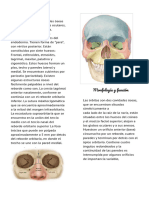
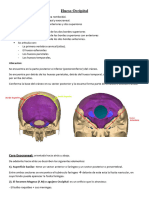
También podría gustarte
- Tema 5 Cavidad OrbitariaDocumento7 páginasTema 5 Cavidad OrbitariaDiana RodriguezAún no hay calificaciones
- Anatomia Del Ojo (Latarjet)Documento7 páginasAnatomia Del Ojo (Latarjet)Jisselle Then Alvarez100% (3)
- Cap 43 y 44 LatarjetDocumento9 páginasCap 43 y 44 LatarjetJaida AbbaAún no hay calificaciones
- Monografia de AnatomiaDocumento38 páginasMonografia de AnatomiaMarco AntonioAún no hay calificaciones
- Anatomía de La Cavidad OrbitariaDocumento11 páginasAnatomía de La Cavidad OrbitariaElizabeth Rondon JimenezAún no hay calificaciones
- Anatomia Órganos de Los SentidosDocumento7 páginasAnatomia Órganos de Los SentidosLucy Orellana RojasAún no hay calificaciones
- Monografia de AnatomiaDocumento38 páginasMonografia de AnatomiaMarco AntonioAún no hay calificaciones
- Tema 4 El OjoDocumento6 páginasTema 4 El OjoArmando J Carrasquel DAún no hay calificaciones
- Ojo AnatomiaDocumento5 páginasOjo AnatomiaNachy SalcedoAún no hay calificaciones
- Organos de La Vision AnatomiaDocumento26 páginasOrganos de La Vision AnatomiaRoci AltamiranoAún no hay calificaciones
- Cavidad OrbitariaDocumento8 páginasCavidad OrbitarialuikarenAún no hay calificaciones
- Anatomia de OjoDocumento13 páginasAnatomia de OjoJaxer Quevedo ThedanieljqAún no hay calificaciones
- Anatomia Esclerotica y CorneaDocumento20 páginasAnatomia Esclerotica y CorneaDr. jorge perez martinez100% (1)
- Esqueleto de La Cavidad Orbitaria MorfoDocumento7 páginasEsqueleto de La Cavidad Orbitaria MorfoYeisy Ruiz SierraAún no hay calificaciones
- Anatomía y fisiología del globo ocularDocumento39 páginasAnatomía y fisiología del globo ocularGianfranco CardonaAún no hay calificaciones
- G L o B o o C U L A R PDFDocumento17 páginasG L o B o o C U L A R PDFjleal22Aún no hay calificaciones
- Huesos de La ÓrbitaDocumento22 páginasHuesos de La ÓrbitaEdgar Antonio DAún no hay calificaciones
- Morfologia Ocular - Dr. EspechDocumento20 páginasMorfologia Ocular - Dr. EspechNicole VitehAún no hay calificaciones
- Orbita y Globo OcularDocumento48 páginasOrbita y Globo OcularGonzalo LunacAún no hay calificaciones
- Apuntes de AnatomiaDocumento8 páginasApuntes de Anatomiaivanna parralesAún no hay calificaciones
- 1.1 Organos de Los Sentidos, 1.2 Ojo y Anexos, 1.3 Vías Neurol Gicas de La Visi NDocumento16 páginas1.1 Organos de Los Sentidos, 1.2 Ojo y Anexos, 1.3 Vías Neurol Gicas de La Visi NAna Karen Moreno PorteAún no hay calificaciones
- Globo OcularDocumento8 páginasGlobo Ocularcelemax40Aún no hay calificaciones
- Anatomia OcularDocumento45 páginasAnatomia OcularCruz Flores Melgarejo100% (1)
- Base Del Craneo o OrificiosDocumento6 páginasBase Del Craneo o OrificiosLuisa Fernanda Triana ValenciaAún no hay calificaciones
- Globo OcularDocumento5 páginasGlobo OcularYOBER KENLLY CABALLERO ROJASAún no hay calificaciones
- Cavidad OrbitariaDocumento14 páginasCavidad OrbitariaDANIELLA ROMINA CHOQUE FLORESAún no hay calificaciones
- Anatomía y función de la órbita ocularDocumento5 páginasAnatomía y función de la órbita ocularDamayra Milla MelchorAún no hay calificaciones
- Ilovepdf MergedDocumento22 páginasIlovepdf MergedASHLY SOFIA MENDEZ ARGUETAAún no hay calificaciones
- Patología de La Órbita TemaDocumento12 páginasPatología de La Órbita Temakenny gomezAún no hay calificaciones
- Anatomia, OcipitalDocumento2 páginasAnatomia, OcipitalElias VillarrealAún no hay calificaciones
- Configuración interna del cráneoDocumento19 páginasConfiguración interna del cráneoAna Lucia Ramirez Molina100% (1)
- Anatomia Del Ojo Y La Orbita OrbitaDocumento10 páginasAnatomia Del Ojo Y La Orbita OrbitaWILMER ERASO ROSEROAún no hay calificaciones
- Exposicion OjoDocumento14 páginasExposicion Ojojohan DiazAún no hay calificaciones
- 2 - Craneo y Cara (Parte 2)Documento30 páginas2 - Craneo y Cara (Parte 2)Juliana CalcumilAún no hay calificaciones
- ÓrbitaDocumento12 páginasÓrbitaAura GallardoAún no hay calificaciones
- MEDACC08 Documento 1.3Documento4 páginasMEDACC08 Documento 1.3Isabel JiménezAún no hay calificaciones
- Cavidad OrbitariaDocumento8 páginasCavidad OrbitariaJhoana velascoAún no hay calificaciones
- Modulo 1 Optometria 1Documento29 páginasModulo 1 Optometria 1Zophy Kérensky Azhkenazi100% (2)
- Cavidad Orbitaria.Documento26 páginasCavidad Orbitaria.orangel fernandezAún no hay calificaciones
- CuestionariosDocumento1 páginaCuestionariosdarkpaintbrosAún no hay calificaciones
- Anatomia Del OjoDocumento8 páginasAnatomia Del OjoYatzari Faridy Aguedo EspecianoAún no hay calificaciones
- NEUROCRANEODocumento10 páginasNEUROCRANEOAredzitha Carrada50% (2)
- OJODocumento3 páginasOJOValentina BolgianiAún no hay calificaciones
- Hueso OccipitalDocumento5 páginasHueso OccipitalLaura Fernanda DíazAún no hay calificaciones
- Regiones Comunes Del Craneo y CaraDocumento18 páginasRegiones Comunes Del Craneo y CaraJheremy SolisAún no hay calificaciones
- Ensayo Investigacion ManuelDocumento11 páginasEnsayo Investigacion Manuelfatima lopezAún no hay calificaciones
- Omoplato Explicacion de PartesDocumento8 páginasOmoplato Explicacion de PartesSebastian BravoAún no hay calificaciones
- Órganos del sistema visual: ojo, vías ópticas y corteza visualDocumento34 páginasÓrganos del sistema visual: ojo, vías ópticas y corteza visualBLANCA LUCILA RODRIGUEZ CARRUITEROAún no hay calificaciones
- Sistema nervioso central: Estructuras óseas y membranas protectorasDocumento126 páginasSistema nervioso central: Estructuras óseas y membranas protectorasPerfil Bladimir SalinasAún no hay calificaciones
- Hueso Esfenoides 1Documento24 páginasHueso Esfenoides 1Jaz MarquezAún no hay calificaciones
- ojoDocumento51 páginasojoyasnaya.cebracAún no hay calificaciones
- Base de CráneoDocumento17 páginasBase de CráneoDaiana Alejandra RomeroAún no hay calificaciones
- Fosas CranealesDocumento15 páginasFosas CranealesManuel ChervoAún no hay calificaciones
- 42 Oftalmología ForenseDocumento28 páginas42 Oftalmología ForenseJulio Adrián Rodríguez ValladaresAún no hay calificaciones
- Relaciones Con Paredes de La Órbita:: Capa Fibrosa (Externa)Documento3 páginasRelaciones Con Paredes de La Órbita:: Capa Fibrosa (Externa)isis ortegaAún no hay calificaciones
- Hueso Maxilar Cirugia AgregadoDocumento13 páginasHueso Maxilar Cirugia AgregadoLyz AnnAún no hay calificaciones
- Tema 1 El CraneoDocumento10 páginasTema 1 El Craneoyoseannysgomez1617Aún no hay calificaciones
- Biofísica de La VisiónDocumento13 páginasBiofísica de La VisiónChristopher EstevezAún no hay calificaciones
- OpioidesDocumento10 páginasOpioidesTitto MéndezAún no hay calificaciones
- Protesis Fija Plural 2016Documento4 páginasProtesis Fija Plural 2016Titto Méndez100% (1)
- Urgencias PerdiodontalesDocumento8 páginasUrgencias PerdiodontalesTitto MéndezAún no hay calificaciones
- Nutrición Infantil 2015Documento14 páginasNutrición Infantil 2015Titto MéndezAún no hay calificaciones
- Evaluacion Imagenológica Cancer OralDocumento8 páginasEvaluacion Imagenológica Cancer OralTitto MéndezAún no hay calificaciones
- Guia Salud Mental BomberosDocumento23 páginasGuia Salud Mental BomberosSamuel E. MarinaoAún no hay calificaciones
- CBCT: Revolucionario escáner dentalDocumento4 páginasCBCT: Revolucionario escáner dentalTitto MéndezAún no hay calificaciones
- Manual Educativo Programa de Salud Bucal - JUNAEBDocumento235 páginasManual Educativo Programa de Salud Bucal - JUNAEBJavier Baquedano Lobos0% (1)
- Tera Pia Periodontal No Qui Rur GicaDocumento10 páginasTera Pia Periodontal No Qui Rur GicaTitto MéndezAún no hay calificaciones
- Etapasde NollaDocumento1 páginaEtapasde NollaTitto MéndezAún no hay calificaciones
- 04 Norma 4Documento5 páginas04 Norma 4Titto MéndezAún no hay calificaciones
- Manual de SemiologiaDocumento206 páginasManual de SemiologiaTitto MéndezAún no hay calificaciones
- Normas en La Atencion OdontologicaDocumento34 páginasNormas en La Atencion OdontologicaTitto MéndezAún no hay calificaciones
- Histologi - A y Anatomi - A PeriodontalDocumento11 páginasHistologi - A y Anatomi - A PeriodontalTitto MéndezAún no hay calificaciones
- Diagnóstico Radiográfico de Caries IIDocumento12 páginasDiagnóstico Radiográfico de Caries IITitto MéndezAún no hay calificaciones
- Ergonomía y Campo QuirurgicoDocumento4 páginasErgonomía y Campo QuirurgicoTitto MéndezAún no hay calificaciones
- Cario IIDocumento10 páginasCario IITitto MéndezAún no hay calificaciones
- EstrésDocumento13 páginasEstrésTitto MéndezAún no hay calificaciones
- 1 y 2 AnaerobiosDocumento10 páginas1 y 2 AnaerobiosTitto MéndezAún no hay calificaciones
- Definición y Clasificación de Los Trastornos Temporomandibulares (TTM)Documento9 páginasDefinición y Clasificación de Los Trastornos Temporomandibulares (TTM)Titto MéndezAún no hay calificaciones
- 04 Norma 4Documento5 páginas04 Norma 4Titto MéndezAún no hay calificaciones
- Hipomovilidad Mandibular CrónicaDocumento2 páginasHipomovilidad Mandibular CrónicaTitto MéndezAún no hay calificaciones
- FonoarticulacionDocumento5 páginasFonoarticulacionTitto MéndezAún no hay calificaciones
- Patologias AmbientalesDocumento12 páginasPatologias AmbientalesTitto MéndezAún no hay calificaciones
- BacteriasDocumento13 páginasBacteriasTitto MéndezAún no hay calificaciones
- Metabolism oDocumento10 páginasMetabolism oTitto MéndezAún no hay calificaciones
- I FisiopatomsDocumento4 páginasI FisiopatomsTitto MéndezAún no hay calificaciones
- Diabetes MellitusDocumento11 páginasDiabetes MellitusTitto MéndezAún no hay calificaciones
- Apuntes Fisiologia Oral 2010Documento47 páginasApuntes Fisiologia Oral 2010Pablo CardenasAún no hay calificaciones
- Apuntes Fisiologia Oral 2010Documento47 páginasApuntes Fisiologia Oral 2010Pablo CardenasAún no hay calificaciones
- Estructura Del Segmento PosteriorDocumento3 páginasEstructura Del Segmento PosteriorMelissa JarabaAún no hay calificaciones
- Anatomia de La OrbitaDocumento21 páginasAnatomia de La OrbitaMila montezumaAún no hay calificaciones
- Diseccion OjoDocumento3 páginasDiseccion OjoJesúsAún no hay calificaciones
- Apuntes de OftalmologíaDocumento184 páginasApuntes de OftalmologíaNicolas FloresAún no hay calificaciones
- UNP-Anatomia y Fisiologia Del Organo de La VisionDocumento33 páginasUNP-Anatomia y Fisiologia Del Organo de La VisionRamirez Morales MiriamAún no hay calificaciones
- Anatomía Del Globo OcularDocumento40 páginasAnatomía Del Globo OcularMarcielo IbarraAún no hay calificaciones
- Embriología ocularDocumento10 páginasEmbriología oculardanna belloAún no hay calificaciones
- Copa Óptica Y Vesícula Del Cristalino: Retina, Iris Y Cuerpo CiliarDocumento3 páginasCopa Óptica Y Vesícula Del Cristalino: Retina, Iris Y Cuerpo CiliarAixa CamposAún no hay calificaciones
- Guía disección ojo vaca biologíaDocumento4 páginasGuía disección ojo vaca biologíaEugaldeAún no hay calificaciones
- Infografía AnatomíaDocumento2 páginasInfografía AnatomíaLiiz Bitiz HerreraAún no hay calificaciones
- Introducción A La OftalmologíaDocumento4 páginasIntroducción A La OftalmologíaFer JuarezAún no hay calificaciones
- Resumen Ojo HISTOLOGIADocumento9 páginasResumen Ojo HISTOLOGIAAntonia ZambranoAún no hay calificaciones
- Sesión 16. Estesiología IIDocumento32 páginasSesión 16. Estesiología IIJosi CostillaAún no hay calificaciones
- Segundo Parcial Funcion Humana C1Documento2 páginasSegundo Parcial Funcion Humana C1Ishiiwatary BimboAún no hay calificaciones
- Tecnicas de IluminacionDocumento2 páginasTecnicas de Iluminacionandrea saavedraAún no hay calificaciones
- Oftalmo Total ResumenDocumento43 páginasOftalmo Total ResumenPamela GonellAún no hay calificaciones
- Disección de Un OjoDocumento18 páginasDisección de Un OjoJulieth Caballero100% (1)
- T9. CristalinoDocumento3 páginasT9. CristalinoKarlii NicholsAún no hay calificaciones
- Trabajo Investigativo-Capa DuaDocumento6 páginasTrabajo Investigativo-Capa DuaDayanna ValenciaAún no hay calificaciones
- Aparato de La VisionDocumento3 páginasAparato de La Visionraul escobarAún no hay calificaciones
- Anatomía y fisiología ocular: estructuras y funciones del ojo humanoDocumento11 páginasAnatomía y fisiología ocular: estructuras y funciones del ojo humanocamilla arandaAún no hay calificaciones
- Iris, Cuerpo Ciliar y CristalinoDocumento50 páginasIris, Cuerpo Ciliar y CristalinoGema Zamora LoorAún no hay calificaciones
- C13 - Desarrollo de Los Ojos y OidosDocumento21 páginasC13 - Desarrollo de Los Ojos y OidosDanny Torres ZAún no hay calificaciones
- 1-Anatomía Del Globo Ocular y Sus Anexos FinalesDocumento45 páginas1-Anatomía Del Globo Ocular y Sus Anexos FinaleswendyluzsanchezAún no hay calificaciones
- Clase N°71. Cirugías de Catarata + LIO PDFDocumento62 páginasClase N°71. Cirugías de Catarata + LIO PDFMILENKA NekoAún no hay calificaciones
- Histología Del OjoDocumento109 páginasHistología Del OjoDulceSanchezAún no hay calificaciones
- Oftalmoscopia Directa Practica2Documento33 páginasOftalmoscopia Directa Practica2Isarú LFAún no hay calificaciones
- Documento de Apoyo Oftalmología 2022Documento6 páginasDocumento de Apoyo Oftalmología 2022Fernanda Gutiérrez EscobarAún no hay calificaciones
- Óptica Fisiológica I - Refracción y fisiología de la visiónDocumento4 páginasÓptica Fisiológica I - Refracción y fisiología de la visiónAlfredo IntriagoAún no hay calificaciones
- Evaluacion Ojo Agosto 5Documento5 páginasEvaluacion Ojo Agosto 5Carlos ChamorroAún no hay calificaciones