También podría gustarte
- Metabolismo de Los Carbohidratos Diapositivas Bioquímica IDocumento46 páginasMetabolismo de Los Carbohidratos Diapositivas Bioquímica ISteven Vásquez100% (4)
- Almacenamiento y Transporte de Ácidos GrasosDocumento3 páginasAlmacenamiento y Transporte de Ácidos GrasosAlison Dayana100% (1)
- Síntesis y degradación de triglicéridosDocumento7 páginasSíntesis y degradación de triglicéridosdocsamu780% (1)
- Parcial 2 Tema 1 Digestion y Absorcion de LipidosDocumento7 páginasParcial 2 Tema 1 Digestion y Absorcion de Lipidoskaren.pina050703Aún no hay calificaciones
- Síntesis y degradación de triacilglicéridos y cuerpos cetónicosDocumento7 páginasSíntesis y degradación de triacilglicéridos y cuerpos cetónicosAnna VicctóriaAún no hay calificaciones
- Lípidos (Lipogénesis, Lipolisis y Biosíntesis de Ag)Documento51 páginasLípidos (Lipogénesis, Lipolisis y Biosíntesis de Ag)alejandracorpus127Aún no hay calificaciones
- Guia LipoproteinasDocumento8 páginasGuia LipoproteinasSUPER MANUELAún no hay calificaciones
- Semana 01: Lípidos: A. Digestión EN EL EstómagoDocumento17 páginasSemana 01: Lípidos: A. Digestión EN EL EstómagoTracy RamírezAún no hay calificaciones
- Met Lipidos Completo IIDocumento206 páginasMet Lipidos Completo IIDaniAún no hay calificaciones
- Tema 9 - Metabolismo LipídicoDocumento22 páginasTema 9 - Metabolismo LipídicoAntonioAún no hay calificaciones
- Metabolismo de LípidosDocumento25 páginasMetabolismo de LípidosTobias Leonel PerezAún no hay calificaciones
- Digestión y AbsorciónDocumento139 páginasDigestión y AbsorciónDaniAún no hay calificaciones
- Absorción y transporte de lípidosDocumento30 páginasAbsorción y transporte de lípidosAbdielE.AraúzAún no hay calificaciones
- Bio Qui MicaDocumento21 páginasBio Qui Micalefef81847Aún no hay calificaciones
- Manejo de LipidosDocumento11 páginasManejo de LipidosOtto CossioAún no hay calificaciones
- BIOCA3Documento10 páginasBIOCA3Tania AAún no hay calificaciones
- Digestión de LípidosDocumento7 páginasDigestión de LípidosDurbis Astopilco Sosa0% (1)
- Digestion y Absorcion de LipidosDocumento43 páginasDigestion y Absorcion de LipidosMayra HenriquezAún no hay calificaciones
- CLASE 05_PN3_Listo (1)Documento84 páginasCLASE 05_PN3_Listo (1)Marcia NoahiAún no hay calificaciones
- Metabolismo de Los Lípidos M.A.BDocumento5 páginasMetabolismo de Los Lípidos M.A.BomarAún no hay calificaciones
- Lípidos - Organos Afectados y EnfermedadesDocumento16 páginasLípidos - Organos Afectados y EnfermedadesCamila Azucena LazoAún no hay calificaciones
- Digestion y Absorcion de Lipidos - LipoproteinasDocumento38 páginasDigestion y Absorcion de Lipidos - LipoproteinasAlexis Barrientos QuichuaAún no hay calificaciones
- La Glucosa Generalmente Se Considera El Principal Precursor de Carbohidratos para LaDocumento5 páginasLa Glucosa Generalmente Se Considera El Principal Precursor de Carbohidratos para LaALEXANDRA XIMENA MENDEZ AYALAAún no hay calificaciones
- Metabolismo de Li Pidos I 280568 Downloable 3134181Documento15 páginasMetabolismo de Li Pidos I 280568 Downloable 3134181sofiaAún no hay calificaciones
- Metabolismo de Lipidos 2° - 2Documento33 páginasMetabolismo de Lipidos 2° - 2iwannarock86Aún no hay calificaciones
- Capitulo 69 ResumenDocumento2 páginasCapitulo 69 ResumenMelissa JimenezAún no hay calificaciones
- METAB. LÍPIDOS-A AlumnosDocumento8 páginasMETAB. LÍPIDOS-A AlumnosAmarildo CristinoAún no hay calificaciones
- Hoy DisertacionDocumento48 páginasHoy DisertacionMaryen GallardoAún no hay calificaciones
- Guía de Estudio de LípidosDocumento16 páginasGuía de Estudio de LípidosSara Nuñez BravoAún no hay calificaciones
- Metabolismo de Grasas y ProteinasDocumento16 páginasMetabolismo de Grasas y ProteinasBritez Paredes Alejandra SilvanaAún no hay calificaciones
- Unidad 3.metabolismo de LipidosDocumento13 páginasUnidad 3.metabolismo de LipidosEliezer FajardoAún no hay calificaciones
- DISLIPIDEMIADocumento5 páginasDISLIPIDEMIAtrispriorlancasterAún no hay calificaciones
- Digestion de LipidosDocumento17 páginasDigestion de Lipidosqfb04100% (7)
- Hidrolisis EnzimaticaDocumento2 páginasHidrolisis EnzimaticaadrianaAún no hay calificaciones
- Triacilgliceroles: absorción, transporte y lipólisisDocumento23 páginasTriacilgliceroles: absorción, transporte y lipólisisOscar Antonio Cisneros GarciaAún no hay calificaciones
- Accion de La Lipasa PancreaticaDocumento12 páginasAccion de La Lipasa Pancreaticajeyson2150% (2)
- Rol Del Hígado en La Regulación Del MetabolismoDocumento24 páginasRol Del Hígado en La Regulación Del MetabolismoBella Stefanny Morales GuevaraAún no hay calificaciones
- Metabolismo de Los LipidosDocumento23 páginasMetabolismo de Los LipidosAlexandra Crissel Hidalgo PurizagaAún no hay calificaciones
- Metabolismo de LípidosDocumento43 páginasMetabolismo de LípidosBELLA TANIA LLAMO DIAZAún no hay calificaciones
- Catabolismo de LípidosDocumento3 páginasCatabolismo de LípidosAna Garcia MartínezAún no hay calificaciones
- Clase 10 y 11 Metabolismo de LipidosDocumento70 páginasClase 10 y 11 Metabolismo de LipidospfloresangielisAún no hay calificaciones
- Lipidometabolismo: digestión, absorción y transporteDocumento40 páginasLipidometabolismo: digestión, absorción y transportehermes alvarezAún no hay calificaciones
- 001-Digestión y Absorción de LípidosDocumento27 páginas001-Digestión y Absorción de LípidosGreissy Campos Cabrera100% (1)
- Lipogenesis Edit.Documento8 páginasLipogenesis Edit.Gabino MexAún no hay calificaciones
- Bioquimica de Digestion de Chos, Proteinas y LipidosDocumento5 páginasBioquimica de Digestion de Chos, Proteinas y LipidosVALENTINA VASQUEZ ARTEAGAAún no hay calificaciones
- Metabolismo lípidos dietaDocumento4 páginasMetabolismo lípidos dietaMariangel Goyo SandovalAún no hay calificaciones
- 2 Fundamentos de Nutrición Clínica - Interrrelaciones Metabólicas 2020 PDFDocumento28 páginas2 Fundamentos de Nutrición Clínica - Interrrelaciones Metabólicas 2020 PDFcucurruscucucuAún no hay calificaciones
- Capitulo 63-72 GuytonDocumento12 páginasCapitulo 63-72 GuytonGuadalupe Arenas RosasAún no hay calificaciones
- Parcial 2 Tema 6 LipolisisDocumento3 páginasParcial 2 Tema 6 Lipolisiskaren.pina050703Aún no hay calificaciones
- Lipidos II ParteDocumento23 páginasLipidos II ParteLUIS GARCIAAún no hay calificaciones
- 2 Resumen CerdosDocumento3 páginas2 Resumen CerdosNathalySinchiAún no hay calificaciones
- 9-Clase-Lípidos 2022Documento26 páginas9-Clase-Lípidos 2022kennedy Campos HuamaniAún no hay calificaciones
- Repaso BIOQUIMICA IIDocumento3 páginasRepaso BIOQUIMICA IIAlma GabrielaAún no hay calificaciones
- Lipolisis de Ácidos GrasosDocumento8 páginasLipolisis de Ácidos GrasosRicardo VelascoAún no hay calificaciones
- LipólisisDocumento15 páginasLipólisisFer ColinAún no hay calificaciones
- Papel metabólico del hígadoDocumento20 páginasPapel metabólico del hígadoRosamaria Arbañil Arakaki100% (1)
- Metabolismo de Los Lípidos.Documento6 páginasMetabolismo de Los Lípidos.trumandemontesAún no hay calificaciones
- BIOQUIMICA CLINICA - Trabajo FinalDocumento8 páginasBIOQUIMICA CLINICA - Trabajo Finalchrissng21Aún no hay calificaciones
- Metabolismo de LípidosDocumento45 páginasMetabolismo de LípidosLAURA PARRA BUITRAGOAún no hay calificaciones
- Dieta Cetogénica : Dieta Cetogénica Deliciosa Y Fácil Para Una Vida Sana Para Perder PesoDe EverandDieta Cetogénica : Dieta Cetogénica Deliciosa Y Fácil Para Una Vida Sana Para Perder PesoAún no hay calificaciones
- Resumen Fotosintesis y Respiracion CelularDocumento2 páginasResumen Fotosintesis y Respiracion CelularKarlaAún no hay calificaciones
- Taller, BiologíaDocumento18 páginasTaller, BiologíaAdrian CorredorAún no hay calificaciones
- BIOENERGETICADocumento18 páginasBIOENERGETICARossibel100% (1)
- Clase 8 - Metabolismo de Carbohidratos (Ciclo de Krebs y Cadena Transportadora de Electrones)Documento8 páginasClase 8 - Metabolismo de Carbohidratos (Ciclo de Krebs y Cadena Transportadora de Electrones)Danny GutiérrezAún no hay calificaciones
- Taller Oxidación Biológica Final-2020-1Documento1 páginaTaller Oxidación Biológica Final-2020-1Fabio Fajardo67% (3)
- Repaso BCMDocumento4 páginasRepaso BCMEdgar AntonioAún no hay calificaciones
- Licenciatura en Kinesiología y Fisiatría Asignatura: "Kinefilaxia" Material DidacticoDocumento69 páginasLicenciatura en Kinesiología y Fisiatría Asignatura: "Kinefilaxia" Material Didacticosofia violaAún no hay calificaciones
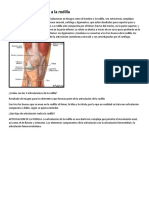
- Elementos Que Forman A La RodillaDocumento3 páginasElementos Que Forman A La RodillaCarlos EliasAún no hay calificaciones
- BiofisicaDocumento25 páginasBiofisicaayelenAún no hay calificaciones
- Fisiología celular y transporte a través de membranasDocumento189 páginasFisiología celular y transporte a través de membranasClara Cali SublimadosAún no hay calificaciones
- Evaluación 3 Periodo Química 10Documento2 páginasEvaluación 3 Periodo Química 10patricia pizarro100% (1)
- Protocolo de ArranqueDocumento4 páginasProtocolo de ArranqueDaniel TorresAún no hay calificaciones
- Informe 06 de Bioquimica AGroindustrialDocumento10 páginasInforme 06 de Bioquimica AGroindustrialAlexito Sotelo0% (1)
- Procesos Bioquímicos PDFDocumento130 páginasProcesos Bioquímicos PDFEstefany Cubillos CortesAún no hay calificaciones
- Cuestionario de Procesos Metabolicos RevisadoDocumento5 páginasCuestionario de Procesos Metabolicos RevisadodanielleAún no hay calificaciones
- Sistemas BioenergéticosDocumento5 páginasSistemas BioenergéticosDiego HerreraAún no hay calificaciones
- MetabolismoDocumento11 páginasMetabolismoJiménez Hernández Heidi MichelleAún no hay calificaciones
- Bases Moleculares y Celulares de La Herencia PDFDocumento136 páginasBases Moleculares y Celulares de La Herencia PDFmacehuatl100% (1)
- Curso Bioquimica de Alimentos PDFDocumento113 páginasCurso Bioquimica de Alimentos PDFEmarthur Castillo100% (1)
- Glucolisis VoetDocumento21 páginasGlucolisis VoetANGIE PAOLA RODELO PANZAAún no hay calificaciones
- Viaje a la célulaDocumento16 páginasViaje a la célulaMario Kike Zarzoza0% (1)
- Bioquímica I test tema agua disolvente electrólitoDocumento10 páginasBioquímica I test tema agua disolvente electrólitoPaula NúñezAún no hay calificaciones
- Practica 8 METABOLISMODocumento12 páginasPractica 8 METABOLISMOSimon Jesus Gonzalez ArangoAún no hay calificaciones
- Metabolismo de Animales Monogástricos y RumiantesDocumento14 páginasMetabolismo de Animales Monogástricos y RumiantesDaniel de Jesús Lopez GutiérrezAún no hay calificaciones
- Lanzadera Glicerol 3 FosfatoDocumento11 páginasLanzadera Glicerol 3 FosfatoAleja Gamba ClavijoAún no hay calificaciones
- Tema 3 BioenergeticaDocumento58 páginasTema 3 BioenergeticaAxel IgleciasAún no hay calificaciones
- Biosintesis de CarbohidratosDocumento13 páginasBiosintesis de CarbohidratosAby GeårAún no hay calificaciones
- Quimica Biologica 3Documento13 páginasQuimica Biologica 3Areliz VillcaAún no hay calificaciones
- El Auge de Las Enfermedades MitocondrialesDocumento8 páginasEl Auge de Las Enfermedades MitocondrialesNinoska MartinezAún no hay calificaciones
- Fisiologia de La PalmaDocumento49 páginasFisiologia de La PalmaDIEGOGRACIAAún no hay calificaciones