importancia de Los InsectosGalo Alberto Robles MontañoAún no hay calificaciones
- Funciones Del Tejido EpitelialDocumento6 páginasFunciones Del Tejido EpitelialAndy ZúñigaAún no hay calificaciones
- Cadena AlimenticiaDocumento2 páginasCadena AlimenticiaMilagros Ochoa VargasAún no hay calificaciones
- Inteligencia Emocional para Padres IDocumento16 páginasInteligencia Emocional para Padres INune EdurneAún no hay calificaciones
- Presentación Antropologia UASD2023 3Documento18 páginasPresentación Antropologia UASD2023 3María beronica Beltre tejadaAún no hay calificaciones
- Altas Dosis: Los efectos milagrosos de dosis extremadamente altas de vitamina D3. El gran secreto que la industria farmacéutica quiere ocultarle.De EverandAltas Dosis: Los efectos milagrosos de dosis extremadamente altas de vitamina D3. El gran secreto que la industria farmacéutica quiere ocultarle.Calificación: 5 de 5 estrellas5/5 (4)
- Muchas Vidas, Muchos Sabios (Many Lives, Many Masters): (Many Lives, Many Masters)De EverandMuchas Vidas, Muchos Sabios (Many Lives, Many Masters): (Many Lives, Many Masters)Calificación: 4 de 5 estrellas4/5 (475)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Terapia cognitiva: Conceptos básicos y profundizaciónDe EverandTerapia cognitiva: Conceptos básicos y profundizaciónCalificación: 5 de 5 estrellas5/5 (1)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Sana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónDe EverandSana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónCalificación: 5 de 5 estrellas5/5 (4)
- Cardiología y enfermedades cardiovascularesDe EverandCardiología y enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- Shiatsu: Conozca las técnicas y tratamientos de una de las terapias más antiguasDe EverandShiatsu: Conozca las técnicas y tratamientos de una de las terapias más antiguasAún no hay calificaciones
- Neuropsicología: Los fundamentos de la materiaDe EverandNeuropsicología: Los fundamentos de la materiaCalificación: 5 de 5 estrellas5/5 (1)
- Psiconeuroinmunología para la práctica clínicaDe EverandPsiconeuroinmunología para la práctica clínicaCalificación: 5 de 5 estrellas5/5 (4)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)
- Abrázame fuerte: Siete conversaciones para un amor duraderoDe EverandAbrázame fuerte: Siete conversaciones para un amor duraderoManu BerásteguiCalificación: 4.5 de 5 estrellas4.5/5 (13)
- TDAH en Adultos. Cómo Reconocer y Tratar a un Adulto con TDAH en 30 Fáciles PasosDe EverandTDAH en Adultos. Cómo Reconocer y Tratar a un Adulto con TDAH en 30 Fáciles PasosCalificación: 4 de 5 estrellas4/5 (8)
- Fisiopatología de las enfermedades cardiovascularesDe EverandFisiopatología de las enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- Psicópatas seriales: Un recorrido por su oscura e inquietante naturalezaDe EverandPsicópatas seriales: Un recorrido por su oscura e inquietante naturalezaCalificación: 4 de 5 estrellas4/5 (3)
- Nuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosDe EverandNuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosCalificación: 3.5 de 5 estrellas3.5/5 (6)
- Dieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasDe EverandDieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasCalificación: 4 de 5 estrellas4/5 (9)
- Psicoterapia breve con niños y adolescentesDe EverandPsicoterapia breve con niños y adolescentesCalificación: 4.5 de 5 estrellas4.5/5 (15)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- ¿Cómo Prevenir O Curar El Cáncer?: ¡Conozca Los 3 Métodos Probados Contra El Cáncer!De Everand¿Cómo Prevenir O Curar El Cáncer?: ¡Conozca Los 3 Métodos Probados Contra El Cáncer!Aún no hay calificaciones