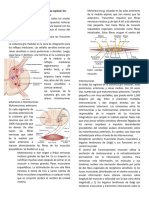
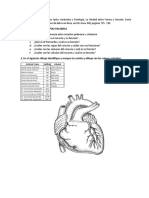
También podría gustarte
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- Metodo RoodDocumento27 páginasMetodo RoodJulio Franco100% (1)
- CV LARENAS, IVAN EspañolDocumento3 páginasCV LARENAS, IVAN EspañolDavid PhillipsAún no hay calificaciones
- Cardiología PDFDocumento51 páginasCardiología PDFPablo Novoa FernándezAún no hay calificaciones
- Trabajo Inicial TAM 19 de Abril (Finalizado 2.0)Documento20 páginasTrabajo Inicial TAM 19 de Abril (Finalizado 2.0)David Phillips0% (2)
- PiCCO Plus Monitorización Cardiopulmonar Mínimamente PDFDocumento11 páginasPiCCO Plus Monitorización Cardiopulmonar Mínimamente PDFmiguel gonzalezAún no hay calificaciones
- Fisiopatología CardiovascularDocumento17 páginasFisiopatología CardiovascularAlex Comun GallegosAún no hay calificaciones
- Resumen NeurobiologiaDocumento20 páginasResumen NeurobiologiaAdrianaAún no hay calificaciones
- La Respiración en Las Artes Marciales PDFDocumento8 páginasLa Respiración en Las Artes Marciales PDFoscar rinconAún no hay calificaciones
- Arco ReflejoDocumento9 páginasArco ReflejoAlba ArGlezAún no hay calificaciones
- Receptores SensorialesDocumento3 páginasReceptores SensorialeslancelotAún no hay calificaciones
- ReflejoS HumanosDocumento13 páginasReflejoS HumanosClaudia ParadaAún no hay calificaciones
- Arco Reflejo WordDocumento7 páginasArco Reflejo WordDayanna Mecías SolórzanoAún no hay calificaciones
- Arco ReflejoDocumento6 páginasArco Reflejomilnar2005_232102646Aún no hay calificaciones
- Sistema Nervioso Katherine SuarezDocumento6 páginasSistema Nervioso Katherine SuarezKatherine SuarezAún no hay calificaciones
- 1 Sistema Sensorio MotorDocumento23 páginas1 Sistema Sensorio MotorMarcos CastroAún no hay calificaciones
- Extra 5. Reflejos EspinalesDocumento6 páginasExtra 5. Reflejos EspinalesAlbaAún no hay calificaciones
- MFH Ii - Ao 2 PDFDocumento9 páginasMFH Ii - Ao 2 PDFCarlismar QueralesAún no hay calificaciones
- Practica Nro 3 Sistema Nervioso (24970)Documento4 páginasPractica Nro 3 Sistema Nervioso (24970)Gabriela GutierrezAún no hay calificaciones
- Arco Reflejo, Acto Reflejo. DoctoraDocumento8 páginasArco Reflejo, Acto Reflejo. DoctoraDayana Franco SilvaAún no hay calificaciones
- 5-Sistema SomatosensorialDocumento13 páginas5-Sistema SomatosensorialJavier FernandezAún no hay calificaciones
- Morfofisiologia Humana 2 para EstudiarDocumento17 páginasMorfofisiologia Humana 2 para Estudiarlesviau44Aún no hay calificaciones
- Arco Reflejo - 26.08Documento14 páginasArco Reflejo - 26.08VALENTINA FANDIãO MIRANDAAún no hay calificaciones
- Clase 20Documento11 páginasClase 20Nere ProañoAún no hay calificaciones
- Clase 4 Organos de Los Sentidos-1Documento32 páginasClase 4 Organos de Los Sentidos-1wilson clavijoAún no hay calificaciones
- SEPARATA Sensibilidad Somática General y Pares CranealesDocumento8 páginasSEPARATA Sensibilidad Somática General y Pares CranealesJosue Dc100% (1)
- Comunicacion Neuroral Tema2Documento6 páginasComunicacion Neuroral Tema2Miguel Antonio NMMAún no hay calificaciones
- Reflejo MedularDocumento6 páginasReflejo MedularNatalie MitreAún no hay calificaciones
- Tejido NerviosoDocumento63 páginasTejido NerviosoyorgeAún no hay calificaciones
- Reflejo LaboratorioDocumento2 páginasReflejo Laboratorioexcadete60% (5)
- 43-44 MC - Reflejos MedularesDocumento15 páginas43-44 MC - Reflejos MedularesTersAún no hay calificaciones
- Fisiologia Laboratorio 2Documento4 páginasFisiologia Laboratorio 2Grecia GarayAún no hay calificaciones
- 04 Concepto de ReflejoDocumento3 páginas04 Concepto de ReflejoMayka RamosAún no hay calificaciones
- Sistema SensorialDocumento22 páginasSistema SensorialPamela SaldiasAún no hay calificaciones
- ReceptorDocumento26 páginasReceptorEsteban SmithAún no hay calificaciones
- Sinapsi y Arco Reflejo ExposicionDocumento4 páginasSinapsi y Arco Reflejo ExposicionAlbaneth De CaireAún no hay calificaciones
- Ao-2 MFH IiDocumento11 páginasAo-2 MFH Iiyosman lópezAún no hay calificaciones
- Tema 56 6.2 Plantilla The GlobeDocumento14 páginasTema 56 6.2 Plantilla The Globeismael palmaAún no hay calificaciones
- Reflejo Medular: ClasificaciónDocumento6 páginasReflejo Medular: ClasificaciónJoab Lira DiazAún no hay calificaciones
- 2constitución Del Sistema Nervioso PeriféricoDocumento7 páginas2constitución Del Sistema Nervioso PeriféricoJeancelis SantamaríaAún no hay calificaciones
- Fisiologia de Los Receptores Nerviosos Perifericos (I)Documento18 páginasFisiologia de Los Receptores Nerviosos Perifericos (I)Marco Almeida100% (2)
- Receptores CutaneosDocumento3 páginasReceptores CutaneosYahaira GutierrezAún no hay calificaciones
- Reflejo MedularDocumento1 páginaReflejo MedulargaaaabiiiiiAún no hay calificaciones
- Cuestionario #3Documento8 páginasCuestionario #3Jimena Obregon GarciaAún no hay calificaciones
- FisiologiaDocumento21 páginasFisiologiaNery Javier MartinezAún no hay calificaciones
- Arco ReflejoDocumento12 páginasArco ReflejoluikarenAún no hay calificaciones
- Receptor Sensorial - Wikipedia, La Enciclopedia LibreDocumento27 páginasReceptor Sensorial - Wikipedia, La Enciclopedia LibreEmeron HoriangelizAún no hay calificaciones
- Receptores CinestésicosDocumento10 páginasReceptores CinestésicosGuzz Jozz D KAún no hay calificaciones
- Capitulo 55 Funciones Motoras de La Medula EspinalDocumento10 páginasCapitulo 55 Funciones Motoras de La Medula EspinalEliezer Sop0% (1)
- Taller TFNPDocumento12 páginasTaller TFNPVane CampoAún no hay calificaciones
- Decir ARCO REFLEJODocumento5 páginasDecir ARCO REFLEJORonaldo Cañas BarreiroAún no hay calificaciones
- El Reflejo RotulianoDocumento10 páginasEl Reflejo RotulianoKlarise VanelaAún no hay calificaciones
- Fibras Nerviosas Cap. 3 SnellDocumento7 páginasFibras Nerviosas Cap. 3 Snelldaniela roldan pinoAún no hay calificaciones
- Capítulo 55Documento5 páginasCapítulo 55Cynthia GonzálezAún no hay calificaciones
- Contro Animales PDFDocumento18 páginasContro Animales PDFMarina SoteloAún no hay calificaciones
- Los ReflejosDocumento18 páginasLos ReflejosAdriana Lucia MeekAún no hay calificaciones
- Arco Reflejo y 1 y 2 MotoneuronaDocumento6 páginasArco Reflejo y 1 y 2 MotoneuronaGloria RamosAún no hay calificaciones
- Sistema NerviosoDocumento43 páginasSistema Nerviosobelen.lopez lopezAún no hay calificaciones
- Tema 2. La NeuronaDocumento13 páginasTema 2. La NeuronaJose García SegoviaAún no hay calificaciones
- GUIA 13 Naturales 8Documento6 páginasGUIA 13 Naturales 8Manuel PlataAún no hay calificaciones
- REFLEJODocumento14 páginasREFLEJOEstefaniAún no hay calificaciones
- Tema 10 Reflejos IDocumento4 páginasTema 10 Reflejos IGlenda G. OrtizAún no hay calificaciones
- Practica FisioG 6Documento11 páginasPractica FisioG 6Antoinette MontannaAún no hay calificaciones
- Arco ReflejoDocumento6 páginasArco ReflejoAnna IbarraAún no hay calificaciones
- PRÁCTICA REFLEJOS IncompletaDocumento7 páginasPRÁCTICA REFLEJOS IncompletaLucía SantosAún no hay calificaciones
- Resumen Capitulo 47Documento7 páginasResumen Capitulo 47Litza Naomi SantosAún no hay calificaciones
- Science 2Documento1 páginaScience 2David PhillipsAún no hay calificaciones
- Misión, Visión y Valores - Causa 350 PDFDocumento6 páginasMisión, Visión y Valores - Causa 350 PDFDavid PhillipsAún no hay calificaciones
- Protocolo Proceso de Reclutamiento Docente 2019Documento8 páginasProtocolo Proceso de Reclutamiento Docente 2019David PhillipsAún no hay calificaciones
- ¿Qué Es La Cardiopatía Isquémica - ¿Cómo Se DiagnosticaDocumento29 páginas¿Qué Es La Cardiopatía Isquémica - ¿Cómo Se DiagnosticaDavid PhillipsAún no hay calificaciones
- Piloto 1.0Documento33 páginasPiloto 1.0David PhillipsAún no hay calificaciones
- 18Documento3 páginas18David PhillipsAún no hay calificaciones
- Informe Encuesta Opinión Uso Uniforme Escolar San Viator 2022-2023 PDFDocumento36 páginasInforme Encuesta Opinión Uso Uniforme Escolar San Viator 2022-2023 PDFDavid PhillipsAún no hay calificaciones
- Descargar ComprobanteDocumento1 páginaDescargar ComprobanteDavid PhillipsAún no hay calificaciones
- 234 Presentación de La CarreraDocumento17 páginas234 Presentación de La CarreraDavid PhillipsAún no hay calificaciones
- Programa TEE-028Documento8 páginasPrograma TEE-028David PhillipsAún no hay calificaciones
- ¿Cómo Se Previene La Preeclampsia en Mujeres de Alto Riesgo Con El Uso de Aspirina Profiláctica y OtDocumento11 páginas¿Cómo Se Previene La Preeclampsia en Mujeres de Alto Riesgo Con El Uso de Aspirina Profiláctica y OtDavid PhillipsAún no hay calificaciones
- Guía 1. P. Zavala, P. Donoso, F. DíazDocumento6 páginasGuía 1. P. Zavala, P. Donoso, F. DíazDavid PhillipsAún no hay calificaciones
- Pasaje Ovalle StgoDocumento1 páginaPasaje Ovalle StgoDavid PhillipsAún no hay calificaciones
- Apuntes de NeuroVarios - Felipe Díaz QuintanaDocumento54 páginasApuntes de NeuroVarios - Felipe Díaz QuintanaDavid PhillipsAún no hay calificaciones
- Guía Trabajo 1 - PaulinaDocumento5 páginasGuía Trabajo 1 - PaulinaDavid PhillipsAún no hay calificaciones
- Resumen de El Sistema Sensoriomotriz Parte IDocumento3 páginasResumen de El Sistema Sensoriomotriz Parte IDavid PhillipsAún no hay calificaciones
- Tridimensionalidad Educador@s Grupal MJPalmeroDocumento2 páginasTridimensionalidad Educador@s Grupal MJPalmeroDavid PhillipsAún no hay calificaciones
- HipoDocumento1 páginaHipoDavid PhillipsAún no hay calificaciones
- Proveedores StepsDocumento1 páginaProveedores StepsDavid PhillipsAún no hay calificaciones
- Sra. NancyDocumento4 páginasSra. NancyDavid PhillipsAún no hay calificaciones
- Proyecto Ley Tabaco Marzo 2019 (Borrador)Documento11 páginasProyecto Ley Tabaco Marzo 2019 (Borrador)David PhillipsAún no hay calificaciones
- Fisiología Del Musculo Cardiaco en Bufo Bufo Como Modelo ExperimentalDocumento6 páginasFisiología Del Musculo Cardiaco en Bufo Bufo Como Modelo ExperimentalDiego Jose Campos LeonAún no hay calificaciones
- BAZO. InmunologiaDocumento3 páginasBAZO. InmunologiaJuan Manuel García GutíerrezAún no hay calificaciones
- Tema 2 2019 2020Documento19 páginasTema 2 2019 2020Cristina D.Aún no hay calificaciones
- Hipoxemia Fisioterapia PDFDocumento15 páginasHipoxemia Fisioterapia PDFJulian LondoñoAún no hay calificaciones
- Respuestas A Neumo MycDocumento16 páginasRespuestas A Neumo MycJean Pierre Carrion ArcelaAún no hay calificaciones
- Cardiologia Apuntes Completos Ano 20142015Documento269 páginasCardiologia Apuntes Completos Ano 20142015Adrian Garcia MAún no hay calificaciones
- Fisiología CarlosDocumento59 páginasFisiología Carlosscarcast07Aún no hay calificaciones
- Practica Respiración MitocondrialDocumento9 páginasPractica Respiración MitocondrialEdgar Asael Garcia Garcia100% (1)
- Ejercicios de OxidacionDocumento3 páginasEjercicios de OxidacionAngel ManriqueAún no hay calificaciones
- Infografia Linfocito TDocumento1 páginaInfografia Linfocito TDaniela FloresAún no hay calificaciones
- Sistema Cardio VascularDocumento11 páginasSistema Cardio VascularValentina AmadoAún no hay calificaciones
- Signos Vitales Resumen Semiologia MedicaDocumento13 páginasSignos Vitales Resumen Semiologia MedicaJinson Mogrovejo ValladarezAún no hay calificaciones
- Triptico Del Aparato ExcretorDocumento2 páginasTriptico Del Aparato Excretorvania valeshka coylaAún no hay calificaciones
- El CorazónDocumento5 páginasEl CorazónEdgar MatailoAún no hay calificaciones
- Presion Arterial Jade Ibáñez, Emmy CarrilloDocumento6 páginasPresion Arterial Jade Ibáñez, Emmy CarrillosandiaAún no hay calificaciones
- Insuficiencia Cardiaca CongestivaDocumento24 páginasInsuficiencia Cardiaca Congestivamario1520Aún no hay calificaciones
- Potencial de Reposo (Biofisica)Documento4 páginasPotencial de Reposo (Biofisica)Rafo CastroAún no hay calificaciones
- Sindromes CoronariosDocumento65 páginasSindromes Coronariosdgovea_domingez6012Aún no hay calificaciones
- Infografia Del Cuerpo HumanoDocumento2 páginasInfografia Del Cuerpo HumanoMICHAEL ANTONIO SEL SANDOVALAún no hay calificaciones
- Soporte Vital Básico NeonatalDocumento21 páginasSoporte Vital Básico NeonatalMaiara FrancoAún no hay calificaciones
- Signos Vitales, Lesiones Elementales de La Piel y NevosDocumento3 páginasSignos Vitales, Lesiones Elementales de La Piel y NevosLissette AlcántaraAún no hay calificaciones
- Infografia Humoral Vs CelularDocumento1 páginaInfografia Humoral Vs Celularmiguel jimenezAún no hay calificaciones
- NARCOTICOSDocumento8 páginasNARCOTICOSMonserrat ValadezAún no hay calificaciones
- Manual de Emergencias Profesor Manuel Quesada 2023 2Documento69 páginasManual de Emergencias Profesor Manuel Quesada 2023 2Jocselyn Pérez NacaradoAún no hay calificaciones
- Bioptron Luz PolarizadaDocumento6 páginasBioptron Luz PolarizadaAbŗąĥăm ČŗuzAún no hay calificaciones
- Informe 2 Neurofisiologia FinalDocumento15 páginasInforme 2 Neurofisiologia FinalCristhian García SotoAún no hay calificaciones