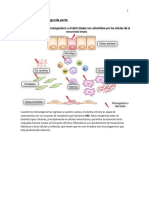
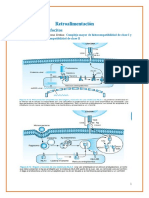
También podría gustarte
- Guía Segundo Parcial InmunologíaDocumento7 páginasGuía Segundo Parcial Inmunologíaanaca59505Aún no hay calificaciones
- Investigaciones InmunologiaDocumento15 páginasInvestigaciones InmunologiaErick Salazar ManzanoAún no hay calificaciones
- Seminario 2. Inmunidad Innata. Parte 2.Documento29 páginasSeminario 2. Inmunidad Innata. Parte 2.lilianabarbagallo.estilistaAún no hay calificaciones
- Inmuno 2Documento36 páginasInmuno 2Al CaralzAún no hay calificaciones
- TEMA 3. La Respuesta Inmune InnataDocumento14 páginasTEMA 3. La Respuesta Inmune InnataNina Souto BlancoAún no hay calificaciones
- Receptores Citosólicos para Pamp y DampDocumento4 páginasReceptores Citosólicos para Pamp y DampPame FloresAún no hay calificaciones
- 3 - PAMPs y RRPDocumento3 páginas3 - PAMPs y RRPGino MartinezAún no hay calificaciones
- La Enfermedad Del Sueño Tiene Sus Orígenes en La Región Afrotropical Del ContinenteDocumento4 páginasLa Enfermedad Del Sueño Tiene Sus Orígenes en La Región Afrotropical Del ContinentejordanAún no hay calificaciones
- Inmunidad InnataDocumento22 páginasInmunidad InnataIsabellaAún no hay calificaciones
- Inmunidad de MucosasDocumento13 páginasInmunidad de MucosasIGNACIO SALVADOR FERNANDEZ GIRONAún no hay calificaciones
- Resumen Respuesta InmunitariaDocumento9 páginasResumen Respuesta InmunitariaMishVencesAún no hay calificaciones
- Sesion 2Documento109 páginasSesion 2Diego LópezAún no hay calificaciones
- Fichas 7Documento7 páginasFichas 7Noelita RiveraAún no hay calificaciones
- Sistema Fagocitico MononuclearDocumento66 páginasSistema Fagocitico MononuclearJose Luis Calle Quispe80% (5)
- T4 InmunoDocumento21 páginasT4 InmunoCris SánchezAún no hay calificaciones
- Actividad Receptores de PRRDocumento11 páginasActividad Receptores de PRRAndres PelayoAún no hay calificaciones
- Chopita - Inmunidad InnataDocumento9 páginasChopita - Inmunidad InnataMartita JuárezAún no hay calificaciones
- Resumen - Generalidades Del Reconocimiento Por La Inmunidad InnataDocumento7 páginasResumen - Generalidades Del Reconocimiento Por La Inmunidad InnataRanciovichAún no hay calificaciones
- Resumen Inmunidad Secretora GRUPO 3Documento10 páginasResumen Inmunidad Secretora GRUPO 3De La Cruz JordanAún no hay calificaciones
- RESPUESTA INNATADocumento4 páginasRESPUESTA INNATAAndrea Mariel PeñaAún no hay calificaciones
- Resumen Capitulo 4 Inmunología Basica AbbasDocumento8 páginasResumen Capitulo 4 Inmunología Basica Abbaswig in a boxAún no hay calificaciones
- Resumen Inmunologia AcoDocumento135 páginasResumen Inmunologia AcoManuel González AlayesAún no hay calificaciones
- El Papel de Los Receptores de Reconocimiento de Patrones en La Inmunidad InnataDocumento46 páginasEl Papel de Los Receptores de Reconocimiento de Patrones en La Inmunidad InnataKArdo23Aún no hay calificaciones
- CITOTOXICIDAD NK Y CTLDocumento2 páginasCITOTOXICIDAD NK Y CTLOctavio ZmAún no hay calificaciones
- Innata 2Documento46 páginasInnata 2henriquecastelanAún no hay calificaciones
- URP Seminario 03 Receptores SolublesDocumento13 páginasURP Seminario 03 Receptores SolublesYébeny Tufino AlvaradoAún no hay calificaciones
- Células, Tejidos y Órganos Del Sistema InmuneDocumento12 páginasCélulas, Tejidos y Órganos Del Sistema InmuneRocío StürtzAún no hay calificaciones
- Respuesta Inmune Innata - 1ra ParteDocumento18 páginasRespuesta Inmune Innata - 1ra Parteirving hilarioAún no hay calificaciones
- Citotoxicidad Mediada Por CélulasDocumento6 páginasCitotoxicidad Mediada Por CélulasSuanny ConsuegraAún no hay calificaciones
- BARRERAS EPITELIALES Y FAGOCITOSIS: DEFENSAS DE LA INMUNIDAD INNATADocumento12 páginasBARRERAS EPITELIALES Y FAGOCITOSIS: DEFENSAS DE LA INMUNIDAD INNATAMariana CaraveoAún no hay calificaciones
- Clase 26 - 8. Inmunidad Innata IDocumento4 páginasClase 26 - 8. Inmunidad Innata IElías Damián Martínez TerénAún no hay calificaciones
- Ensayo 2 InmunologiaDocumento8 páginasEnsayo 2 InmunologiaGaddiel ibarraAún no hay calificaciones
- Desórdenes en Receptores CelularesDocumento40 páginasDesórdenes en Receptores CelularesMarco MadridAún no hay calificaciones
- Receptores Citosólicos para PAMPY DAMP 1Documento11 páginasReceptores Citosólicos para PAMPY DAMP 1BiiancaRodriguezAún no hay calificaciones
- INMUNIDADDocumento9 páginasINMUNIDADLizenia BTAún no hay calificaciones
- FagocitosisDocumento32 páginasFagocitosisMagui DanyAún no hay calificaciones
- Cuestionario Rii EgsDocumento6 páginasCuestionario Rii EgsStephanie RoblesAún no hay calificaciones
- Inmunidad InnataDocumento17 páginasInmunidad InnataWraysthell CordovaAún no hay calificaciones
- Fagocitosis, proceso de ingestión de patógenosDocumento10 páginasFagocitosis, proceso de ingestión de patógenosJuliana AmayaAún no hay calificaciones
- Receptores de La Inmunidad Innata, Otros Receptores para El Reconocimiento de Patrón y Receptores de La Inmunidad AdaptativaDocumento8 páginasReceptores de La Inmunidad Innata, Otros Receptores para El Reconocimiento de Patrón y Receptores de La Inmunidad AdaptativaDonia Abou ArabAún no hay calificaciones
- Tema 3 Antígenos, Receptores y Barreras Del SIDocumento38 páginasTema 3 Antígenos, Receptores y Barreras Del SIFernando Gual BarrosoAún no hay calificaciones
- inmuno borradorDocumento8 páginasinmuno borradorjuan guamanAún no hay calificaciones
- Mhs Tramo Dos SisamDocumento73 páginasMhs Tramo Dos SisamZiaraAún no hay calificaciones
- Receptores de los linfocitos B y TDocumento11 páginasReceptores de los linfocitos B y TPaola PintoAún no hay calificaciones
- Inmunidad InnataDocumento14 páginasInmunidad InnataMelissa RodriguezAún no hay calificaciones
- 5.clase 7 - 9 Inmunidad Innata IIIDocumento10 páginas5.clase 7 - 9 Inmunidad Innata IIIElías Damián Martínez TerénAún no hay calificaciones
- Receptores CitosolicosDocumento33 páginasReceptores CitosolicosVanessa YapoAún no hay calificaciones
- Inmunidad InnataDocumento38 páginasInmunidad InnataAnyello BuitragoAún no hay calificaciones
- Mecanismos Generales de Comunicación CelularDocumento4 páginasMecanismos Generales de Comunicación CelularPatricia SolanoAún no hay calificaciones
- Enfermedades InmunesDocumento25 páginasEnfermedades InmunesIsabella TabaresAún no hay calificaciones
- Presentacion Receptores Citosolicos y de MembranaDocumento41 páginasPresentacion Receptores Citosolicos y de Membranaisabella vAún no hay calificaciones
- inmunologia de lepraDocumento8 páginasinmunologia de lepraEfrain Teran CarbonoAún no hay calificaciones
- 169 Sistema Inmunitario Cutáneo PDFDocumento63 páginas169 Sistema Inmunitario Cutáneo PDFJ C Torres Formalab100% (1)
- Presentacion de AntigenosDocumento4 páginasPresentacion de Antigenoscami_0724Aún no hay calificaciones
- Capítulo 8Documento7 páginasCapítulo 8Mayeli UrrutiaAún no hay calificaciones
- Activación de Linfocitos T y B: Reconocimiento Antigénico y SeñalizaciónDocumento22 páginasActivación de Linfocitos T y B: Reconocimiento Antigénico y SeñalizaciónDove CameronAún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Parque AviacionDocumento17 páginasParque AviacionReyner CovarrubiasAún no hay calificaciones
- Con Solid A Do Partida Me Tra DoDocumento6 páginasCon Solid A Do Partida Me Tra DoRobbyn QSAún no hay calificaciones
- Pre Supuesto Client eDocumento1 páginaPre Supuesto Client evictorAún no hay calificaciones
- Pre Supuesto Client eDocumento1 páginaPre Supuesto Client evictorAún no hay calificaciones
- Pre Supuesto Client eDocumento1 páginaPre Supuesto Client evictorAún no hay calificaciones
- Vitaminas PresentacionDocumento13 páginasVitaminas PresentaciondeibyAún no hay calificaciones
- Semana 9. Metabolismo de Los Ácidos Grasos, Biosíntesis de Fosfolípidos, Glicólipidos, Colesterol.Documento69 páginasSemana 9. Metabolismo de Los Ácidos Grasos, Biosíntesis de Fosfolípidos, Glicólipidos, Colesterol.Carmen Villegas GermànAún no hay calificaciones
- Proteasas digestivas: Enzimas proteolíticas para digerir proteínasDocumento4 páginasProteasas digestivas: Enzimas proteolíticas para digerir proteínasJonathanAún no hay calificaciones
- MACRONUTRIENTESDocumento5 páginasMACRONUTRIENTESMagaly Perez PerezAún no hay calificaciones
- Caso Clinico N°11Documento20 páginasCaso Clinico N°11LYNDA FERNANDA RODRIGUEZ MEDINAAún no hay calificaciones
- Examen BiotecnologiaDocumento4 páginasExamen BiotecnologiaJuan FigueroaAún no hay calificaciones
- Gluconeogénesis y Metabolismo Del GlucógenoDocumento35 páginasGluconeogénesis y Metabolismo Del GlucógenoLucia Estrada ZuñigaAún no hay calificaciones
- Menú Área Suero Ensayos (Web)Documento4 páginasMenú Área Suero Ensayos (Web)Daniela ReyesAún no hay calificaciones
- Mulgatol JuniorDocumento4 páginasMulgatol JuniorMiriam Vargas DehneAún no hay calificaciones
- Bases Cientificas en El Consumo de Complementos y Suplementos Alimenticios-Magdalena NievesDocumento22 páginasBases Cientificas en El Consumo de Complementos y Suplementos Alimenticios-Magdalena NievesShaneDanielChanEsparzaAún no hay calificaciones
- Practica 1. Reaccion de Aminoacidos y ProteinasDocumento2 páginasPractica 1. Reaccion de Aminoacidos y ProteinasPierdant O-oAún no hay calificaciones
- Carbohidratos, lípidos, vitaminas y metabolismo mineralDocumento10 páginasCarbohidratos, lípidos, vitaminas y metabolismo mineralGreciaAún no hay calificaciones
- La Chia Salvia Hispanica L Desarrollo Alimentos SaludablesDocumento43 páginasLa Chia Salvia Hispanica L Desarrollo Alimentos SaludablesPablo Fernando Abarca PereiraAún no hay calificaciones
- Laboratorio 4Documento7 páginasLaboratorio 4Jahiro Rivas FloresAún no hay calificaciones
- Introducción Extracción Del ADNDocumento2 páginasIntroducción Extracción Del ADNAlejandra TKTAún no hay calificaciones
- Reacciones Enzimáticas de Oxido ReducciónDocumento5 páginasReacciones Enzimáticas de Oxido ReducciónDynarko RuizAún no hay calificaciones
- Simulador Glucólisis y Ciclo de Krebs Bioquímica Parcial 2Documento9 páginasSimulador Glucólisis y Ciclo de Krebs Bioquímica Parcial 2Isaac GómezAún no hay calificaciones
- Historia Del AdnDocumento1 páginaHistoria Del Adnmarco jeffreyAún no hay calificaciones
- PROSTAGLANDINASDocumento33 páginasPROSTAGLANDINASjunieth sequeiraAún no hay calificaciones
- GLUCÓLISISDocumento6 páginasGLUCÓLISISCamila ContrerasAún no hay calificaciones
- Fermentacion HeterolacticaDocumento2 páginasFermentacion HeterolacticaRomina Arévalo0% (1)
- Serie Didáctica Qca de La VidaDocumento104 páginasSerie Didáctica Qca de La VidaCarlos Alfredo LópezAún no hay calificaciones
- Biotina y MarcadoresDocumento8 páginasBiotina y MarcadoresNaty Liz Cantuña100% (1)
- Carbohidratos Glúcidos: Marta Gutiérrez Del CampoDocumento26 páginasCarbohidratos Glúcidos: Marta Gutiérrez Del CampoHéctor MejiasAún no hay calificaciones
- Modulo Ciencias Naturales Ciclo V Grado DecimoDocumento99 páginasModulo Ciencias Naturales Ciclo V Grado DecimoJuan CarlosAún no hay calificaciones
- VitaminasDocumento25 páginasVitaminasAde RendónAún no hay calificaciones
- Bioquimica EjerciciosDocumento2 páginasBioquimica EjerciciosSteven Cristofer Lopez Carbajal100% (6)
- 2do PARCIAL DE BIOQUIMICA 2020 OooooDocumento19 páginas2do PARCIAL DE BIOQUIMICA 2020 OooooIdyel T. Cris100% (1)
- Tema 26 - Hemostasia y CoagulaciónDocumento27 páginasTema 26 - Hemostasia y Coagulaciónangye daniela contreras contrerasAún no hay calificaciones
- Los Lípidos para Cuarto de SecundariaDocumento8 páginasLos Lípidos para Cuarto de SecundariaElden CulquicondorAún no hay calificaciones