También podría gustarte
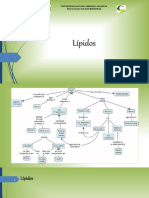
- Los LipidosDocumento40 páginasLos LipidosNelson PerezAún no hay calificaciones
- Síntesis de productos químicos derivados de la biomasa empleando catálisis heterogénea: heterociclos de la biomasaDe EverandSíntesis de productos químicos derivados de la biomasa empleando catálisis heterogénea: heterociclos de la biomasaAún no hay calificaciones
- LÍPIDOSDocumento15 páginasLÍPIDOSJOSE ATENCIAAún no hay calificaciones
- Fisiología renal y metabolismo hidrosalino: Segunda ediciónDe EverandFisiología renal y metabolismo hidrosalino: Segunda ediciónAún no hay calificaciones
- LOS FOSFOLÍPIDOS: ESTRUCTURA, FUNCIONES Y APLICACIONESDocumento7 páginasLOS FOSFOLÍPIDOS: ESTRUCTURA, FUNCIONES Y APLICACIONESOlivia Umanzor DiazAún no hay calificaciones
- Los GlicerolípidosDocumento10 páginasLos GlicerolípidosAnonymous x1GAh2pIbAún no hay calificaciones
- Fosfolipidos y GlucolipidosDocumento8 páginasFosfolipidos y Glucolipidosrafael ramirezAún no hay calificaciones
- Fosfolípidos: componentes esenciales de las membranas celularesDocumento9 páginasFosfolípidos: componentes esenciales de las membranas celularesFrancisco Lancha ChuquilinAún no hay calificaciones
- Lípidos ComplejosDocumento55 páginasLípidos ComplejosAna FilichAún no hay calificaciones
- FOSFOLIPIDOSDocumento3 páginasFOSFOLIPIDOSDavid VillacresAún no hay calificaciones
- FOSFOLIPIDOSDocumento8 páginasFOSFOLIPIDOSSaritaSánchezBustamanteAún no hay calificaciones
- Separata 4 LipidosDocumento12 páginasSeparata 4 LipidosKati RondonAún no hay calificaciones
- Cap52 - Metabolismo de Los Fosfátidos de Glicerina y EsfingolípidosDocumento13 páginasCap52 - Metabolismo de Los Fosfátidos de Glicerina y EsfingolípidosDANIEL ALEJANDRO100% (1)
- TEMA 6 LípidosDocumento14 páginasTEMA 6 LípidosDanielys Perez100% (1)
- LipidosDocumento8 páginasLipidoslauritajuAún no hay calificaciones
- Lipidos y Membranas Celulares Parte IDocumento43 páginasLipidos y Membranas Celulares Parte IMaria AraujoAún no hay calificaciones
- P4 Determinacion de Fosfolipidos, Acidos Grasos Saturados e Insaturados FINALDocumento29 páginasP4 Determinacion de Fosfolipidos, Acidos Grasos Saturados e Insaturados FINALJose AlfredoAún no hay calificaciones
- Practica No. 5. LipidosDocumento9 páginasPractica No. 5. LipidosDaniel Villabona BeltranAún no hay calificaciones
- La CardiolipinaDocumento10 páginasLa Cardiolipinauniversal04-82100% (1)
- ApolipoproteinasDocumento6 páginasApolipoproteinaskatheryn pilcoAún no hay calificaciones
- 4 Fosfolipidos PDFDocumento53 páginas4 Fosfolipidos PDFYoly RiosAún no hay calificaciones
- Los EicosanoidesDocumento14 páginasLos EicosanoidesalriccardiAún no hay calificaciones
- FosfolipidosDocumento6 páginasFosfolipidosJovan Rivero MeralesAún no hay calificaciones
- Deber LipidosDocumento7 páginasDeber LipidosCarlitOox Little MOnzterAún no hay calificaciones
- Biología Humana 6Documento43 páginasBiología Humana 6CELY YELINA CABRERA ZEBALLOSAún no hay calificaciones
- Clasificación LipidosDocumento11 páginasClasificación LipidosAugusto MayoralAún no hay calificaciones
- Lípidos: biomoléculas orgánicas de consistencia sólida o aceitosaDocumento3 páginasLípidos: biomoléculas orgánicas de consistencia sólida o aceitosaJackson López LaviAún no hay calificaciones
- Lipidos ComplejosDocumento36 páginasLipidos ComplejosZaira SolAún no hay calificaciones
- Los Lípidos II. Lípidos ComplejosDocumento83 páginasLos Lípidos II. Lípidos ComplejosSebas Santos Pérez-AnguloAún no hay calificaciones
- LípidosDocumento29 páginasLípidosCarolina VasquezAún no hay calificaciones
- LipidosDocumento10 páginasLipidosMario RangelAún no hay calificaciones
- ABQ05-Bioquímica de Los LípidosDocumento16 páginasABQ05-Bioquímica de Los LípidosUYIAún no hay calificaciones
- Conferencia 4 Lipidos 2024Documento51 páginasConferencia 4 Lipidos 2024jsoyeon568Aún no hay calificaciones
- Ácido FosfatídicoDocumento8 páginasÁcido Fosfatídicosant_grAún no hay calificaciones
- QuimicaDocumento43 páginasQuimicaElvergalargaAún no hay calificaciones
- Ácidos Grasos, Fosfolipidos, Esfingolipidos y EsteroidesDocumento10 páginasÁcidos Grasos, Fosfolipidos, Esfingolipidos y EsteroidesKarina Rivera RincónAún no hay calificaciones
- Entrega Activitats PAU 3Documento5 páginasEntrega Activitats PAU 3DavidAún no hay calificaciones
- Qbio U2 MD06 2022Documento6 páginasQbio U2 MD06 2022Azul CelesteAún no hay calificaciones
- Los Fosfolípidos Son Lípidos Iónicos Polares Compuestos de 1Documento7 páginasLos Fosfolípidos Son Lípidos Iónicos Polares Compuestos de 1Laura Juliana Sacristan YepesAún no hay calificaciones
- Informe de LipoproteínasDocumento16 páginasInforme de LipoproteínasFiorelaMHAún no hay calificaciones
- NUT - Química Metabolismo LipidosDocumento11 páginasNUT - Química Metabolismo LipidosMateo CavallerisAún no hay calificaciones
- Documento PDFDocumento16 páginasDocumento PDFLolitaAún no hay calificaciones
- Lipidos Bq. I 2020Documento61 páginasLipidos Bq. I 2020Niko FavelaAún no hay calificaciones
- Cuadernillo LípidosDocumento10 páginasCuadernillo LípidosAlba PazAún no hay calificaciones
- 7.síntesis de TGDocumento68 páginas7.síntesis de TGDanny Murphy Cahua NuñezAún no hay calificaciones
- 5D Compuestos de Interes BiologicoDocumento20 páginas5D Compuestos de Interes BiologicolujulAún no hay calificaciones
- GuiaDocumento10 páginasGuiaLeidy RiveraAún no hay calificaciones
- Cuestionario de LipidosDocumento6 páginasCuestionario de LipidosLeo RogelAún no hay calificaciones
- LípidosDocumento17 páginasLípidosjose varelaAún no hay calificaciones
- Función y estructura de los principales lípidos de membranaDocumento17 páginasFunción y estructura de los principales lípidos de membranaMonica CazaresAún no hay calificaciones
- Taller - LipidosDocumento8 páginasTaller - LipidosLuis MicoltaAún no hay calificaciones
- Clase 9 - LIPIDOSDocumento46 páginasClase 9 - LIPIDOSpfloresangielisAún no hay calificaciones
- 12 Trabajo Escrito Metabolismo de Acilgliceroles y EsfingolipidosDocumento30 páginas12 Trabajo Escrito Metabolismo de Acilgliceroles y EsfingolipidosTirza Betsabé Lúe RivasAún no hay calificaciones
- Glucolisis Anaerobia BacanDocumento36 páginasGlucolisis Anaerobia BacanBruno Paolo Yalico LeónAún no hay calificaciones
- GlicerofosfolípidosDocumento2 páginasGlicerofosfolípidosDiomedes DiazAún no hay calificaciones
- Guia Farmaco 3Documento11 páginasGuia Farmaco 3MarcTopsAún no hay calificaciones
- Investigación PDFDocumento8 páginasInvestigación PDFBautista Reyes Adela RianelAún no hay calificaciones
- LIPIDOSDocumento43 páginasLIPIDOSFrancy Julieth Triviño BohorquezAún no hay calificaciones
- Taller Polisacáridos y LípidosDocumento10 páginasTaller Polisacáridos y LípidosKaren UrbanoAún no hay calificaciones
- Proyecto 0obtimizacion y Verificacion Del Metodo Analitico para Valoracion Del Piroxicam TabletaDocumento13 páginasProyecto 0obtimizacion y Verificacion Del Metodo Analitico para Valoracion Del Piroxicam TabletaJon LopezAún no hay calificaciones
- Act 1 Libro cuchara menguanteDocumento2 páginasAct 1 Libro cuchara menguanteeducaciontecnologica.isrycAún no hay calificaciones
- OrgZeo FarmaceuticaDocumento74 páginasOrgZeo FarmaceuticaVero HernandezAún no hay calificaciones
- Wuolah Free Tema 5 EnzimologiaDocumento12 páginasWuolah Free Tema 5 EnzimologiaJuan Rodriguez PérezAún no hay calificaciones
- 2 1 Glúcidos - 1Documento35 páginas2 1 Glúcidos - 1LINA GhahremanlouAún no hay calificaciones
- 4 Enlace QuïmicoDocumento6 páginas4 Enlace Quïmico60943807Aún no hay calificaciones
- CÁLCULO DE CONCENTRACIONES DE EQUILIBRIO PARA LA REACCIÓN DE BROMURODocumento28 páginasCÁLCULO DE CONCENTRACIONES DE EQUILIBRIO PARA LA REACCIÓN DE BROMUROLind AguilarAún no hay calificaciones
- TurbinasDocumento4 páginasTurbinasCristy B-flyAún no hay calificaciones
- Super Intensivo Oxigenadas 1Documento4 páginasSuper Intensivo Oxigenadas 1Liliana CondoriAún no hay calificaciones
- Nomenclatura de Oxoácidos 13-10-2020Documento2 páginasNomenclatura de Oxoácidos 13-10-2020Cristian JhonatanAún no hay calificaciones
- Estado Cristalino de Los MetalesDocumento5 páginasEstado Cristalino de Los MetalesShane EnriqueAún no hay calificaciones
- Ficha Técnica Removedor Sapolin (Cumple Componentes Similares)Documento5 páginasFicha Técnica Removedor Sapolin (Cumple Componentes Similares)SSERVICES SASAún no hay calificaciones
- QUIMICADocumento5 páginasQUIMICAmaria lezamaAún no hay calificaciones
- IsómerosDocumento19 páginasIsómerosClemente JoseAún no hay calificaciones
- Mapa Conceptual Sobre La Clasificación, Almacenamiento Temporal y Disposición de Los Residuos de Los Productos QuímicosDocumento2 páginasMapa Conceptual Sobre La Clasificación, Almacenamiento Temporal y Disposición de Los Residuos de Los Productos Químicosdavid gomez100% (6)
- Parcial de NomenclaturaDocumento2 páginasParcial de NomenclaturaJorge TunarozaAún no hay calificaciones
- La Industria Del Calzado y Su Impacto AmbientalDocumento4 páginasLa Industria Del Calzado y Su Impacto AmbientalJhennys GonzalezAún no hay calificaciones
- BiologyDocumento12 páginasBiologymatsssuwuAún no hay calificaciones
- Practica 6Documento3 páginasPractica 6vanngpAún no hay calificaciones
- Protocolo: Revestimientos de Muros "Enlucido Yeso Muros" RE - REV-10 Rev. 1 #CorrelativoDocumento50 páginasProtocolo: Revestimientos de Muros "Enlucido Yeso Muros" RE - REV-10 Rev. 1 #CorrelativojeryAún no hay calificaciones
- Ejercicios de Estequiometria I MedioDocumento10 páginasEjercicios de Estequiometria I MedioGuillermo Paniagua Valdebenito100% (1)
- Practica 10Documento2 páginasPractica 10XimenaAún no hay calificaciones
- Cuestionario de Repaso Acidos NucelicosDocumento4 páginasCuestionario de Repaso Acidos NucelicosYorlen VillaAún no hay calificaciones
- Calentamiento de SustanciasDocumento12 páginasCalentamiento de SustanciasKro BrtlAún no hay calificaciones
- Filtros Membranas Ficha PDocumento3 páginasFiltros Membranas Ficha PSonia CondoriAún no hay calificaciones
- Reactores químicos para tratamiento de aguas residualesDocumento19 páginasReactores químicos para tratamiento de aguas residualesAdri BellamyAún no hay calificaciones
- Trabajo - 01 - P1217B - 2020 - 1Documento8 páginasTrabajo - 01 - P1217B - 2020 - 1Isaac Farfan CondorAún no hay calificaciones
- Norma NFPA 10 Español.Documento125 páginasNorma NFPA 10 Español.Rodrigo86F100% (4)
- Informe Guia Transporte ActivoDocumento5 páginasInforme Guia Transporte ActivoFerney GomezAún no hay calificaciones
- Datos Obtenidos, Curva de Calibración CROMODocumento1 páginaDatos Obtenidos, Curva de Calibración CROMOFernando Olivares LealAún no hay calificaciones
- Minerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.De EverandMinerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.Calificación: 3 de 5 estrellas3/5 (2)
- Lentes de contacto: composición química y propiedadesDe EverandLentes de contacto: composición química y propiedadesCalificación: 3 de 5 estrellas3/5 (2)
- Manual técnico de refrigerantesDe EverandManual técnico de refrigerantesCalificación: 4 de 5 estrellas4/5 (4)
- Fisicoquímica II: Materia, electricidad y magnetismo. Fuerzas y camposDe EverandFisicoquímica II: Materia, electricidad y magnetismo. Fuerzas y camposAún no hay calificaciones
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- Fundamentos de procesos químicosDe EverandFundamentos de procesos químicosCalificación: 5 de 5 estrellas5/5 (3)
- Investigación de mecanismos de reacción en química orgánicaDe EverandInvestigación de mecanismos de reacción en química orgánicaAún no hay calificaciones
- Principios básicos de bioquímica de los alimentosDe EverandPrincipios básicos de bioquímica de los alimentosCalificación: 4.5 de 5 estrellas4.5/5 (2)
- La química al alcance de todosDe EverandLa química al alcance de todosCalificación: 1 de 5 estrellas1/5 (1)
- Fundamentos de química aplicados a las ciencias de la saludDe EverandFundamentos de química aplicados a las ciencias de la saludAún no hay calificaciones
- Nanotecnología: Fundamentos y aplicacionesDe EverandNanotecnología: Fundamentos y aplicacionesAún no hay calificaciones
- Técnicas y medidas básicas en el laboratorio de químicaDe EverandTécnicas y medidas básicas en el laboratorio de químicaCalificación: 4.5 de 5 estrellas4.5/5 (6)
- Bioquímica: metabolismo energético, conceptos y aplicaciónDe EverandBioquímica: metabolismo energético, conceptos y aplicaciónCalificación: 4 de 5 estrellas4/5 (7)
- Lecciones de Química HeterocíclicaDe EverandLecciones de Química HeterocíclicaCalificación: 5 de 5 estrellas5/5 (1)
- Estructura atómica y enlace químicoDe EverandEstructura atómica y enlace químicoCalificación: 3 de 5 estrellas3/5 (2)
- Química orgánica: ejercicios de aplicaciónDe EverandQuímica orgánica: ejercicios de aplicaciónCalificación: 5 de 5 estrellas5/5 (3)
- Nomenclatura de las sustancias químicasDe EverandNomenclatura de las sustancias químicasCalificación: 5 de 5 estrellas5/5 (2)
- Guías de laboratorio de bioquímica para la carrera de químicaDe EverandGuías de laboratorio de bioquímica para la carrera de químicaCalificación: 5 de 5 estrellas5/5 (1)
- UF1667 - Tratamiento de agua potableDe EverandUF1667 - Tratamiento de agua potableCalificación: 4.5 de 5 estrellas4.5/5 (2)