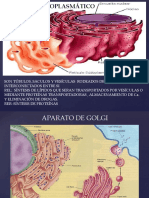
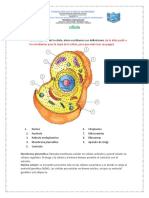
También podría gustarte
- Resumen - Cap 5Documento8 páginasResumen - Cap 5Pamela De LimaAún no hay calificaciones
- Teorico Cã - Mo Estã - N Organizadas Las Cã - Lulas 4-4-23Documento48 páginasTeorico Cã - Mo Estã - N Organizadas Las Cã - Lulas 4-4-23Gabriela IbañezAún no hay calificaciones
- CélulaDocumento18 páginasCélulaABAD TAún no hay calificaciones
- Estructura y Función Celular 1.Documento22 páginasEstructura y Función Celular 1.macavecaAún no hay calificaciones
- Metabolismo VegetalDocumento45 páginasMetabolismo VegetalAlejandro GuzmanAún no hay calificaciones
- Celula, Morfologia CelularDocumento8 páginasCelula, Morfologia Celularscarletlugo4751Aún no hay calificaciones
- Estructura y Funcion de Celulas Procariotas y Eucariotas - Docx2Documento11 páginasEstructura y Funcion de Celulas Procariotas y Eucariotas - Docx2Ixell CastAún no hay calificaciones
- Como Están Organizadas Las CélulasDocumento5 páginasComo Están Organizadas Las Célulasdanny100% (1)
- 1º - Apuntes Nivel CelularDocumento8 páginas1º - Apuntes Nivel CelularAlejandra Rommy Vera CabreraAún no hay calificaciones
- Organelos Membranosos y No Membranosos ScribdDocumento4 páginasOrganelos Membranosos y No Membranosos ScribdJoseAún no hay calificaciones
- Material Audiovisual CélulaDocumento25 páginasMaterial Audiovisual CélulaCristian MendozaAún no hay calificaciones
- Trabajo de Biologia de CelulaDocumento10 páginasTrabajo de Biologia de CelulaHarold BetancourtAún no hay calificaciones
- G1 Estructura y Funcion CelularDocumento14 páginasG1 Estructura y Funcion CelularYaritza AhumadaAún no hay calificaciones
- Apuntes Celula EucariotaDocumento14 páginasApuntes Celula EucariotaGloria Angelica Acuña DelgadoAún no hay calificaciones
- Neurociencias ResumenesDocumento15 páginasNeurociencias ResumenesMelany Aldana DAIYOUBAún no hay calificaciones
- Practica Iii La CelulaDocumento6 páginasPractica Iii La CelulaIglesia Brisas de AmorAún no hay calificaciones
- Citología VegetalDocumento46 páginasCitología VegetalSheraly carrascoAún no hay calificaciones
- Monografia La CelulaDocumento22 páginasMonografia La CelulaJessicaMeléndezAún no hay calificaciones
- Las Celulas Eucariotas y ProcariotasDocumento6 páginasLas Celulas Eucariotas y Procariotasagarta1955Aún no hay calificaciones
- Célula, Función, Morfología y Fisiología.Documento3 páginasCélula, Función, Morfología y Fisiología.marsy royed100% (1)
- La Célula Estructura y OrganizaciónDocumento1 páginaLa Célula Estructura y OrganizaciónArtic AAún no hay calificaciones
- La Célula ExamenDocumento11 páginasLa Célula Examengris431Aún no hay calificaciones
- Celula VegetalDocumento8 páginasCelula VegetalNico SosaAún no hay calificaciones
- Célula Vegetal 2013 PDFDocumento26 páginasCélula Vegetal 2013 PDFivan villarrealAún no hay calificaciones
- 1º - Apuntes Nivel CelularDocumento8 páginas1º - Apuntes Nivel CelularyismelyfmaAún no hay calificaciones
- Citoplasma ImprimirDocumento3 páginasCitoplasma ImprimirAdriana SanchezAún no hay calificaciones
- Apuntes La CélulaDocumento8 páginasApuntes La CélulaLuis TrañaAún no hay calificaciones
- Trabajo La CélulaDocumento16 páginasTrabajo La CélulaMara Gregorina RodriguezAún no hay calificaciones
- RMN - 2D Varios Diapos PDFDocumento32 páginasRMN - 2D Varios Diapos PDFquimica4dfAún no hay calificaciones
- Célula VegetalDocumento6 páginasCélula VegetalMiguel OsorioAún no hay calificaciones
- Estructura y Funcion de Celulas Procariotas y EucariotasDocumento6 páginasEstructura y Funcion de Celulas Procariotas y EucariotasJose Gonzales100% (1)
- Apuntes CélulaDocumento6 páginasApuntes CélulaCésar Humberto Villa DoyharcabalAún no hay calificaciones
- Resumen 2 - APBDocumento5 páginasResumen 2 - APBAlejandro Pesqueira BojorquezAún no hay calificaciones
- Parcial BiologiaDocumento24 páginasParcial BiologialorejojedaAún no hay calificaciones
- Lectura Nº1 Repaso Biología CelularDocumento8 páginasLectura Nº1 Repaso Biología CelularCristian FloresAún no hay calificaciones
- Descripcion de La CelulaDocumento6 páginasDescripcion de La CelulaAntonio Guexpal TepoxAún no hay calificaciones
- Taller Morfología CélulasDocumento6 páginasTaller Morfología Célulasana_fouAún no hay calificaciones
- Examen Biogia II Bim 2011Documento13 páginasExamen Biogia II Bim 2011Carmen Rosario Villanes PantojaAún no hay calificaciones
- 2021 Temario Ciencias Naturales Segundo CicloDocumento311 páginas2021 Temario Ciencias Naturales Segundo CicloTía Yoli RamírezAún no hay calificaciones
- CelulasDocumento14 páginasCelulasJosue RebolledoAún no hay calificaciones
- Apuntes Nivel Celular AIEPDocumento6 páginasApuntes Nivel Celular AIEPcokimaster100% (1)
- Módulo 6 I La CelulaDocumento77 páginasMódulo 6 I La CelulaVale Jaramillo GilAún no hay calificaciones
- 05 Citoplasma - CitoesqueletoDocumento19 páginas05 Citoplasma - CitoesqueletoAlfredo Sam Alvaro HerreraAún no hay calificaciones
- La Célula. Célula Procariota y EucariotaDocumento1 páginaLa Célula. Célula Procariota y EucariotaFleming Perla100% (1)
- Estructura y Función en Procariotas y EucariotasDocumento87 páginasEstructura y Función en Procariotas y EucariotasempresvarAún no hay calificaciones
- La Unidad de La VidaDocumento14 páginasLa Unidad de La VidaCarlos LuboAún no hay calificaciones
- La Célula VegetalDocumento14 páginasLa Célula VegetalBriam Sting Mallqui EspirituAún no hay calificaciones
- Lectura Sobre La CelulaDocumento7 páginasLectura Sobre La Celulalogan8412100% (3)
- La CélulaDocumento12 páginasLa Célulamartin muñozAún no hay calificaciones
- CelulaDocumento92 páginasCelulaGwen SM100% (10)
- Celula y Sus PartesDocumento11 páginasCelula y Sus PartesJimmy Karo SolorzanoAún no hay calificaciones
- Clase Celula (Autoguardado)Documento70 páginasClase Celula (Autoguardado)127269100% (3)
- 1 CélulaDocumento20 páginas1 CélulaDanna Sofia Argote FuentesAún no hay calificaciones
- Interior de La CélulaDocumento11 páginasInterior de La CélulaAriel PalajAún no hay calificaciones
- Las células, Constructoras de vida: Cells: Constructing Living ThingsDe EverandLas células, Constructoras de vida: Cells: Constructing Living ThingsCalificación: 3.5 de 5 estrellas3.5/5 (2)
- Histología y Embrología del Sitema NerviosoDe EverandHistología y Embrología del Sitema NerviosoCalificación: 4 de 5 estrellas4/5 (4)
- Biología Molecular y Celular: Una guía introductoria para aprender Biología Celular y MolecularDe EverandBiología Molecular y Celular: Una guía introductoria para aprender Biología Celular y MolecularAún no hay calificaciones
- Anemia Para Todos: Medicina Para TodosDe EverandAnemia Para Todos: Medicina Para TodosCalificación: 3 de 5 estrellas3/5 (1)
- Oszlak - La Formación Del Estado ArgentinoDocumento16 páginasOszlak - La Formación Del Estado ArgentinoAnonymous G4vVr3nL100% (3)
- Apunte 11Documento6 páginasApunte 11Lara ReevesAún no hay calificaciones
- BiomoleculasDocumento9 páginasBiomoleculasLara ReevesAún no hay calificaciones
- Apunte 5Documento2 páginasApunte 5Lara ReevesAún no hay calificaciones
- Apunte 7Documento7 páginasApunte 7Lara ReevesAún no hay calificaciones
- Apunte 4Documento10 páginasApunte 4Lara ReevesAún no hay calificaciones
- Apunte 6Documento5 páginasApunte 6Lara ReevesAún no hay calificaciones
- Apunte 3Documento4 páginasApunte 3Lara ReevesAún no hay calificaciones
- AnatomiaDocumento185 páginasAnatomiaTania C Mascareño VargasAún no hay calificaciones
- La Naturaleza Del Mundo MicrobianoDocumento17 páginasLa Naturaleza Del Mundo MicrobianoGustavo CandalloAún no hay calificaciones
- Diap Unidad 13 Apa Golgi-Liso-PeroxDocumento15 páginasDiap Unidad 13 Apa Golgi-Liso-PeroxMaria MaciasAún no hay calificaciones
- La CelulaDocumento19 páginasLa CelulaGamma RiveraAún no hay calificaciones
- Cuestionario N 2 BotanicaDocumento15 páginasCuestionario N 2 BotanicaJose Maria Flores RuizAún no hay calificaciones
- Ticket Paes III Medio Celula Teoria y Estructura ClavesDocumento23 páginasTicket Paes III Medio Celula Teoria y Estructura ClavesVanessa Mitocondria Gonzalez DonosoAún no hay calificaciones
- La Celula y Sus Componentes - OdtDocumento6 páginasLa Celula y Sus Componentes - OdtRodrigo RoderigAún no hay calificaciones
- Unidad 04 - Celula Eucariota Sin RespuestasDocumento11 páginasUnidad 04 - Celula Eucariota Sin RespuestasCinthya Eusebia Quispe PomaAún no hay calificaciones
- Especialización CelularDocumento12 páginasEspecialización CelularValeria MirandaAún no hay calificaciones
- Repaso Biología MolecularDocumento10 páginasRepaso Biología MolecularRodrigo RodríguezAún no hay calificaciones
- Taller N°2, BiologiaDocumento15 páginasTaller N°2, BiologiaMIGUEL ANGEL SUAZA ESPINOSAAún no hay calificaciones
- Universidad Nacional de Tucumán - Biología Vegetal. Notas de Clase 2012Documento99 páginasUniversidad Nacional de Tucumán - Biología Vegetal. Notas de Clase 2012Mariangeles GaviornoAún no hay calificaciones
- Manual de AnatomiaDocumento269 páginasManual de AnatomiaDaniel CarbajalAún no hay calificaciones
- Guía de BiologíaDocumento78 páginasGuía de BiologíaJuan Alejandro Quezada FariasAún no hay calificaciones
- Practica de CelulaDocumento8 páginasPractica de CelulaCamilo Andres Barreiro HerreraAún no hay calificaciones
- TALLER DE BIOLOGIA CELULAR - RespuestoDocumento7 páginasTALLER DE BIOLOGIA CELULAR - RespuestoValentina GallegoAún no hay calificaciones
- Plancha II PDFDocumento46 páginasPlancha II PDFJose Luis T SAún no hay calificaciones
- Celula CitoplasmaDocumento132 páginasCelula CitoplasmaSalma AlvarezAún no hay calificaciones
- DBIO1069 Biología Celular Endosomas y Tráfico Vesicular PDFDocumento18 páginasDBIO1069 Biología Celular Endosomas y Tráfico Vesicular PDFgustavo vargas diazAún no hay calificaciones
- La Célula Actividad en ClaseDocumento3 páginasLa Célula Actividad en Clasecarlos arrietaAún no hay calificaciones
- Banco de Preguntas de Biologia.Documento85 páginasBanco de Preguntas de Biologia.Cristian MamaniAún no hay calificaciones
- Manual de Laboratorio Biologia BLCDocumento50 páginasManual de Laboratorio Biologia BLCyestrada11Aún no hay calificaciones
- GT 2020 CN G6 Las CélulasDocumento11 páginasGT 2020 CN G6 Las CélulasJesus MejiaAún no hay calificaciones
- Manual de Patología General - Escuela Medicina PUC - Cap. 1 y 2Documento97 páginasManual de Patología General - Escuela Medicina PUC - Cap. 1 y 2Pablo Espinoza ContrerasAún no hay calificaciones
- Ensayo Sobre La Célula OnlineDocumento3 páginasEnsayo Sobre La Célula Onlineleidy885100% (2)
- Aparato de Golgi, Lisosomas, Vacuolas, PeroxisomasDocumento33 páginasAparato de Golgi, Lisosomas, Vacuolas, PeroxisomasAndi Dionisio100% (1)
- Tarea V Funciones de Cada Una de Las Estructuras CelularesDocumento2 páginasTarea V Funciones de Cada Una de Las Estructuras CelularesEddy Liranzo FañaAún no hay calificaciones
- BancodepreguntashistoDocumento30 páginasBancodepreguntashistoVan NanaAún no hay calificaciones
- El Sistema de Endomembranas - pptpara2010.Pptcap.VDocumento138 páginasEl Sistema de Endomembranas - pptpara2010.Pptcap.VlalitotAún no hay calificaciones
- Tema 1 - La Organización Celular de Los Ssvv.Documento92 páginasTema 1 - La Organización Celular de Los Ssvv.Señas IosuAún no hay calificaciones