También podría gustarte
- Mitos Sobre La Depresión Moderada y Grave.Documento8 páginasMitos Sobre La Depresión Moderada y Grave.Alonso HerreraAún no hay calificaciones
- Cuestionario de InmunologiaDocumento8 páginasCuestionario de Inmunologiasergito_pazAún no hay calificaciones
- 14-Inmunidad Frente A ParásitosDocumento16 páginas14-Inmunidad Frente A Parásitosyccardenasm100% (1)
- Consentimiento Hialuron PenDocumento3 páginasConsentimiento Hialuron PenReme Bedoya Santana50% (2)
- Los bacteriófagos: Los virus que combaten infeccionesDe EverandLos bacteriófagos: Los virus que combaten infeccionesAún no hay calificaciones
- Mecanismos de Evasion de La Respuesta InmuneDocumento8 páginasMecanismos de Evasion de La Respuesta InmuneNnznznznAún no hay calificaciones
- AJO en POLVO Hoja de SeguridadDocumento4 páginasAJO en POLVO Hoja de SeguridadKatherine TafurAún no hay calificaciones
- Sistemas de vigilancia alimentaria nutricionalDocumento12 páginasSistemas de vigilancia alimentaria nutricionalPia EscobalAún no hay calificaciones
- Inmunidad Adaptativa.Documento7 páginasInmunidad Adaptativa.Alejandro Dario Bravo BermelloAún no hay calificaciones
- Cap 16Documento36 páginasCap 16Alejandra Obregon100% (3)
- I-5. Mecanismos de EvasiónDocumento10 páginasI-5. Mecanismos de EvasiónEricksen SalazarAún no hay calificaciones
- T 10 Respuesta Inmune A Virus y Bacterias UPSJB 2023-IIDocumento32 páginasT 10 Respuesta Inmune A Virus y Bacterias UPSJB 2023-IIOswaldo Churi Silva AltamiranoAún no hay calificaciones
- Introducción A La FISIOLOGÍA de La RESPUESTA INMUNEDocumento50 páginasIntroducción A La FISIOLOGÍA de La RESPUESTA INMUNEGabrielAún no hay calificaciones
- Tema 24 Inmunidad BacteriasDocumento12 páginasTema 24 Inmunidad BacteriasAna María RodrigoAún no hay calificaciones
- Inmuno Logia Apuntes Mecanismos de EvasionDocumento7 páginasInmuno Logia Apuntes Mecanismos de EvasionEDGAR AXEL GONZALEZ DUENASAún no hay calificaciones
- Introducción A La InmunologíaDocumento1 páginaIntroducción A La InmunologíaYami MoralesAún no hay calificaciones
- Expocisión - Mecanismos de Evasión ViralDocumento37 páginasExpocisión - Mecanismos de Evasión ViralLuis Gerardo Camacho PlascenciaAún no hay calificaciones
- Intestinal Nematode Parasites. Mechanism of Resistance (Parásitos Intestinales - Nematodos y Mecanismos de Resistencia)Documento1 páginaIntestinal Nematode Parasites. Mechanism of Resistance (Parásitos Intestinales - Nematodos y Mecanismos de Resistencia)Alejandra Salcedo ReyesAún no hay calificaciones
- Mecanismos Evasion ParasitariaDocumento38 páginasMecanismos Evasion ParasitariaLuke CarsaAún no hay calificaciones
- Inmunidad Antiviral - Bacteriana - Antiparasitaria 2022 ViñaDocumento48 páginasInmunidad Antiviral - Bacteriana - Antiparasitaria 2022 ViñaEnfermería UNABAún no hay calificaciones
- Taller Parasitologia Juan Rozo-Kelly LopezDocumento8 páginasTaller Parasitologia Juan Rozo-Kelly LopezJuan Felipe Rozo MorenoAún no hay calificaciones
- CAPITULO 1 Introducción Al Sistema InmunitarioDocumento17 páginasCAPITULO 1 Introducción Al Sistema InmunitarioNatanael TGonzAún no hay calificaciones
- Taller Parasitologia Juan Rozo-Kelly LopezDocumento8 páginasTaller Parasitologia Juan Rozo-Kelly LopezJuan Felipe Rozo MorenoAún no hay calificaciones
- Grupo F - Cuestionario 04 - Acuña Pérez Juan JesúsDocumento7 páginasGrupo F - Cuestionario 04 - Acuña Pérez Juan JesúsJuan Jesús Acuña PėrezAún no hay calificaciones
- Mecanismos de EvasiónDocumento5 páginasMecanismos de EvasiónADRIANA PAOLA GARCIA BARAJASAún no hay calificaciones
- 02 - Inmunidad Innata - NNPDocumento54 páginas02 - Inmunidad Innata - NNPGilbert Canedo AvilaAún no hay calificaciones
- Expo InmunoDocumento20 páginasExpo InmunoDiego Torres AltamiranoAún no hay calificaciones
- Evasion de La Respuesta Inmune (Parasitos)Documento28 páginasEvasion de La Respuesta Inmune (Parasitos)GuzMan ZCarlozAún no hay calificaciones
- Diapositivas Mecanismos de Defensa Contra HongosDocumento19 páginasDiapositivas Mecanismos de Defensa Contra HongosSteph VivasAún no hay calificaciones
- Cap 16 InmunologiaDocumento5 páginasCap 16 InmunologiamartinvillarrealuasmedicinaAún no hay calificaciones
- TRABAJO PRACTICO de INMUNIDAD INNATADocumento5 páginasTRABAJO PRACTICO de INMUNIDAD INNATALAURAAún no hay calificaciones
- SEMANA 6 Inmunidad Tumoral y ViralDocumento70 páginasSEMANA 6 Inmunidad Tumoral y ViralAnne NikolAún no hay calificaciones
- Resumen Inmunologia Teorico 2020Documento158 páginasResumen Inmunologia Teorico 2020Romy PechAún no hay calificaciones
- LN Cap 1.35Documento38 páginasLN Cap 1.35Daniela Patricia Alvarez AravenaAún no hay calificaciones
- Generalidades Sobre El Sistema InmunitarioDocumento30 páginasGeneralidades Sobre El Sistema InmunitarioMiguel TaveraAún no hay calificaciones
- Mecanismo de EvasionDocumento21 páginasMecanismo de EvasionDeysiAún no hay calificaciones
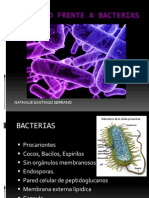
- Inmunidad Frente A BacteriasDocumento23 páginasInmunidad Frente A BacteriasNathalie SantiagoAún no hay calificaciones
- Generalidades Del Sistema InmuneDocumento3 páginasGeneralidades Del Sistema InmuneTahiana De La CruzAún no hay calificaciones
- Clase 2Documento48 páginasClase 2aracelyjimenez460Aún no hay calificaciones
- Ehrlichia CanisDocumento5 páginasEhrlichia CanisNaruto ShikamaruAún no hay calificaciones
- Clase 1 Inmunidad Innata Parte IDocumento9 páginasClase 1 Inmunidad Innata Parte ILiliana SolariAún no hay calificaciones
- Inmunologos I y Ii ParcialDocumento108 páginasInmunologos I y Ii ParcialJazmin MateosAún no hay calificaciones
- A Resposta Inmunitaria Frente A ParásitosDocumento5 páginasA Resposta Inmunitaria Frente A Parásitoscolordeestrellas92Aún no hay calificaciones
- Mecanismos de Evasión de La Respuesta Inmunitaria en Los Parásitos SantyDocumento4 páginasMecanismos de Evasión de La Respuesta Inmunitaria en Los Parásitos SantyJORGE SANTIAGO LAZCANO SHERMANAún no hay calificaciones
- Evasion Inmunitaria Por Parte de Los ParasitosDocumento4 páginasEvasion Inmunitaria Por Parte de Los ParasitosSandra VilloAún no hay calificaciones
- Funciones Efectoras de Las CitoquinasDocumento31 páginasFunciones Efectoras de Las Citoquinaskaterin maiguaAún no hay calificaciones
- Inmunologia General UPSJBDocumento53 páginasInmunologia General UPSJBAnita LopezAún no hay calificaciones
- Inmunology TuberculosisDocumento50 páginasInmunology Tuberculosisfrancisco salazar huamaniAún no hay calificaciones
- Gabriel Hernandez Fuentes Respuesta InmunitariaDocumento6 páginasGabriel Hernandez Fuentes Respuesta InmunitariaGabriel HernándezAún no hay calificaciones
- S.2 Micro Oral (Pregunta y Respuesta)Documento12 páginasS.2 Micro Oral (Pregunta y Respuesta)Cristian Pincheira ArayaAún no hay calificaciones
- Mecanismos de Respuesta InmuneDocumento21 páginasMecanismos de Respuesta InmuneJocelyn GualAún no hay calificaciones
- 12 Resistencia A Bacterias y HongosDocumento5 páginas12 Resistencia A Bacterias y HongosValeria Dos SantosAún no hay calificaciones
- INTRODUCCION A LA INMUNOLOGIA y RII 2017Documento75 páginasINTRODUCCION A LA INMUNOLOGIA y RII 2017Christian MartinoniAún no hay calificaciones
- Inmunología Regueiro Cap 18Documento10 páginasInmunología Regueiro Cap 18Natalia Espíndola BalderasAún no hay calificaciones
- Inmunidad Frente A Patógenos Intracelulares 2016Documento63 páginasInmunidad Frente A Patógenos Intracelulares 2016Daii YJonaAún no hay calificaciones
- Inmunología de La Infección Por HelmintosDocumento19 páginasInmunología de La Infección Por HelmintosYhulia Racchell Del Cid100% (1)
- Clase 2 Respuesta Inmune IDocumento60 páginasClase 2 Respuesta Inmune IPierinna CarrascoAún no hay calificaciones
- Respuesta Inmune Contra Infecciones Por Protozoarios y ElmintosDocumento18 páginasRespuesta Inmune Contra Infecciones Por Protozoarios y ElmintosFranklin garryAún no hay calificaciones
- Cap 6 Resumen Patologia Estructural y FuncionalDocumento38 páginasCap 6 Resumen Patologia Estructural y FuncionalMariela GuzmanAún no hay calificaciones
- CAPÍTULO 12 - Respuestas Efectoras - Inmunidad Mediada Por Anticuerpos y Por CélulasDocumento54 páginasCAPÍTULO 12 - Respuestas Efectoras - Inmunidad Mediada Por Anticuerpos y Por CélulasFernanda López LópezAún no hay calificaciones
- 2 INMUNUOLOGIA - Gus - P2 - 2Documento56 páginas2 INMUNUOLOGIA - Gus - P2 - 2Julieta suarezAún no hay calificaciones
- Inmunidad Frente A Hongos y ParásitosDocumento3 páginasInmunidad Frente A Hongos y ParásitosMari CorderoAún no hay calificaciones
- 1 Inmunología GeneralidadesDocumento41 páginas1 Inmunología GeneralidadesdragoncytoAún no hay calificaciones
- Universidad VeracruzanaDocumento11 páginasUniversidad VeracruzanaAlonso AAún no hay calificaciones
- Ictericia PowerDocumento46 páginasIctericia PowerAlonso AAún no hay calificaciones
- DiagramaDocumento1 páginaDiagramaAlonso AAún no hay calificaciones
- DiagramaDocumento1 páginaDiagramaAlonso AAún no hay calificaciones
- Artículo EspecialDocumento10 páginasArtículo EspecialAlonso AAún no hay calificaciones
- 22 Uara 1Documento4 páginas22 Uara 1Alonso AAún no hay calificaciones
- Menu Martintxo CorregidoDocumento16 páginasMenu Martintxo CorregidoMUNIECOFANTASMAAún no hay calificaciones
- 3RO Sesion de Clases Entero 1Documento9 páginas3RO Sesion de Clases Entero 1willan piero chavez zoteloAún no hay calificaciones
- Semana 3 Cultivo de PecesDocumento2 páginasSemana 3 Cultivo de PecesPaola ClementeAún no hay calificaciones
- Cómo Se Crea La Suerte Inteligente para Sacarle El Máximo Provecho A Lo InesperadoDocumento22 páginasCómo Se Crea La Suerte Inteligente para Sacarle El Máximo Provecho A Lo InesperadoLo Con100% (1)
- Caso Clinico - Insuficiencia Renal AgudaDocumento5 páginasCaso Clinico - Insuficiencia Renal AgudaNAZARETH DAHANA ORTIZ NUÑEZAún no hay calificaciones
- 3 Concepto Castillo MoralesDocumento12 páginas3 Concepto Castillo MoralesGianfraco BravoAún no hay calificaciones
- Proyecto HierroDocumento18 páginasProyecto HierroDanicho Lara100% (1)
- Adherencia BorradorDocumento23 páginasAdherencia BorradorLissette PérezAún no hay calificaciones
- Resumen Tarea 9. Analisis Biomecanico de La Marcha - Maria BermudezDocumento16 páginasResumen Tarea 9. Analisis Biomecanico de La Marcha - Maria BermudezMaria BermudezAún no hay calificaciones
- Presentación Manejo Del EstrésDocumento24 páginasPresentación Manejo Del EstrésDiego Sierra100% (2)
- Procesos y Definiciones SMAWDocumento5 páginasProcesos y Definiciones SMAWPROYECTOS GRUPO VIRUAún no hay calificaciones
- NTC4574SalmonellaDocumento39 páginasNTC4574SalmonellalabconcldAún no hay calificaciones
- Ficha Técnica Caneca Plásica 55 Galones 01 TAI 208 11 Ene 20Documento1 páginaFicha Técnica Caneca Plásica 55 Galones 01 TAI 208 11 Ene 20Julio BuendiaAún no hay calificaciones
- 1.b. Compostaje Como Una Forma - Español PDFDocumento48 páginas1.b. Compostaje Como Una Forma - Español PDFCarolina Contreras CastroAún no hay calificaciones
- Fdaa 50Documento146 páginasFdaa 50Irving RodriguezAún no hay calificaciones
- Investigación de Operaciones - T1Documento28 páginasInvestigación de Operaciones - T1Yhurema D'l Carmen Geldres TorresAún no hay calificaciones
- Curr Vitae21Documento7 páginasCurr Vitae21Esteban Choque CondeAún no hay calificaciones
- El Cuestionario de Experiencias Relacionadas Con Los Videjuegos (Cerv) PDFDocumento9 páginasEl Cuestionario de Experiencias Relacionadas Con Los Videjuegos (Cerv) PDFEliezer Araya PizarroAún no hay calificaciones
- 2015 MILANO y HAMUY Intervenciones en La Clinica de NiñosDocumento9 páginas2015 MILANO y HAMUY Intervenciones en La Clinica de NiñosMaggui GómezAún no hay calificaciones
- Aislamiento MecánicoDocumento23 páginasAislamiento MecánicoMárquez Pernia R100% (1)
- Presentacion COPASSTDocumento23 páginasPresentacion COPASSThernan ospino valleAún no hay calificaciones
- Infarto Del Miocardio AgudoDocumento25 páginasInfarto Del Miocardio AgudolizyaraAún no hay calificaciones
- Guia 20Documento6 páginasGuia 20YosaharaVidalMateoAún no hay calificaciones
- FC 24 24Documento28 páginasFC 24 24Christian VarelaAún no hay calificaciones
- Info Biomusica 6 en 1Documento5 páginasInfo Biomusica 6 en 1Pere SantAún no hay calificaciones
- Mapa Mental SIGNODocumento1 páginaMapa Mental SIGNOYanela Cojal Rodriguez100% (1)