También podría gustarte
- LIR Memorama Fisiologia PDFDocumento393 páginasLIR Memorama Fisiologia PDFPepita Sar100% (1)
- Biologia e Introduccion A La Biologia CelularDocumento128 páginasBiologia e Introduccion A La Biologia CelularAngie EstradaAún no hay calificaciones
- DecretoDocumento5 páginasDecretocielmarinaAún no hay calificaciones
- 2018 Primer Tomo A (Generalidades y Embrio)Documento34 páginas2018 Primer Tomo A (Generalidades y Embrio)GenesisCarreroRivasAún no hay calificaciones
- Angulos y Rectas ParalelasDocumento6 páginasAngulos y Rectas ParalelaswaldirAún no hay calificaciones
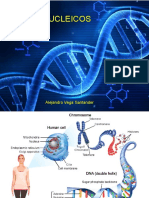
- 1-07 Acidos NucléicosDocumento55 páginas1-07 Acidos NucléicosDan EmpleoAún no hay calificaciones
- Estrategia de PreciosDocumento21 páginasEstrategia de PrecioswaldirAún no hay calificaciones
- Examen de BioquimicaDocumento1 páginaExamen de BioquimicaJose Eduardo Salazar SandovalAún no hay calificaciones
- Vitamina CDocumento20 páginasVitamina CPiero JuarezAún no hay calificaciones
- Ejercicios de Examen Quimica OrganicaDocumento6 páginasEjercicios de Examen Quimica OrganicaMiguel Eduardo Rangel CorderoAún no hay calificaciones
- El Psicópata Una Mente Amoral Tras La Máscara de La CorduraDocumento32 páginasEl Psicópata Una Mente Amoral Tras La Máscara de La CorduraLiseth Paola Vesga Pamplona100% (1)
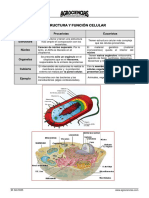
- TEMA 4 Estructura y Función Celular 2013 IDocumento9 páginasTEMA 4 Estructura y Función Celular 2013 I20110145Aún no hay calificaciones
- Manual de BioquímicaDocumento77 páginasManual de BioquímicaAldair PazAún no hay calificaciones
- Metabolismo de LípidosDocumento25 páginasMetabolismo de LípidosTobias Leonel PerezAún no hay calificaciones
- Comunicacion CelularDocumento25 páginasComunicacion CelularVictoria Sepulveda Salazar100% (1)
- Código Genético y Síntesis de ProteínasDocumento15 páginasCódigo Genético y Síntesis de ProteínasMariana100% (1)
- Apuntes Biologia Celular y TisularDocumento152 páginasApuntes Biologia Celular y TisularFatima Jerez50% (2)
- Introducciòn A La BioquìmicaDocumento14 páginasIntroducciòn A La BioquìmicaGeovanna K. OrtizAún no hay calificaciones
- Bioquimica Stryer 6 Edicion (Librosmedicospdf - Net)Documento1137 páginasBioquimica Stryer 6 Edicion (Librosmedicospdf - Net)Jhordan Castillo DiazAún no hay calificaciones
- Fisiología de Guyton - CompendioDocumento134 páginasFisiología de Guyton - CompendioAngel Alcivar “AAAS”Aún no hay calificaciones
- Receptores CelularesDocumento246 páginasReceptores Celularesclaudina bustamante herrera75% (4)
- INMUNOHISTOQUIMICADocumento35 páginasINMUNOHISTOQUIMICAbelinchon9667% (3)
- LehningerDocumento1283 páginasLehningerMelissa DiazAún no hay calificaciones
- BERNE y LEVY. FisiologíaDocumento11 páginasBERNE y LEVY. FisiologíaCarolina Barrio0% (1)
- Citología ExfoliativaDocumento98 páginasCitología ExfoliativaGisela ReyesAún no hay calificaciones
- Panamericana Anatomia Humana en Casos Clinicos 5a Edicion 856124Documento4 páginasPanamericana Anatomia Humana en Casos Clinicos 5a Edicion 856124Jeshue BarrazaAún no hay calificaciones
- Pentosas FosfatoDocumento39 páginasPentosas FosfatoKarolAún no hay calificaciones
- Manual de Bioquimica 5ta Edicion 3a Revisión314Documento140 páginasManual de Bioquimica 5ta Edicion 3a Revisión314juan100% (1)
- Bioquimica Devlin 4a EdicionDocumento621 páginasBioquimica Devlin 4a EdicionLuisAún no hay calificaciones
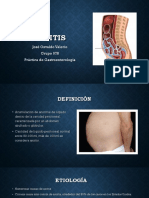
- AscitisDocumento13 páginasAscitisJose Osvaldo ValerioAún no hay calificaciones
- Uniones IntercelularesDocumento27 páginasUniones IntercelularesSusana Salas NuñezAún no hay calificaciones
- Uniones CelularesDocumento7 páginasUniones CelularesLeninLeo 18Aún no hay calificaciones
- DiureticosDocumento8 páginasDiureticoslucia mayaAún no hay calificaciones
- Examen de Bioquimica I UnidadDocumento3 páginasExamen de Bioquimica I UnidadRicardo Gustavo Iparraguirre Cortavitarte100% (1)
- Comorbilidad AspèrgerDocumento35 páginasComorbilidad AspèrgerCarfer CastilloAún no hay calificaciones
- Resumen Biologia Celular y MolecularDocumento9 páginasResumen Biologia Celular y MolecularMatilde Miranda100% (1)
- Cadena RespiratoriaDocumento2 páginasCadena RespiratoriaJesus RamirezAún no hay calificaciones
- Temas Examen de Bioquímica IIDocumento9 páginasTemas Examen de Bioquímica IIManuel Alejandro Iza GutierrezAún no hay calificaciones
- Copia Del Documento de EriceDocumento3 páginasCopia Del Documento de EriceIsa Linares de la CalAún no hay calificaciones
- Amortiguadores SanguíneosDocumento3 páginasAmortiguadores SanguíneosAna Alejandra Deluque CampoAún no hay calificaciones
- Diario de Laboratorio Bioquímica 1Documento10 páginasDiario de Laboratorio Bioquímica 1Jaime Wilfredo Garcia VieraAún no hay calificaciones
- ZOOLOGÍA DE VERTEBRADOS Clase 4 Peces IIDocumento47 páginasZOOLOGÍA DE VERTEBRADOS Clase 4 Peces IIJordan RuizAún no hay calificaciones
- Cap91011 AnestbloqbDocumento11 páginasCap91011 AnestbloqbDANIEL ALEJANDROAún no hay calificaciones
- Práctica Lab Medición de Líquidos in Vivo e in VitroDocumento8 páginasPráctica Lab Medición de Líquidos in Vivo e in VitroJesús MartínezAún no hay calificaciones
- Apreciación Del Poder Regulador de Las Soluciones Buffer y El Efecto de La ConcentraciónDocumento8 páginasApreciación Del Poder Regulador de Las Soluciones Buffer y El Efecto de La ConcentraciónXochitlAún no hay calificaciones
- Efecto de La Eritropoyetina Exógena, Sobre El Proceso de Eritropoyesis en Ratas Wistar MachoDocumento33 páginasEfecto de La Eritropoyetina Exógena, Sobre El Proceso de Eritropoyesis en Ratas Wistar Machovilchis_magu100% (9)
- DescargaDocumento6 páginasDescargaDavid PradoAún no hay calificaciones
- Síntesis Del Grupo HemoDocumento9 páginasSíntesis Del Grupo HemoDario TaimalAún no hay calificaciones
- Enzimas SéricasDocumento27 páginasEnzimas SéricasMariciell Larissa Gonzales TiconaAún no hay calificaciones
- Aplicaciones de La Formula de HendersonDocumento3 páginasAplicaciones de La Formula de HendersonClaudymar GonzalezAún no hay calificaciones
- Presentación Completa - Sem4Documento75 páginasPresentación Completa - Sem4Lucia InfinitivelifeAún no hay calificaciones
- Trastornos de LeucocitosDocumento54 páginasTrastornos de LeucocitosToni Pons AlbalatAún no hay calificaciones
- Solucion Examen Global de Bioquimica Octubre 2009Documento4 páginasSolucion Examen Global de Bioquimica Octubre 2009Prof. Víctor M. Vitoria100% (1)
- Tarea 3Documento3 páginasTarea 3Alfonso Mejia YoplaAún no hay calificaciones
- Cap 18 Glucolisis y Ox Del Piruvato HARPER Bioquimica Ilustrada 28a Ed-10ArchivoDocumento8 páginasCap 18 Glucolisis y Ox Del Piruvato HARPER Bioquimica Ilustrada 28a Ed-10ArchivoAbigail GajardoAún no hay calificaciones
- SalbutamolDocumento5 páginasSalbutamolTres Tesoros QiGong TaichiAún no hay calificaciones
- Evaluacion Multiple Choice. RespiratorioDocumento2 páginasEvaluacion Multiple Choice. RespiratorioDEBORAAún no hay calificaciones
- La Fisiopatología Como Base Fundamental Del Diagnóstico ClínicoDocumento3 páginasLa Fisiopatología Como Base Fundamental Del Diagnóstico ClínicoEnmanuel Guillen MartinezAún no hay calificaciones
- Moore Embriología 11a Ed.Documento260 páginasMoore Embriología 11a Ed.Gloria Adannelli MartínezAún no hay calificaciones
- Quimica de RigalliDocumento213 páginasQuimica de RigalliAdrian BrusaAún no hay calificaciones
- Inhibicion EnzimaticaDocumento17 páginasInhibicion EnzimaticaAlex Apaza MamaniAún no hay calificaciones
- Sistemas y Aparatos Del Cuerpo HumanoDocumento37 páginasSistemas y Aparatos Del Cuerpo HumanoGisela Pino PizáAún no hay calificaciones
- Bioquimica Tercera Edicion Libro de TextDocumento6 páginasBioquimica Tercera Edicion Libro de TextAracely PeñaAún no hay calificaciones
- Traducción: Transcripción, Traduc Ción Y Replicación CelularDocumento1 páginaTraducción: Transcripción, Traduc Ción Y Replicación CelularNtuys ChAún no hay calificaciones
- Victoria Sanchez 9C Estructura Del AdnDocumento8 páginasVictoria Sanchez 9C Estructura Del AdnVictoria Sanchez NietoAún no hay calificaciones
- Organizador Grafico Información GenéticaDocumento3 páginasOrganizador Grafico Información Genéticalquinonez9760Aún no hay calificaciones
- Procesos de Iniciación de Cadena Rezagada en La Replicación de ADN de EucariotasDocumento3 páginasProcesos de Iniciación de Cadena Rezagada en La Replicación de ADN de EucariotasAdriana Vanesa Tueros GuzmánAún no hay calificaciones
- Bipiav9g 10Documento9 páginasBipiav9g 10Milton Edward Robles ArquerosAún no hay calificaciones
- Trabajos en Altura 07-08Documento32 páginasTrabajos en Altura 07-08waldirAún no hay calificaciones
- 2021 Motivacion LaboralDocumento20 páginas2021 Motivacion LaboralwaldirAún no hay calificaciones
- Modulo 1 - Tema 2. Tipos de Modelización Con LS-DYNA I PDFDocumento6 páginasModulo 1 - Tema 2. Tipos de Modelización Con LS-DYNA I PDFwaldirAún no hay calificaciones
- Nectar de LimonDocumento35 páginasNectar de LimonwaldirAún no hay calificaciones
- Marco Conceptual de ImportacionesDocumento54 páginasMarco Conceptual de ImportacioneswaldirAún no hay calificaciones
- Integracion EconomicaDocumento6 páginasIntegracion EconomicawaldirAún no hay calificaciones
- Contrato de Agencias de AduanasDocumento6 páginasContrato de Agencias de AduanaswaldirAún no hay calificaciones
- Anemia HemoliticaDocumento15 páginasAnemia HemoliticaMaría Alejandra Gutiérrez TorresAún no hay calificaciones
- Caso Gen 6Documento3 páginasCaso Gen 6andrea100% (1)
- Efectos Teratogenicos de Fármacos AntiepilépticosDocumento29 páginasEfectos Teratogenicos de Fármacos AntiepilépticosAlejandro Sánchez100% (1)
- Citas Apa 1Documento3 páginasCitas Apa 1Dayana VergaraAún no hay calificaciones
- Teorias Biologicas Del EnvejecimientoDocumento5 páginasTeorias Biologicas Del EnvejecimientoAntonia J. Jara PozoAún no hay calificaciones
- Plan de Trabajo Mariano MelgarDocumento12 páginasPlan de Trabajo Mariano MelgarWilberth Corimanya GamarraAún no hay calificaciones
- Clase 10 Mutaciones CromosomicasDocumento49 páginasClase 10 Mutaciones Cromosomicasapi-3697648100% (2)
- 10 Evaluacion Preoperatoria Del Paciente Pediatrico MR MontenegroDocumento26 páginas10 Evaluacion Preoperatoria Del Paciente Pediatrico MR MontenegroMarcoMadueñoMejíaAún no hay calificaciones
- Acordeón AnatomíaDocumento3 páginasAcordeón AnatomíaGabbie HernándezAún no hay calificaciones
- Inteligencia Emocional 3ra TAVDocumento28 páginasInteligencia Emocional 3ra TAVVICTOR RICARDO DELGADO JULONAún no hay calificaciones
- El Fenómeno de RaynaudDocumento5 páginasEl Fenómeno de RaynaudIllıHerzen Von Eisllıl GerechtigkeitAún no hay calificaciones
- Macazal o Moringa OlefeiraDocumento3 páginasMacazal o Moringa OlefeiraConcepcion De Jesus GonzalezAún no hay calificaciones
- VPH Mitos y VerdadesDocumento29 páginasVPH Mitos y Verdadespaito2363Aún no hay calificaciones
- Taller PurinasDocumento9 páginasTaller PurinaslennyAún no hay calificaciones
- Fitopatologia Agricola Iievaluacion de La Severidad de La Antracnosis en Vainas de ArvejaDocumento3 páginasFitopatologia Agricola Iievaluacion de La Severidad de La Antracnosis en Vainas de ArvejaRomán DelgadoAún no hay calificaciones
- 2 AnomaliasgeneticasycongenitasDocumento124 páginas2 AnomaliasgeneticasycongenitasKAROL100% (1)
- M InmunologiaDocumento126 páginasM InmunologiaHilma Bermudez Gonzalez100% (1)
- APUNTES SOBRE MEDICINA TRADICIONAL CHINA - Marzo 2014 PDFDocumento11 páginasAPUNTES SOBRE MEDICINA TRADICIONAL CHINA - Marzo 2014 PDFMaricielo FloresAún no hay calificaciones
- Presentación CruzadaDocumento20 páginasPresentación CruzadaSus ArAún no hay calificaciones
- BioseguridadDocumento33 páginasBioseguridadJosé María MeloAún no hay calificaciones
- Geriatria (Por Competencias)Documento10 páginasGeriatria (Por Competencias)GABOVALAMEAún no hay calificaciones
- Ensayo TriptofanitoDocumento5 páginasEnsayo Triptofanitodany_2890% (1)
- CEFTRIAXONADocumento4 páginasCEFTRIAXONAzarate73Aún no hay calificaciones