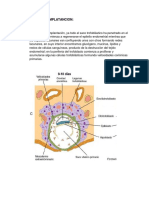
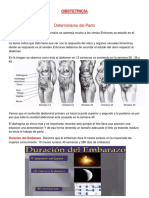
También podría gustarte
- Fecundación PDFDocumento4 páginasFecundación PDFAndrés PachónAún no hay calificaciones
- Cómo se produce la fecundación humana paso a pasoDocumento4 páginasCómo se produce la fecundación humana paso a pasostephany tamayoAún no hay calificaciones
- La FecundaciónDocumento3 páginasLa FecundaciónjuberAún no hay calificaciones
- FECUNDACIÓNDocumento3 páginasFECUNDACIÓNEstrellita CrisantoAún no hay calificaciones
- FECUNDACIÓNDocumento4 páginasFECUNDACIÓNAlyelis MarcanoAún no hay calificaciones
- Embriología: Fecundación, formación del blastocisto e implantaciónDocumento9 páginasEmbriología: Fecundación, formación del blastocisto e implantaciónLluvia CalderonAún no hay calificaciones
- FECUNDACION PlacentaDocumento13 páginasFECUNDACION Placentanathaly torresAún no hay calificaciones
- Fecundación. Capas Germinales, Ejes Del Cuerpo. Notocorda y MesodermoDocumento7 páginasFecundación. Capas Germinales, Ejes Del Cuerpo. Notocorda y MesodermoAradya GrajalesAún no hay calificaciones
- Proceso de Fecundación Del ÓvuloDocumento5 páginasProceso de Fecundación Del Óvulolucero martinezAún no hay calificaciones
- Fecundacion CtaDocumento3 páginasFecundacion CtaPiero OsccoAún no hay calificaciones
- Primera Semana de GestaciónDocumento6 páginasPrimera Semana de GestaciónBrenda Itzel Martinez AgustinAún no hay calificaciones
- Embriología - FecundaciónDocumento7 páginasEmbriología - Fecundaciónfariab11Aún no hay calificaciones
- Histología (Tema 2) (Embriologia)Documento21 páginasHistología (Tema 2) (Embriologia)Nathalia CasanovaAún no hay calificaciones
- Penetración de La Corona RadiadaDocumento2 páginasPenetración de La Corona RadiadaRicardojosue Gonzalez AcuñaAún no hay calificaciones
- FECUNDACIÓNDocumento6 páginasFECUNDACIÓNFrida SofíaAún no hay calificaciones
- 9-FecundacionDocumento4 páginas9-Fecundacionri21002Aún no hay calificaciones
- ENSAYODocumento5 páginasENSAYOANGELICA DIONICIO HERNANDEZAún no hay calificaciones
- SEMANA 03 PRÁCTICA EMBRIOLOGÍA Reconocimiento de Estructuras Formadas Durante La Primera Semana Del DesarrolloDocumento8 páginasSEMANA 03 PRÁCTICA EMBRIOLOGÍA Reconocimiento de Estructuras Formadas Durante La Primera Semana Del DesarrolloDiana CruzAún no hay calificaciones
- FecundaciónDocumento4 páginasFecundaciónbriandaaliciaangulo50% (2)
- Fecundación e Implantación Del EmbriónDocumento11 páginasFecundación e Implantación Del EmbriónFernanda GaxAún no hay calificaciones
- Fecundacion y Primera Se Mana de DesarrolloDocumento34 páginasFecundacion y Primera Se Mana de DesarrolloJuly CabariqueAún no hay calificaciones
- FecundaciónDocumento19 páginasFecundaciónBrandonn Montes HerediaAún no hay calificaciones
- 3 Concepcion y Desarrollo Del Huevo Enf.g.oDocumento7 páginas3 Concepcion y Desarrollo Del Huevo Enf.g.oGuizada SantAún no hay calificaciones
- FECUNDACIÓN Teoria PDFDocumento24 páginasFECUNDACIÓN Teoria PDFJoha LuceroAún no hay calificaciones
- Reproduccion Animal - 2Documento24 páginasReproduccion Animal - 2RUBEN MORALES BETANCURAún no hay calificaciones
- Etapa 1, 2 y 3Documento2 páginasEtapa 1, 2 y 3Luis TretoAún no hay calificaciones
- L Zárate - Actividad 3 (Fecundación y Desarrollo Embrionario) - 0Documento13 páginasL Zárate - Actividad 3 (Fecundación y Desarrollo Embrionario) - 0Lucy ZárateAún no hay calificaciones
- Concepcion HumanaDocumento4 páginasConcepcion HumanaNayelyAún no hay calificaciones
- FECUNDACIONDocumento8 páginasFECUNDACION70856141Aún no hay calificaciones
- Primeras Semanas Del Desarrollo EmbrionarioDocumento86 páginasPrimeras Semanas Del Desarrollo EmbrionarioIan RoblesAún no hay calificaciones
- 16 Desarrollo EmbrionarioDocumento39 páginas16 Desarrollo EmbrionarioSebastián CarrascalAún no hay calificaciones
- Objetivo PrincipalDocumento13 páginasObjetivo PrincipalErwin Yapuchura ApazaAún no hay calificaciones
- Fecundacion e ImplantacionDocumento14 páginasFecundacion e ImplantacionJessica BrocaAún no hay calificaciones
- ReproducciónDocumento3 páginasReproducciónelsy del rocio z. r.Aún no hay calificaciones
- Fecundación: Unión de óvulo y espermatozoideDocumento30 páginasFecundación: Unión de óvulo y espermatozoideLIA DANESSY JUAREZ NUÑEZAún no hay calificaciones
- Primera y Segunda Semana de DesarrolloDocumento8 páginasPrimera y Segunda Semana de DesarrolloAlen Gutierrez Guzman100% (1)
- Cuestionario EmbriologíaDocumento89 páginasCuestionario EmbriologíaDavenia SaldañaAún no hay calificaciones
- FECUNDACIONDocumento6 páginasFECUNDACIONDOLORES SANCHEZAún no hay calificaciones
- La Fecundación Paso A PasoDocumento2 páginasLa Fecundación Paso A PasoLizardo RodriguezAún no hay calificaciones
- Fecundación y segmentación: proceso de la fecundación y primeras divisiones celularesDocumento9 páginasFecundación y segmentación: proceso de la fecundación y primeras divisiones celularesNicole BetancourtAún no hay calificaciones
- FecundacionDocumento23 páginasFecundacionMarie ClavijoAún no hay calificaciones
- 23-Desarrollo EmbrionarioDocumento6 páginas23-Desarrollo EmbrionarioAgustin MontañaAún no hay calificaciones
- DREMRD - Desarrollo y GeneticaDocumento21 páginasDREMRD - Desarrollo y Geneticacamila garciaAún no hay calificaciones
- Resumen Embriologia UbaDocumento9 páginasResumen Embriologia UbaJonathan Romero AlvarezAún no hay calificaciones
- Linea Del Tiempo. Evolución Del EmbarazoDocumento13 páginasLinea Del Tiempo. Evolución Del EmbarazoPopoCourt67% (3)
- Fases de la fecundación e inicio del ciclo ováricoDocumento16 páginasFases de la fecundación e inicio del ciclo ováricoKaren AlmazánAún no hay calificaciones
- Fecundación 1Documento10 páginasFecundación 1DAINNEAún no hay calificaciones
- 2 Primera SemanaDocumento5 páginas2 Primera Semanaalisonarauz262003Aún no hay calificaciones
- Establecimiento de Gestacion en BovinoDocumento14 páginasEstablecimiento de Gestacion en BovinoVenezuelaGanadera.com100% (2)
- FECUNDACIÓN Tema 44 FinalDocumento15 páginasFECUNDACIÓN Tema 44 FinaltareasontimeyaAún no hay calificaciones
- FecundacionDocumento14 páginasFecundacionKatheryne DesiderioAún no hay calificaciones
- FecundacionDocumento23 páginasFecundacionalexnel100% (1)
- Primera Semana de Desarrollo Moore PDFDocumento6 páginasPrimera Semana de Desarrollo Moore PDFAdrian FreudAún no hay calificaciones
- FECUNDACIÓNDocumento2 páginasFECUNDACIÓNanderson colomaAún no hay calificaciones
- FecundaciónDocumento2 páginasFecundaciónAprendamos Juntos NiñosAún no hay calificaciones
- Etapas de La FecundacionDocumento3 páginasEtapas de La FecundacionDavid OlivaAún no hay calificaciones
- Periodo Embrionario (P1)Documento3 páginasPeriodo Embrionario (P1)Cindy LopezAún no hay calificaciones
- Embarazo, parto y lactancia del bebéDe EverandEmbarazo, parto y lactancia del bebéAún no hay calificaciones
- Giardia intestinalisDocumento7 páginasGiardia intestinalissalomendAún no hay calificaciones
- El derecho registralDocumento6 páginasEl derecho registralsalomendAún no hay calificaciones
- Que es concurso del delitoDocumento11 páginasQue es concurso del delitosalomendAún no hay calificaciones
- Cryptosporidium sppDocumento3 páginasCryptosporidium sppsalomendAún no hay calificaciones
- Definición del derecho ambientalDocumento3 páginasDefinición del derecho ambientalsalomendAún no hay calificaciones
- Cuándo Se Usan Los Alfabloqueantes AdrenergicosDocumento4 páginasCuándo Se Usan Los Alfabloqueantes AdrenergicossalomendAún no hay calificaciones
- CryptosporidiumDocumento9 páginasCryptosporidiumsalomendAún no hay calificaciones
- Enfermedades cardiovascularesDocumento12 páginasEnfermedades cardiovascularessalomendAún no hay calificaciones
- Ley PenalDocumento14 páginasLey PenalsalomendAún no hay calificaciones
- Comida ChileDocumento6 páginasComida ChilesalomendAún no hay calificaciones
- Las FuenteDocumento9 páginasLas FuentesalomendAún no hay calificaciones
- Regla de OroDocumento3 páginasRegla de OrosalomendAún no hay calificaciones
- Amebas ComensalesDocumento4 páginasAmebas ComensalessalomendAún no hay calificaciones
- Función Del Médico ForenseDocumento3 páginasFunción Del Médico ForensesalomendAún no hay calificaciones
- Topografía TorácicaDocumento10 páginasTopografía TorácicasalomendAún no hay calificaciones
- Ecología ParasitariaDocumento10 páginasEcología ParasitariasalomendAún no hay calificaciones
- FarmacodinamiaDocumento7 páginasFarmacodinamiasalomendAún no hay calificaciones
- AmbientalDocumento7 páginasAmbientalsalomendAún no hay calificaciones
- Definición de FarmacologíaDocumento12 páginasDefinición de FarmacologíasalomendAún no hay calificaciones
- ParvovirusDocumento5 páginasParvovirussalomendAún no hay calificaciones
- 5 Disco Germinativo BilaminarDocumento30 páginas5 Disco Germinativo BilaminarPaul SoriaAún no hay calificaciones
- GUIA Desarrollo Embrionario Embarzo PartoDocumento18 páginasGUIA Desarrollo Embrionario Embarzo PartoMariela MontemurroAún no hay calificaciones
- 1er Examen Todo EmbriologiaDocumento23 páginas1er Examen Todo EmbriologiaJorge Luis Gonzalez Mendoza100% (1)
- Semana 2 implatación: día 9 formación redes lacunaresDocumento1 páginaSemana 2 implatación: día 9 formación redes lacunaresInes MezaAún no hay calificaciones
- Clínica ginecológica UNLaRDocumento56 páginasClínica ginecológica UNLaRAlexis LavatelliAún no hay calificaciones
- FECUNDACIONDocumento24 páginasFECUNDACIONgabrielbbr100% (1)
- Taller Hormonas Sexuales y Ciclo MenstrualDocumento2 páginasTaller Hormonas Sexuales y Ciclo MenstrualLAURA VALENTINA ALARCON RIOSAún no hay calificaciones
- Segunda SemanaDocumento6 páginasSegunda SemanaggggAún no hay calificaciones
- Cambios Anatomofisiológicos en La Mujer GestanteDocumento14 páginasCambios Anatomofisiológicos en La Mujer GestanteMaria Belen LoorAún no hay calificaciones
- Desarrollo Embrionario - Etapas y Sus Características (Semana A Semana) - LifederDocumento15 páginasDesarrollo Embrionario - Etapas y Sus Características (Semana A Semana) - LifederMevis Paola Ruiz JuncoAún no hay calificaciones
- Semana 7 - Membranas Fetales y PlacentaDocumento71 páginasSemana 7 - Membranas Fetales y PlacentaAnthony Quispe Ramirez0% (1)
- Guia Desarrollo EmbrionarioDocumento7 páginasGuia Desarrollo EmbrionarioFredman Cruz YatAún no hay calificaciones
- Avo 13 BiologiaDocumento8 páginasAvo 13 BiologiaAgustina CandelariaAún no hay calificaciones
- Abortos EspontaneosDocumento28 páginasAbortos EspontaneosRafael Damaso SantosAún no hay calificaciones
- Guia EmbriologiaDocumento119 páginasGuia Embriologiaemely nuñez100% (1)
- Fases del desarrollo embrionario y fetal: de la fecundación a la implantaciónDocumento42 páginasFases del desarrollo embrionario y fetal: de la fecundación a la implantaciónPatricio andres Villarroel GonzalezAún no hay calificaciones
- Fecundación e implantación enDocumento6 páginasFecundación e implantación enHéctor Alfonso Cuevas CruzAún no hay calificaciones
- La Etapa Prenatal o Fase Pre... PsicologiaDocumento12 páginasLa Etapa Prenatal o Fase Pre... PsicologiaVictor TerronesAún no hay calificaciones
- Cuadernillo de Biología 3er. AñoDocumento18 páginasCuadernillo de Biología 3er. AñoAdriana Ondategui100% (1)
- Vaca RepetidoraDocumento108 páginasVaca Repetidoradaniel cruzadoAún no hay calificaciones
- Factor Uterino Infertilidad PDFDocumento11 páginasFactor Uterino Infertilidad PDFDenisse FurlanAún no hay calificaciones
- copia.docxDocumento67 páginascopia.docxSebastian Hernandez UknowAún no hay calificaciones
- Embriologia DiapositivasDocumento12 páginasEmbriologia DiapositivasEidi liseth Gómez perdomoAún no hay calificaciones
- Fecundación, Segmentación e ImplantaciónDocumento51 páginasFecundación, Segmentación e ImplantaciónYisselly Villalobos100% (1)
- Temas de ObstetriciaDocumento86 páginasTemas de ObstetriciaSalazar ArnoldAún no hay calificaciones
- Segunda Semana Del DesarrolloDocumento30 páginasSegunda Semana Del DesarrolloAmarau Flores RoqueAún no hay calificaciones
- Lista de Preguntas Obstetricia-2018Documento14 páginasLista de Preguntas Obstetricia-2018Alex OsorioAún no hay calificaciones
- Aborto ExpontaneoDocumento0 páginasAborto ExpontaneoGabriela AnguloAún no hay calificaciones
- Embrio Origen Del Desarrollo Humano .KeyDocumento201 páginasEmbrio Origen Del Desarrollo Humano .KeylauraAún no hay calificaciones
- Ultimo Caso (7966)Documento7 páginasUltimo Caso (7966)Mario LLerenaAún no hay calificaciones