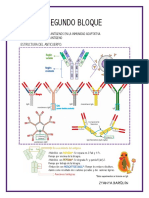
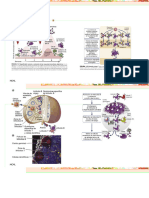
También podría gustarte
- Sueroterapia 1 PDFDocumento15 páginasSueroterapia 1 PDFJudasPriest100% (1)
- Guía Visual de Enfermedades Infecciosas PDFDocumento560 páginasGuía Visual de Enfermedades Infecciosas PDFJacinto Luque Aguilar100% (7)
- Interrelación Entre La NutriciónDocumento5 páginasInterrelación Entre La NutriciónGloria SalasAún no hay calificaciones
- Inmunoglobulinas 2010Documento36 páginasInmunoglobulinas 2010Esteban J Higuera-LizcanoAún no hay calificaciones
- Ensayo B02 4MDocumento31 páginasEnsayo B02 4MCony100% (1)
- Apuntes Del Tema de AnticuerposDocumento7 páginasApuntes Del Tema de AnticuerposPaola PintoAún no hay calificaciones
- Anual Decimo CCNN 2019-2020Documento12 páginasAnual Decimo CCNN 2019-2020damianAún no hay calificaciones
- Unidad 2 2022Documento112 páginasUnidad 2 2022alejandroescobar264Aún no hay calificaciones
- Estructura de Las Igs PDFDocumento14 páginasEstructura de Las Igs PDFEnny StrifeAún no hay calificaciones
- Clase - Articuerpos - 20206dDocumento130 páginasClase - Articuerpos - 20206dJA BrocaAún no hay calificaciones
- INMUNOGLOBULINASDocumento22 páginasINMUNOGLOBULINASMacky Alejandra Gómez MedranoAún no hay calificaciones
- InmulogíaDocumento19 páginasInmulogíaIgnacio RotellaAún no hay calificaciones
- Anticuerpos y Respuesta Humoral SecundariaDocumento20 páginasAnticuerpos y Respuesta Humoral SecundariaPabsAún no hay calificaciones
- Inmunoglobulinas y BCRDocumento14 páginasInmunoglobulinas y BCRLucas Francisco Vargas RodriguezAún no hay calificaciones
- InmunoglobulinasDocumento13 páginasInmunoglobulinasgiovanna ybarraAún no hay calificaciones
- Cadenas Pesadas y LigerasDocumento7 páginasCadenas Pesadas y LigerasGisbelly PayampsAún no hay calificaciones
- Anticuerpos InmunologíaDocumento3 páginasAnticuerpos InmunologíadieegoavromAún no hay calificaciones
- Wuolah Free Tema 3Documento14 páginasWuolah Free Tema 3JESUS LADRON DE GUEVARA MIRANDAAún no hay calificaciones
- Mapa Mental Anticuerpos.Documento6 páginasMapa Mental Anticuerpos.Brenda VergaraAún no hay calificaciones
- PowerPoint PresentationDocumento44 páginasPowerPoint PresentationHenry Ponce TamayAún no hay calificaciones
- REGUEIRO Capitulo 4Documento9 páginasREGUEIRO Capitulo 4MARIA CELESTEAún no hay calificaciones
- GM KM PresentaciónDocumento40 páginasGM KM PresentaciónJavierRodriguezAún no hay calificaciones
- 6 TCR BCR Igs y ToleranciaDocumento14 páginas6 TCR BCR Igs y ToleranciaGuille OliveraAún no hay calificaciones
- EstructuraDocumento8 páginasEstructuraZDKJFGHAún no hay calificaciones
- 05 - Receptores BCR y AnticuerposDocumento4 páginas05 - Receptores BCR y Anticuerpossanti2022Aún no hay calificaciones
- Anticuerpos y AntigenosDocumento10 páginasAnticuerpos y AntigenoskatherineAún no hay calificaciones
- Anticuerpos Monoclonales y Su Uso Como Tratamiento de Lupus Eritematoso SistémicoDocumento8 páginasAnticuerpos Monoclonales y Su Uso Como Tratamiento de Lupus Eritematoso SistémicoMargarita López AlvarezAún no hay calificaciones
- Recombinación Sitio Especifica y Recombinación SomáticaDocumento26 páginasRecombinación Sitio Especifica y Recombinación SomáticaEmerson Ynca PalominoAún no hay calificaciones
- Inmunoglobulina MDocumento7 páginasInmunoglobulina MElian HernándezAún no hay calificaciones
- Anticuerpos y Antigenos 1Documento3 páginasAnticuerpos y Antigenos 1mirope caceresAún no hay calificaciones
- RESUMEN de Inmunoglobulinas M FinalDocumento5 páginasRESUMEN de Inmunoglobulinas M FinalMaría IsabelAún no hay calificaciones
- ESTRUCTURA de Los ANTICUERPOSDocumento7 páginasESTRUCTURA de Los ANTICUERPOSJuan LozadaAún no hay calificaciones
- Repaso Capitulo 5-9Documento10 páginasRepaso Capitulo 5-9Araceli hernandez JaimeAún no hay calificaciones
- AnticuerposDocumento15 páginasAnticuerposfrediry david romero aularAún no hay calificaciones
- 4 MHC Y Present AgDocumento76 páginas4 MHC Y Present AgGeorge RojoAún no hay calificaciones
- 10 Inmunoglobulinas e Inmunidad HumoralDocumento27 páginas10 Inmunoglobulinas e Inmunidad HumoralJ. JoseAún no hay calificaciones
- IgbDocumento24 páginasIgbEsteban J Higuera-LizcanoAún no hay calificaciones
- AnticuerposDocumento30 páginasAnticuerposDavid ValdésAún no hay calificaciones
- DiversidadinmuneDocumento46 páginasDiversidadinmuneraphaelAún no hay calificaciones
- InmunlogiaDocumento3 páginasInmunlogiaLópez Venegas Jaziel PaulAún no hay calificaciones
- Antigenos y AnticuerposDocumento34 páginasAntigenos y AnticuerposmariaAún no hay calificaciones
- Linfocitos BDocumento38 páginasLinfocitos Blady laura restrepoAún no hay calificaciones
- Clase02 PDFDocumento11 páginasClase02 PDFNever MindAún no hay calificaciones
- Fundamentos en Inmunohematologà - ADocumento18 páginasFundamentos en Inmunohematologà - Ajuan pablo padgetAún no hay calificaciones
- Zyanopsis de Inmuno 2.0 - Segundo BloqueDocumento30 páginasZyanopsis de Inmuno 2.0 - Segundo BloqueSofía Vanessa Velázquez JuárezAún no hay calificaciones
- Capítulo 5 - TCR y BCR PDFDocumento7 páginasCapítulo 5 - TCR y BCR PDFJorgeAún no hay calificaciones
- 3.3. AnticuerposDocumento139 páginas3.3. AnticuerposGybran VargasAún no hay calificaciones
- Inmunoglobulina eDocumento26 páginasInmunoglobulina eLpMr Marco AntonioAún no hay calificaciones
- AnticuerposDocumento14 páginasAnticuerposMariano Félix DemasiAún no hay calificaciones
- Tema 3 AnticuerposDocumento6 páginasTema 3 AnticuerposJesus MoralesAún no hay calificaciones
- Organización y Expresión de Los Genes Receptores de LinfocitosDocumento32 páginasOrganización y Expresión de Los Genes Receptores de LinfocitosYareth FloresAún no hay calificaciones
- Resumen Del Prmer Parcial de InmunoDocumento19 páginasResumen Del Prmer Parcial de Inmunorocio bogadoAún no hay calificaciones
- AnticuerposDocumento21 páginasAnticuerposGomez UrsuAún no hay calificaciones
- AnticuerposDocumento58 páginasAnticuerposnomade.astralAún no hay calificaciones
- INMUNOGLOBULINASDocumento5 páginasINMUNOGLOBULINASLizbthAún no hay calificaciones
- Clases 9 y 10 - InmunologíaDocumento21 páginasClases 9 y 10 - Inmunologíafernanda.paredes.1Aún no hay calificaciones
- Antigenos (+clase 3)Documento32 páginasAntigenos (+clase 3)Sandra Gisel KunigkAún no hay calificaciones
- Estructura y Funcion Proteica II.Documento6 páginasEstructura y Funcion Proteica II.jesus sanchez coelhoAún no hay calificaciones
- AnticuerpoDocumento6 páginasAnticuerpocarolinaAún no hay calificaciones
- 3 Semana de InmunologiaDocumento53 páginas3 Semana de InmunologiaHector SalazarAún no hay calificaciones
- Linfocitos B e Inmunidad Humoral PDFDocumento7 páginasLinfocitos B e Inmunidad Humoral PDFJennyffer LopezAún no hay calificaciones
- Tema 9 AnticuerposDocumento40 páginasTema 9 AnticuerposDaniel OrtizAún no hay calificaciones
- Inmunidad HumoralultDocumento47 páginasInmunidad Humoralultbn.tarazona.ramirezAún no hay calificaciones
- Psicofármacos II - AnsiolíticosDocumento14 páginasPsicofármacos II - AnsiolíticosCLAUDIA LORENA RAMIREZ FRANCOAún no hay calificaciones
- Atención de Intoxicaciones en UrgenciasDocumento94 páginasAtención de Intoxicaciones en UrgenciasCLAUDIA LORENA RAMIREZ FRANCOAún no hay calificaciones
- Atención de Intoxicaciones en UrgenciasDocumento94 páginasAtención de Intoxicaciones en UrgenciasCLAUDIA LORENA RAMIREZ FRANCOAún no hay calificaciones
- Tiroideos y AntitiroideosDocumento40 páginasTiroideos y AntitiroideosCLAUDIA LORENA RAMIREZ FRANCOAún no hay calificaciones
- Auto InmunidadDocumento24 páginasAuto InmunidadCLAUDIA LORENA RAMIREZ FRANCOAún no hay calificaciones
- PR Hepáticas Anormales 2021.en - EsDocumento17 páginasPR Hepáticas Anormales 2021.en - EsCLAUDIA LORENA RAMIREZ FRANCOAún no hay calificaciones
- Discusion de RCVDocumento24 páginasDiscusion de RCVCLAUDIA LORENA RAMIREZ FRANCOAún no hay calificaciones
- Inmunorreacciones MixtasDocumento23 páginasInmunorreacciones MixtasArlen EscoberAún no hay calificaciones
- Exposición MicrobiologíaDocumento47 páginasExposición MicrobiologíaDIEGO STIVEN GARCIA ACEVEDOAún no hay calificaciones
- Curs Reumatologie PDFDocumento209 páginasCurs Reumatologie PDFanaflorinapadu6458Aún no hay calificaciones
- Complejo Mayor de Histocompatibilidad 2Documento58 páginasComplejo Mayor de Histocompatibilidad 2Raquel Tejada LantiguaAún no hay calificaciones
- Técnicas para El Estudio de Localización y Función de Proteínas y Ácidos NucleicosDocumento3 páginasTécnicas para El Estudio de Localización y Función de Proteínas y Ácidos Nucleicossofia perezAún no hay calificaciones
- Bruno Roxana Prueba PericialDocumento25 páginasBruno Roxana Prueba PericialRamon MartinezAún no hay calificaciones
- Caso Clinico Corregido 3 Nefrologia IiDocumento13 páginasCaso Clinico Corregido 3 Nefrologia IiYosting RojasAún no hay calificaciones
- Glosario Actividad 1 PDFDocumento6 páginasGlosario Actividad 1 PDFDayana VegaAún no hay calificaciones
- Formato Oficial para Iniciar Tramite de Visita A Internos en Los CeferesosDocumento6 páginasFormato Oficial para Iniciar Tramite de Visita A Internos en Los CeferesosAulcy GarciaAún no hay calificaciones
- Tesis VacunasDocumento47 páginasTesis VacunasBrunovvAún no hay calificaciones
- Clase 11 Citoquinas - SeroinmunologíaDocumento80 páginasClase 11 Citoquinas - SeroinmunologíaCésar Morante ChavarryAún no hay calificaciones
- Equipo 4 "Linfopoyesis" ENF02A 20-2Documento16 páginasEquipo 4 "Linfopoyesis" ENF02A 20-2Ruth Cobo RosalesAún no hay calificaciones
- F CV3 3B 1 - Silabo - Microbiologia - 2019 1Documento10 páginasF CV3 3B 1 - Silabo - Microbiologia - 2019 1Jose DanielAún no hay calificaciones
- Clase PPT Bacterias, Virus, InmunidadDocumento45 páginasClase PPT Bacterias, Virus, InmunidadPaulina GonzálezAún no hay calificaciones
- Hematopoyesis ExpoDocumento23 páginasHematopoyesis ExpoDavid PinedoAún no hay calificaciones
- AnafilaxiaDocumento3 páginasAnafilaxiaAmp CiclicoAún no hay calificaciones
- Tema InmunologiaDocumento9 páginasTema InmunologiaJUAN QUINTEROAún no hay calificaciones
- Tesis Pnie-Cancer 28Documento49 páginasTesis Pnie-Cancer 28Isela PérezAún no hay calificaciones
- Guías de Atención Lepra - Malaria - Dengue - Leishmaniasis - Fiebre Amarilla - Enfermedad de Chagas 2007Documento285 páginasGuías de Atención Lepra - Malaria - Dengue - Leishmaniasis - Fiebre Amarilla - Enfermedad de Chagas 2007obelixoAún no hay calificaciones
- Cuidados de Enfermería en Pacientes OncológicosDocumento4 páginasCuidados de Enfermería en Pacientes OncológicosLaTanaScalaAún no hay calificaciones
- Cronograma Biologia Histologia Embriologia GeneticaDocumento4 páginasCronograma Biologia Histologia Embriologia GeneticaestudianteubaAún no hay calificaciones
- COVID-19 FARMACOLOGÍA Seminario 3 OKDocumento51 páginasCOVID-19 FARMACOLOGÍA Seminario 3 OKEhtilMezaPastranaAún no hay calificaciones
- Bases Farmacológicas Del Tratamiento de La Mastitis BovinaDocumento14 páginasBases Farmacológicas Del Tratamiento de La Mastitis BovinaLuisa R. CalderónAún no hay calificaciones
- Vih Sida Harrison 149615 Downloadable 1669274Documento71 páginasVih Sida Harrison 149615 Downloadable 1669274OKELLY JASIEL GRANDA RIVERAAún no hay calificaciones
- 2021.12.03 - Killa, Canino, Perfil VSDocumento2 páginas2021.12.03 - Killa, Canino, Perfil VSLucero DávilaAún no hay calificaciones
- Roche A-Z (2007)Documento198 páginasRoche A-Z (2007)datitoxAún no hay calificaciones