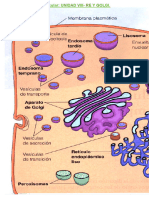
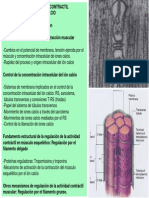
También podría gustarte
- Páginas de Manual de Biología Celular-2Documento16 páginasPáginas de Manual de Biología Celular-2camila.sanchez20066Aún no hay calificaciones
- 20.retículo Endoplasmico RugosoDocumento22 páginas20.retículo Endoplasmico RugosoNadia SanchezAún no hay calificaciones
- Re, Golgi, Lisosoma, Peroxisoma, MitocondriaDocumento118 páginasRe, Golgi, Lisosoma, Peroxisoma, MitocondriaJosselin Saavedra LópezAún no hay calificaciones
- SEMDocumento11 páginasSEMjulissagonzalezzuritaAún no hay calificaciones
- Retículo RugosoDocumento6 páginasRetículo RugosoZuriel HernandezAún no hay calificaciones
- El Retículo EndoplasmáticoDocumento11 páginasEl Retículo EndoplasmáticoMartin CrisantoAún no hay calificaciones
- Citología Parcial 2Documento58 páginasCitología Parcial 2Aroa SilvaAún no hay calificaciones
- Trabajo de BiologiaDocumento11 páginasTrabajo de Biologialoaizaj12Aún no hay calificaciones
- Transporte de ProteinasDocumento9 páginasTransporte de ProteinasGise MathierAún no hay calificaciones
- Complejo RE y Aparato de GolgiDocumento38 páginasComplejo RE y Aparato de GolgileinyAún no hay calificaciones
- Sistema de Membranas Citoplasmaticas o Sistema de EndomembranasDocumento9 páginasSistema de Membranas Citoplasmaticas o Sistema de EndomembranasValentina CelaniAún no hay calificaciones
- Presentacion Retículo EndoplásmicoDocumento46 páginasPresentacion Retículo Endoplásmicobresojiar100% (2)
- Biologia SEMANA 9Documento4 páginasBiologia SEMANA 9john salchichon67% (9)
- Reticulo Endoplasmatico - Distribución y Transporte de ProteínasDocumento5 páginasReticulo Endoplasmatico - Distribución y Transporte de ProteínasSara NogueiraAún no hay calificaciones
- Proteinas Biologia LauDocumento14 páginasProteinas Biologia LauRita TolabaAún no hay calificaciones
- Distribución y Transporte de Proteínas PDFDocumento38 páginasDistribución y Transporte de Proteínas PDFtaurito26100% (1)
- CITOLOGIADocumento3 páginasCITOLOGIARo11Aún no hay calificaciones
- Cap.10 Orgánulos Que Participan en La Sintesis y DegradacionDocumento21 páginasCap.10 Orgánulos Que Participan en La Sintesis y DegradacionJuan Carlos Ledo GarciaAún no hay calificaciones
- Sistema de EndomembranasDocumento27 páginasSistema de EndomembranasSofía TorresAún no hay calificaciones
- MEMBRANA. ExposicionDocumento16 páginasMEMBRANA. ExposicionJuan PabloAún no hay calificaciones
- Sistema de EndomembranasDocumento35 páginasSistema de EndomembranasMaria Paula Quintero TamayoAún no hay calificaciones
- Taller EndomembranasDocumento9 páginasTaller EndomembranasPatty Lopez0% (1)
- Problema Integrador TP3Documento1 páginaProblema Integrador TP3Kevin Andres Z.Aún no hay calificaciones
- Reticulo Endoplasmatico RugosoDocumento9 páginasReticulo Endoplasmatico RugosoJt La Verdad Sin Verdad100% (1)
- Tema 9. Retículo Endoplásmico Liso y RugosoDocumento48 páginasTema 9. Retículo Endoplásmico Liso y RugosoCristina BlancoAún no hay calificaciones
- Retículo Endoplásmico y Secreción de ProteínasDocumento7 páginasRetículo Endoplásmico y Secreción de ProteínasKevin Andres Z.Aún no hay calificaciones
- Modulo 8-04Documento17 páginasModulo 8-04Gustavo RaeleAún no hay calificaciones
- Tema 5 Endomembranas Acortado 2018-2019Documento46 páginasTema 5 Endomembranas Acortado 2018-2019Daniel Fernandez-ValenciaAún no hay calificaciones
- Tema 5Documento9 páginasTema 5Irene Pulido SerranoAún no hay calificaciones
- E11 Retà - Culo EndoplasmáTicoDocumento28 páginasE11 Retà - Culo EndoplasmáTicoIago AraújoAún no hay calificaciones
- Distribución y Transporte de ProteínasDocumento41 páginasDistribución y Transporte de ProteínasJhonny Ponce Agui100% (2)
- Unidad 3 - Sistemas de EndomembranasDocumento17 páginasUnidad 3 - Sistemas de EndomembranasMelAún no hay calificaciones
- Sistema de EndomembranasDocumento50 páginasSistema de EndomembranasAngela AliagaAún no hay calificaciones
- Sistema de EndomenbranasDocumento25 páginasSistema de Endomenbranaselmedico2013Aún no hay calificaciones
- 10.1 - Sist Endome - Reticulo EndoplasmaticoDocumento32 páginas10.1 - Sist Endome - Reticulo Endoplasmaticojanire cabezasAún no hay calificaciones
- Retículo Endoplásmico Pi MZVDocumento29 páginasRetículo Endoplásmico Pi MZVCool BoyLVAún no hay calificaciones
- TEMA 10 Apuntes Bio-CelDocumento12 páginasTEMA 10 Apuntes Bio-CelBelén FernándezAún no hay calificaciones
- Biologia - Sistema de EndomembranasDocumento39 páginasBiologia - Sistema de EndomembranasDRAW KING ArtAún no hay calificaciones
- Unidad 5 BioDocumento7 páginasUnidad 5 Biomolina.ailen99Aún no hay calificaciones
- Taller OrgánulosDocumento23 páginasTaller OrgánulosJose TôrrësAún no hay calificaciones
- Tráfico VesicularDocumento55 páginasTráfico VesicularAficionadoAún no hay calificaciones
- Vesículas de Transporte o de TransiciónDocumento4 páginasVesículas de Transporte o de TransiciónÁlex Willan Acuña SilvaAún no hay calificaciones
- Retículo EndoplasmáticoDocumento20 páginasRetículo EndoplasmáticoAlba Macipe FeitoAún no hay calificaciones
- Tema 6 AcabadoDocumento13 páginasTema 6 AcabadoMagdalena Oñate SerranoAún no hay calificaciones
- Biología Re y GolgiDocumento6 páginasBiología Re y GolgiNabila InsfranAún no hay calificaciones
- Resolucion Guia 4 - Ayudantias DBIO1070Documento11 páginasResolucion Guia 4 - Ayudantias DBIO1070MarianaAún no hay calificaciones
- TEMA 5 - v1Documento19 páginasTEMA 5 - v1Bea LopezAún no hay calificaciones
- Reticulo EndoplasmaticoDocumento9 páginasReticulo EndoplasmaticoEnrique Ochoa0% (1)
- Reticulo Endoplasmatico Aparato de Golgi LisosomasDocumento28 páginasReticulo Endoplasmatico Aparato de Golgi LisosomasDiegoAún no hay calificaciones
- Retículo EndoplasmáticoDocumento8 páginasRetículo Endoplasmáticocongaconga123Aún no hay calificaciones
- Bloque Tematico 2 BiologiaDocumento39 páginasBloque Tematico 2 Biologiaangelica carrilloAún no hay calificaciones
- CLASE 3 Sistema de EndomembranasDocumento30 páginasCLASE 3 Sistema de EndomembranasKeren RuizAún no hay calificaciones
- Retículo Endoplásmico Rugoso.: Fisiología CelularDocumento23 páginasRetículo Endoplásmico Rugoso.: Fisiología CelularYane RosalesAún no hay calificaciones
- 13) Reticulo EndoplasmicoDocumento27 páginas13) Reticulo EndoplasmicoPerla Gutiérrez RogelAún no hay calificaciones
- Clase 11 - Sistema de EndomembranasDocumento29 páginasClase 11 - Sistema de EndomembranasBruno Emanuelli100% (1)
- Reticulo Endoplasmatico Aparato de Golgi Lisosomas-With-Cover-Page-V2 PDFDocumento29 páginasReticulo Endoplasmatico Aparato de Golgi Lisosomas-With-Cover-Page-V2 PDFScarlett OrtizAún no hay calificaciones
- Peroxisoma y REL1Documento7 páginasPeroxisoma y REL1Uther PendragonAún no hay calificaciones
- Sistemas de Endomembranas - Peroxisomas - Vacuola 2021-IIDocumento49 páginasSistemas de Endomembranas - Peroxisomas - Vacuola 2021-IIKaren FelixAún no hay calificaciones
- MITOCONDRIADocumento7 páginasMITOCONDRIAJorge Luis100% (1)
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
- Celula Mitosis MeiosisDocumento11 páginasCelula Mitosis MeiosisMonikaDiosa04Aún no hay calificaciones
- ALFALFADocumento6 páginasALFALFACarmen IuzAún no hay calificaciones
- 4 M Todos Proteina Lic. Q. Marcela TorresDocumento32 páginas4 M Todos Proteina Lic. Q. Marcela TorresisabellaAún no hay calificaciones
- 08 PAU MetabolismoDocumento24 páginas08 PAU MetabolismoevaAún no hay calificaciones
- BMC8 9 12Documento10 páginasBMC8 9 12Juan Carlos LescanoAún no hay calificaciones
- Autoevaluaciones - 2 Bioquimica Metabolica Trabajo FinalDocumento15 páginasAutoevaluaciones - 2 Bioquimica Metabolica Trabajo FinalDyan Di TorreAún no hay calificaciones
- Fases de La FarmacocinéticaDocumento1 páginaFases de La FarmacocinéticaDiana JoselinAún no hay calificaciones
- Clase 01 Biologia Cpech - Organizacion Biologica v.2 (OliverClases)Documento39 páginasClase 01 Biologia Cpech - Organizacion Biologica v.2 (OliverClases)Oliver Henriquez Aracena100% (7)
- Hormonas VegetalesDocumento12 páginasHormonas VegetalesVALENTINA SANCHEZ GAVIRIAAún no hay calificaciones
- La Boca, La Faringe.Documento18 páginasLa Boca, La Faringe.Florinda RojasAún no hay calificaciones
- Ciclo Celular y Procesos de CrecimientoDocumento3 páginasCiclo Celular y Procesos de CrecimientoPacoLAún no hay calificaciones
- Tema IV Bacteriología MédicaDocumento37 páginasTema IV Bacteriología Médicavania anayaAún no hay calificaciones
- Tema 5. Digestión, Absorción y MetabolismoDocumento36 páginasTema 5. Digestión, Absorción y MetabolismoMarcos Belmar OrtizAún no hay calificaciones
- Movimiento Microbiano y Respuestas SensorialesDocumento12 páginasMovimiento Microbiano y Respuestas SensorialesIsabella MartinezAún no hay calificaciones
- Resistencia Bacteriana1Documento58 páginasResistencia Bacteriana1Elisa ArteagaAún no hay calificaciones
- Respuestas Guia de Biologia 2º Medio Manipulacion GeneticaDocumento3 páginasRespuestas Guia de Biologia 2º Medio Manipulacion GeneticaJaviera ValenzuelaAún no hay calificaciones
- Material GeneticoDocumento9 páginasMaterial GeneticoBrayan Gregori Uceda PaicoAún no hay calificaciones
- Reticulo Endoplasmatico Liso .....Documento7 páginasReticulo Endoplasmatico Liso .....DJ CRONOS100% (4)
- Hormonas (Mecanismo de Accion)Documento1 páginaHormonas (Mecanismo de Accion)Laisha Lynn Teran SantiagoAún no hay calificaciones
- MONOGASTRICOSDocumento21 páginasMONOGASTRICOSLupita SeFiAún no hay calificaciones
- Colegio Comunal Albert Einstein: "Educamos para Forjar Personas en Valores, Convivencia y Comportamiento Social"Documento2 páginasColegio Comunal Albert Einstein: "Educamos para Forjar Personas en Valores, Convivencia y Comportamiento Social"kiromy ytAún no hay calificaciones
- Cuestionario Principios Basicos de FarmacologiaDocumento4 páginasCuestionario Principios Basicos de FarmacologiaJüän Vërdthy FüëńtësAún no hay calificaciones
- Interacciones Farmacológicas de Los Anticuerpos MonoclonalesDocumento8 páginasInteracciones Farmacológicas de Los Anticuerpos MonoclonalesjosueAún no hay calificaciones
- Nuevo Documento de Microsoft WordDocumento3 páginasNuevo Documento de Microsoft WordLaura CalderonAún no hay calificaciones
- Bioquimica Carbohidratos y LipidosDocumento16 páginasBioquimica Carbohidratos y LipidosRodrigo Iriarte Gomez100% (1)
- 4enzimas Restriccion 1Documento44 páginas4enzimas Restriccion 1Ickroz ZuñigaAún no hay calificaciones
- El Nucléolo enDocumento29 páginasEl Nucléolo enFredy Alberto CabasAún no hay calificaciones
- Practica 7 Delgado y DiazDocumento10 páginasPractica 7 Delgado y DiazEdith Citlalli Diaz EstradaAún no hay calificaciones
- Modelos Preguntas Teoricas RespuestasDocumento29 páginasModelos Preguntas Teoricas RespuestasKatyalejandra Verjel NavaroAún no hay calificaciones
- Informe Biologia Acción de Una Enzima de Tejidos Vegetales y AnimalesDocumento12 páginasInforme Biologia Acción de Una Enzima de Tejidos Vegetales y AnimalesJuan Camilo Crespo Duque100% (1)