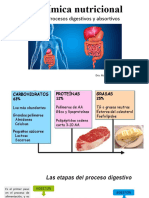
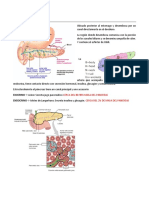
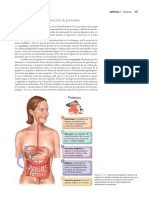
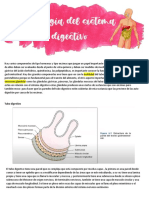
También podría gustarte
- Practica 02. Factores Que Modifican La Actividad EnzimaticaDocumento6 páginasPractica 02. Factores Que Modifican La Actividad EnzimaticaJeysson Cruzado Arteaga100% (1)
- Ross Jeffries - Frases Comadreja Patrones Palabras Trance Suavizantes - ESTEEEEDocumento21 páginasRoss Jeffries - Frases Comadreja Patrones Palabras Trance Suavizantes - ESTEEEEAriel Gamarra100% (1)
- Clase 10 Secreción Pancreatia y BiliarDocumento11 páginasClase 10 Secreción Pancreatia y BiliarLUCIA ANDREA LOPEZ GATICAAún no hay calificaciones
- Ficha S. Endocrino DuodenoDocumento2 páginasFicha S. Endocrino DuodenoMichelle Apunte RamosAún no hay calificaciones
- 2da Charla CirugiaDocumento4 páginas2da Charla CirugiaEstela QuirosAún no hay calificaciones
- Procesos de Digestion Unidad 2Documento118 páginasProcesos de Digestion Unidad 2evelyna.gomezAún no hay calificaciones
- Apuntes Pancreatitis IPChile 2019Documento9 páginasApuntes Pancreatitis IPChile 2019Constanza Silva CastroAún no hay calificaciones
- Fisio - Regulación de Secreción PancreáticaDocumento3 páginasFisio - Regulación de Secreción PancreáticaIsabela Cristina Verdugo SinibaldiAún no hay calificaciones
- Páncreas ExocrinoDocumento3 páginasPáncreas ExocrinoLiizbeth Aguilar LuisAún no hay calificaciones
- Clase 4 Digestión ProteicaDocumento17 páginasClase 4 Digestión ProteicaAguss FernandezAún no hay calificaciones
- Funciones Secretoras y DigestivasDocumento6 páginasFunciones Secretoras y DigestivasCarlos DelgadoAún no hay calificaciones
- PancreasDocumento10 páginasPancreasgiuliana fernandez campodonicoAún no hay calificaciones
- Composición Química de La Gastrina y Otras HormonasDocumento16 páginasComposición Química de La Gastrina y Otras HormonasHeidi PerezAún no hay calificaciones
- Revision PancreatitisDocumento13 páginasRevision PancreatitisCynthia LucasAún no hay calificaciones
- EstómagoDocumento13 páginasEstómagoSeba OrellanaAún no hay calificaciones
- Fisiologia Del Higado, Vesicula Biliar y Secreción PancreaticaDocumento10 páginasFisiologia Del Higado, Vesicula Biliar y Secreción PancreaticaMaria Fernanda Pingo FloresAún no hay calificaciones
- El Páncreas Fisiológicamente Esta Conformado Por Dos Tipos de Tejidos Que Determinan Su FunciónDocumento5 páginasEl Páncreas Fisiológicamente Esta Conformado Por Dos Tipos de Tejidos Que Determinan Su Funciónerika cabreraAún no hay calificaciones
- Pancreas ExocrinoDocumento10 páginasPancreas ExocrinoTamara Alejandra TorresAún no hay calificaciones
- Clase 3 Secreci N Pancre Tica Digesti N y Absorci N de Prote NasDocumento29 páginasClase 3 Secreci N Pancre Tica Digesti N y Absorci N de Prote NasPazAún no hay calificaciones
- Secreción GastrointestinalDocumento11 páginasSecreción GastrointestinalSergio ValdezAún no hay calificaciones
- Perspectivas de La Nutricion PROTEINASDocumento26 páginasPerspectivas de La Nutricion PROTEINASObed Efrain Castillo MartinezAún no hay calificaciones
- Subtema 3 Digestion Absorcion de ProteinasDocumento15 páginasSubtema 3 Digestion Absorcion de ProteinasXochitlAún no hay calificaciones
- Secreción PancreáticaDocumento7 páginasSecreción Pancreáticarociofigueroa.m130Aún no hay calificaciones
- Secreciones Del Intestino Delgado, Tampones, EnzimasDocumento4 páginasSecreciones Del Intestino Delgado, Tampones, EnzimasJuan D HuaychoAún no hay calificaciones
- Secreción PancreáticaDocumento5 páginasSecreción PancreáticaJuan Ernesto RojasAún no hay calificaciones
- Funciones de Las Principales Hormona y Otras Sustancias Del Aparato Digestivo Cruz - Campos - CarlaitzelDocumento4 páginasFunciones de Las Principales Hormona y Otras Sustancias Del Aparato Digestivo Cruz - Campos - CarlaitzelAmalia Colín CruzAún no hay calificaciones
- Fisiologia DigestivoDocumento20 páginasFisiologia DigestivoMaría de los Angeles PerezAún no hay calificaciones
- Sistema DigestivoDocumento1 páginaSistema DigestivoBritney GoicocheaAún no hay calificaciones
- Digestion Proteina Carbohidratos LipiidosDocumento4 páginasDigestion Proteina Carbohidratos LipiidosjosueAún no hay calificaciones
- Clase 22 Sistema Digestivo PDFDocumento26 páginasClase 22 Sistema Digestivo PDFQuimica TecnologiaAún no hay calificaciones
- Profundizacion ESIC ESFUNO Digestivo PDFDocumento27 páginasProfundizacion ESIC ESFUNO Digestivo PDFEvelyn FENFAún no hay calificaciones
- CLASE 6 Secreciones PancreáticasDocumento9 páginasCLASE 6 Secreciones PancreáticascopiastopoAún no hay calificaciones
- Control de La Secrecion PancreaticaDocumento8 páginasControl de La Secrecion PancreaticaSoany M. Alanoca CoaquiraAún no hay calificaciones
- Secreción SalivalDocumento5 páginasSecreción SalivalO'nell hernandezAún no hay calificaciones
- ++actividades. Hígado y Páncreas, AluDocumento4 páginas++actividades. Hígado y Páncreas, AluJordi JimAún no hay calificaciones
- Pancreas NOTAS MEDICASDocumento2 páginasPancreas NOTAS MEDICASLuis ValdezAún no hay calificaciones
- Función Bioquímica Del Intestino Grueso. Duarte ValiannaDocumento8 páginasFunción Bioquímica Del Intestino Grueso. Duarte ValiannaJandry Alexander Saltos SanzAún no hay calificaciones
- Digestion en Diferentes EspeciesDocumento11 páginasDigestion en Diferentes EspeciesHello CASHAún no hay calificaciones
- Colecistoquinina y SecretinaDocumento2 páginasColecistoquinina y SecretinaGeranni RondonAún no hay calificaciones
- Secreción PancreáticaDocumento12 páginasSecreción PancreáticaArleth LandivarAún no hay calificaciones
- Digestión en Intestino Delgado e Intestino Grueso. Absorción Digestiva.1Documento13 páginasDigestión en Intestino Delgado e Intestino Grueso. Absorción Digestiva.1Haide BlancoAún no hay calificaciones
- 01 Triptico La BilisDocumento2 páginas01 Triptico La BilisMARILUZ SAAVEDRA MAMANIAún no hay calificaciones
- Exposicion BioquimicaDocumento23 páginasExposicion Bioquimicamarco antonioAún no hay calificaciones
- Secreción PancreáticaDocumento3 páginasSecreción PancreáticaSAMIAún no hay calificaciones
- Pancreas ExocrinoDocumento21 páginasPancreas ExocrinoVALENTINA CAMILA RIQUELME CLAROSAún no hay calificaciones
- t4 Digestión y Absorción de Los AlimentosDocumento34 páginast4 Digestión y Absorción de Los Alimentoshbetancor2005Aún no hay calificaciones
- Presentacion Salud Del Cuerpo Humano Ilustracion RosaDocumento20 páginasPresentacion Salud Del Cuerpo Humano Ilustracion RosaAnthony GomezAún no hay calificaciones
- Unidad 4 Metabolismo Proteinas 1Documento18 páginasUnidad 4 Metabolismo Proteinas 1202203540Aún no hay calificaciones
- Capítulo 29 Berne y LevyDocumento7 páginasCapítulo 29 Berne y LevyHunHunahpuAún no hay calificaciones
- Digestion y MetabolismoDocumento17 páginasDigestion y MetabolismoNicole CaballeroAún no hay calificaciones
- Cuadro Fisiología Gástrica e IntestinalDocumento6 páginasCuadro Fisiología Gástrica e IntestinalAllison OportaAún no hay calificaciones
- Secrecion PancretaticaDocumento1 páginaSecrecion PancretaticaAri SantiagoAún no hay calificaciones
- DigestiónDocumento12 páginasDigestiónNatalia EstevesAún no hay calificaciones
- Fisiología Animal Informe 8Documento9 páginasFisiología Animal Informe 8desiree BOURONCLEAún no hay calificaciones
- Resumen Secreción IntestinalDocumento10 páginasResumen Secreción IntestinalAriana Elizabeth De Paz RodasAún no hay calificaciones
- Bioquimica Segunda SolemneDocumento21 páginasBioquimica Segunda SolemneRocio ContrerasAún no hay calificaciones
- Celulas de La Digestion 2Documento3 páginasCelulas de La Digestion 2JHUNIOR GEINER QUILICHE MARINAún no hay calificaciones
- PH y Acidez Gastrica Practica Lab 2 RochiDocumento3 páginasPH y Acidez Gastrica Practica Lab 2 Rochiuser1818Aún no hay calificaciones
- Digestion y Absorcion de NutrientesDocumento8 páginasDigestion y Absorcion de NutrientesJuanita AcostaAún no hay calificaciones
- PDF EstómagoDocumento1 páginaPDF Estómagoidk98017Aún no hay calificaciones
- Aparato Digestivo (Habilidades)Documento3 páginasAparato Digestivo (Habilidades)Natalia Fernandez LoeroAún no hay calificaciones
- La Epigenética Como Consecuencia de La Influencia Del Entorno Medioambiental y Materno en El Desarrollo EmbrionarioDocumento10 páginasLa Epigenética Como Consecuencia de La Influencia Del Entorno Medioambiental y Materno en El Desarrollo Embrionariocamila gonzalezAún no hay calificaciones
- Biologia-Un Recorrido Por Los Seres Vivos - de MicheliDocumento112 páginasBiologia-Un Recorrido Por Los Seres Vivos - de MicheliPablo Flores MainiAún no hay calificaciones
- Detalles de Los Distintos Tipos de RostroDocumento31 páginasDetalles de Los Distintos Tipos de Rostromariano desposorio100% (1)
- Tejido CartilaginosoDocumento5 páginasTejido CartilaginosoDaniela QuirozAún no hay calificaciones
- Bioprocesamiento de La LisinaDocumento4 páginasBioprocesamiento de La LisinaMaribi AranaAún no hay calificaciones
- Centro Escolar Colonia Tazumal 2Documento6 páginasCentro Escolar Colonia Tazumal 2Isai SolisAún no hay calificaciones
- Anatomia de Cabeza y CuelloDocumento12 páginasAnatomia de Cabeza y CuellovictorAún no hay calificaciones
- Secuencia Didactica Evolució 2Documento7 páginasSecuencia Didactica Evolució 2Luana CanzanoAún no hay calificaciones
- Xxix Olimpiada Metropolitana de BiologíaDocumento4 páginasXxix Olimpiada Metropolitana de BiologíaTani Mar EstrellaAún no hay calificaciones
- Clase 8 de OctubreDocumento3 páginasClase 8 de OctubreJuan SandovalAún no hay calificaciones
- PLAN MICRO S.11 2do. BGU BIOLOGIA-2Documento4 páginasPLAN MICRO S.11 2do. BGU BIOLOGIA-2ROSA100% (1)
- La Mariposa de Velmiro Ayala GaunaDocumento3 páginasLa Mariposa de Velmiro Ayala GaunaVanesa Pezzelatto100% (2)
- Plan 8 OctavoDocumento39 páginasPlan 8 OctavoFabian Andres Pineda BohorquezAún no hay calificaciones
- Ramas de La Ecologia y Ciencias RelacionadasDocumento4 páginasRamas de La Ecologia y Ciencias RelacionadasbammrsandmanAún no hay calificaciones
- Evaluacion Sumativa Ciencias Naturales Cuarto BasicoDocumento7 páginasEvaluacion Sumativa Ciencias Naturales Cuarto BasicoJAVIERA PENAAún no hay calificaciones
- Equilibrio Gibbs DonnanDocumento2 páginasEquilibrio Gibbs DonnanDanielAún no hay calificaciones
- Introducción A La Biologia VegetalDocumento20 páginasIntroducción A La Biologia VegetalVriel BolonAún no hay calificaciones
- Guia 1 Segundo Periodo Grado 4 - Ecosistemas ColombianosDocumento5 páginasGuia 1 Segundo Periodo Grado 4 - Ecosistemas ColombianosXIMENA CONDEAún no hay calificaciones
- Informe Sobre La MitocondriaDocumento6 páginasInforme Sobre La MitocondriaDenisse Lizeth Santiago Casaos100% (1)
- UNIDAD N 1 Ecolo - Docx2Documento5 páginasUNIDAD N 1 Ecolo - Docx2Brenda LimaAún no hay calificaciones
- Unidad 2Documento42 páginasUnidad 2Pedro RománAún no hay calificaciones
- Epitelio - LáminasDocumento27 páginasEpitelio - LáminasBárbara LeiteAún no hay calificaciones
- Facultad de Ciencias MédicasDocumento2 páginasFacultad de Ciencias MédicasJulia MacherettAún no hay calificaciones
- Boletín INIA #037 PDFDocumento110 páginasBoletín INIA #037 PDFdiegodiegodiego1Aún no hay calificaciones
- Secreciones y LíquidosDocumento51 páginasSecreciones y Líquidosnuedus corporativoAún no hay calificaciones
- Botánica - Tema 08Documento31 páginasBotánica - Tema 08Perez BalcarceAún no hay calificaciones
- T. 2.1 Cuestiones Selectividad La Materia VivaDocumento2 páginasT. 2.1 Cuestiones Selectividad La Materia VivaMarta Montero SanchezAún no hay calificaciones
- Crucigrama Tres - Maria Catalina Torres ArizaDocumento7 páginasCrucigrama Tres - Maria Catalina Torres ArizaCatalina TorresAún no hay calificaciones