También podría gustarte
- 4 Musculo EsqueleticoDocumento40 páginas4 Musculo EsqueleticoAngel LRAún no hay calificaciones
- Silabo de Fisiologia Medica - Obstetricia 2018Documento14 páginasSilabo de Fisiologia Medica - Obstetricia 2018LUIS RVAún no hay calificaciones
- Libro Guia PDF - Carlos Montero AyalaDocumento217 páginasLibro Guia PDF - Carlos Montero AyalaLeonardo Patrick Vega EstradaAún no hay calificaciones
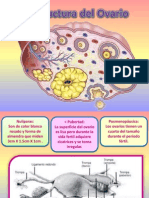
- OvarioDocumento11 páginasOvarioHanka Llosa PekovaAún no hay calificaciones
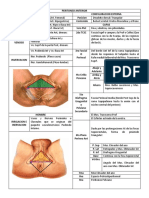
- Cuadro Sinoptico de Los Aparatos ReproductoresDocumento3 páginasCuadro Sinoptico de Los Aparatos ReproductoresMiguel angel Moreno alvaroAún no hay calificaciones
- Organelos Citoplasmaticos PDFDocumento41 páginasOrganelos Citoplasmaticos PDFMaria GorozabelAún no hay calificaciones
- Practica 6Documento7 páginasPractica 6Francesca Barreto CandiaAún no hay calificaciones
- CitocinasDocumento3 páginasCitocinasalan fernando100% (1)
- Caso Clinico N 6Documento6 páginasCaso Clinico N 6Johnny Sangalli ChuimaAún no hay calificaciones
- CASO CLÍNICO - EspondiloartrosisDocumento6 páginasCASO CLÍNICO - EspondiloartrosismanuelaAún no hay calificaciones
- Electrocardiograma DiapositivasDocumento15 páginasElectrocardiograma DiapositivasCristina Herrera100% (1)
- Histologia Animal 2017Documento54 páginasHistologia Animal 2017esstseess100% (1)
- Perineo Ant y PostDocumento3 páginasPerineo Ant y PostBristan Maraza PedemonteAún no hay calificaciones
- Estructura y maduración del espermatozoideDocumento3 páginasEstructura y maduración del espermatozoideJuanito PerezAún no hay calificaciones
- Taller de LeucogramaDocumento5 páginasTaller de Leucogramaoscar suarezAún no hay calificaciones
- Marco TeóricoDocumento24 páginasMarco TeóricoJohan AndradeAún no hay calificaciones
- Practica Sobre Sistema Nervioso - Anatomía ComparadaDocumento15 páginasPractica Sobre Sistema Nervioso - Anatomía ComparadaFLORES ALAYO JAVIER ALEJANDROAún no hay calificaciones
- Espacios PerifaringeosDocumento2 páginasEspacios PerifaringeosMaria PiñaAún no hay calificaciones
- Embriología Final 3Documento220 páginasEmbriología Final 3Alexandra & ElvisAún no hay calificaciones
- Introduccion A La Inmunologia PowerpointDocumento1 páginaIntroduccion A La Inmunologia PowerpointGabyGarciaAún no hay calificaciones
- Solucion Del Cuestionario 1 - MicroDocumento3 páginasSolucion Del Cuestionario 1 - MicroJosé Carlos Ccq100% (1)
- Morfofisiologia Embriologia - Caso Clinico 10Documento9 páginasMorfofisiologia Embriologia - Caso Clinico 10Joao Steve Raico RiveraAún no hay calificaciones
- Unidad I. Generalidades 2Documento15 páginasUnidad I. Generalidades 2Rorro8128363Aún no hay calificaciones
- Gonadas EmbriologiaDocumento1 páginaGonadas EmbriologiaMariana AltamiranoAún no hay calificaciones
- Trabajo de Investigación de Anatomía 4Documento7 páginasTrabajo de Investigación de Anatomía 4Jhasmani Cerezo100% (2)
- Embriologia Del Sistema Nervioso CentralDocumento5 páginasEmbriologia Del Sistema Nervioso Centralanayshizu100% (1)
- Sistema circulatorio región yugularDocumento12 páginasSistema circulatorio región yugularproagro12Aún no hay calificaciones
- Aparato reproductor masculinoDocumento11 páginasAparato reproductor masculinoAle HerreraAún no hay calificaciones
- Enfermedades Parasitarias Acantocefaliasis y NematodesDocumento6 páginasEnfermedades Parasitarias Acantocefaliasis y NematodesWilliams Rene Huanca CorimayoAún no hay calificaciones
- Diapositivas Musculos de Pelvis IIDocumento18 páginasDiapositivas Musculos de Pelvis IIjose luquezAún no hay calificaciones
- HematosisDocumento15 páginasHematosisDuzzter Garcia JimenezAún no hay calificaciones
- ProteinasDocumento6 páginasProteinasRodrigo LopezAún no hay calificaciones
- Sistematización Del Sistema Nervioso CentralDocumento8 páginasSistematización Del Sistema Nervioso CentralErmes ManuelAún no hay calificaciones
- Tejido EpitelialDocumento71 páginasTejido EpitelialLuis Castañeda ZavaletaAún no hay calificaciones
- Clase 4 - Sistema NeuroMuscularDocumento32 páginasClase 4 - Sistema NeuroMuscularAyelen SchwabAún no hay calificaciones
- LA CELULA Una Vision GeneralDocumento173 páginasLA CELULA Una Vision GeneralChristopher KruthoffAún no hay calificaciones
- CoenorusDocumento14 páginasCoenoruscelidaAún no hay calificaciones
- Sistemas reproductores masculino y femeninoDocumento9 páginasSistemas reproductores masculino y femeninojavillus100% (1)
- InmunofisiologiaDocumento61 páginasInmunofisiologiaCarlos Raul Solano Nizama0% (1)
- Genes y cromosomas, la información hereditariaDocumento15 páginasGenes y cromosomas, la información hereditariaMelissa Reyes LaraAún no hay calificaciones
- Frmacosantiparasitarios 130102190031 Phpapp01Documento51 páginasFrmacosantiparasitarios 130102190031 Phpapp01Wilmary Rodriguez MayorgaAún no hay calificaciones
- Fases de Los Neurotransmisores 2Documento1 páginaFases de Los Neurotransmisores 2María José Espinosa100% (1)
- Diagnóstico HelmintológicoDocumento45 páginasDiagnóstico HelmintológicoJoeMttCjAún no hay calificaciones
- FecundaciónDocumento3 páginasFecundaciónELIA MERITXELL JACOME MOROAún no hay calificaciones
- Manual de Prácticas Sistema Nervioso y Endócrino ENF02A 2022Documento26 páginasManual de Prácticas Sistema Nervioso y Endócrino ENF02A 2022Ruth Cobo RosalesAún no hay calificaciones
- Sistema tegumentario: funciones, partes, enfermedades y diversidadDocumento21 páginasSistema tegumentario: funciones, partes, enfermedades y diversidadJojary Alicea GarcíaAún no hay calificaciones
- Enfermedades Del Sistema NerviosoDocumento8 páginasEnfermedades Del Sistema NerviosoOscar Javier Ruiz OrtizAún no hay calificaciones
- Examen terminología médicaDocumento1 páginaExamen terminología médicaMike PalmaAún no hay calificaciones
- Cartel LCRDocumento1 páginaCartel LCRdariosoto100% (1)
- Cuestionario de Vascularizacion Del CorazónDocumento2 páginasCuestionario de Vascularizacion Del CorazónBrianda LopezAún no hay calificaciones
- EL APARATO REPRODUCTOR Caracteres Anatomicos Funcionales 1Documento59 páginasEL APARATO REPRODUCTOR Caracteres Anatomicos Funcionales 1Anonymous SyIBC4Os9Aún no hay calificaciones
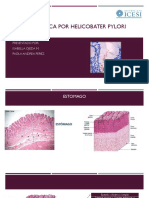
- Histologia: Úlcera Gástrica Por Helicobacter PyloriDocumento18 páginasHistologia: Úlcera Gástrica Por Helicobacter PyloriIsabella OjedaAún no hay calificaciones
- Patologias de La Glandula MamariaDocumento24 páginasPatologias de La Glandula MamariaÓscar David Ramírez MosqueraAún no hay calificaciones
- Patología Veterinaria I-Mvz-501 - Sem-09 - 2023-1Documento52 páginasPatología Veterinaria I-Mvz-501 - Sem-09 - 2023-1maricielo condori lopez100% (1)
- GASTRULACIONDocumento4 páginasGASTRULACIONLaura Daniela Leon MorenoAún no hay calificaciones
- Estructura piel animalesDocumento75 páginasEstructura piel animaleslilianapradaAún no hay calificaciones
- CLASE 6 Nematodos de Porcinos 2022Documento80 páginasCLASE 6 Nematodos de Porcinos 2022Gracia LoPeAún no hay calificaciones
- Adrenergicos TeoriaDocumento14 páginasAdrenergicos TeoriaDiana Oviedo LeonardoAún no hay calificaciones
- Sistema Nervioso Autonomo. Dra ZanuttiniDocumento80 páginasSistema Nervioso Autonomo. Dra ZanuttinirosaninamangovillegasAún no hay calificaciones
- Sistema nervioso autónomo: anatomía y funciónDocumento15 páginasSistema nervioso autónomo: anatomía y funciónAlejandra82% (11)
- Alimentación SaludableDocumento1 páginaAlimentación SaludableDavid GutierrezAún no hay calificaciones
- Monografia Craneo Fase 2Documento24 páginasMonografia Craneo Fase 2David GutierrezAún no hay calificaciones
- Biosafety and Biosecurity2Documento3 páginasBiosafety and Biosecurity2David GutierrezAún no hay calificaciones
- Caso Clinico 1Documento2 páginasCaso Clinico 1David GutierrezAún no hay calificaciones
- AcetilcolinaDocumento26 páginasAcetilcolinaAlejandro Jacob IturraAún no hay calificaciones
- Tejido MuscularDocumento39 páginasTejido MuscularnkmedunAún no hay calificaciones
- Tetanos Grupo BDocumento21 páginasTetanos Grupo BCarlos Barreto CervantesAún no hay calificaciones
- Farmacología. Generalidades Sistema Nervioso Autónomo: Dr. Julio José Chirinos Castillo Especialista en Medicina InternaDocumento45 páginasFarmacología. Generalidades Sistema Nervioso Autónomo: Dr. Julio José Chirinos Castillo Especialista en Medicina InternaSammy BenavidesAún no hay calificaciones
- Farmacología Especial (FARMACIA UCM)Documento246 páginasFarmacología Especial (FARMACIA UCM)Yolanda Herrero GonzalezAún no hay calificaciones
- FACULTAD DE CIENCIAS DE LA SALUD Pratica 5Documento3 páginasFACULTAD DE CIENCIAS DE LA SALUD Pratica 5Diego ChavezAún no hay calificaciones
- Drogas Anticolinesterasas-1Documento13 páginasDrogas Anticolinesterasas-1ANGELA LINARESAún no hay calificaciones
- Neurotransmisores y el impulso nerviosoDocumento12 páginasNeurotransmisores y el impulso nerviosoYatziri Camila Hernández JiménezAún no hay calificaciones
- Respuestas A Las 6 PreguntasDocumento8 páginasRespuestas A Las 6 PreguntasMichell OrdoñezAún no hay calificaciones
- Banco de Farmacologia Primer ParcialDocumento11 páginasBanco de Farmacologia Primer ParcialBoriz Rengifo GallegosAún no hay calificaciones
- Efectos Del Flúor en La Salud HumanaDocumento6 páginasEfectos Del Flúor en La Salud HumanayennycarolinaramirezAún no hay calificaciones
- Ordi Fisio I ResumenesDocumento70 páginasOrdi Fisio I ResumenesLupita PerezAún no hay calificaciones
- Incontinencia UrinariaDocumento66 páginasIncontinencia UrinariaEllys Macías Peralta100% (1)
- Folleto Clasificación Del Modo de Acción de Insecticidas y Acaricidas v.5 Ene19Documento28 páginasFolleto Clasificación Del Modo de Acción de Insecticidas y Acaricidas v.5 Ene19Gian Carlos Silva DiazAún no hay calificaciones
- Cuidados de Enfermeria Paciente Intoxicado - Tema 13 FinalDocumento72 páginasCuidados de Enfermeria Paciente Intoxicado - Tema 13 FinalLilyMolly100% (2)
- Intoxicacion Por OrganofosforadosDocumento10 páginasIntoxicacion Por OrganofosforadosJefferson CórdobaAún no hay calificaciones
- OrgonofosforadosDocumento7 páginasOrgonofosforadosKARELYS CHIRINOSAún no hay calificaciones
- Musculo LisoDocumento14 páginasMusculo LisoDiana OlivaresAún no hay calificaciones
- Clase AgonistasDocumento12 páginasClase AgonistasOctavo Semestre100% (1)
- Texto Farmacología IDocumento47 páginasTexto Farmacología ILexArcangel EstradaAún no hay calificaciones
- Neurotransmisores y NeuromoduladoresDocumento106 páginasNeurotransmisores y NeuromoduladoresJob Alan89% (9)
- Cerebro Triuno: Aprendizaje Integral con el CerebroDocumento22 páginasCerebro Triuno: Aprendizaje Integral con el CerebroLeonardo PuglieseAún no hay calificaciones
- Cap. 8 NeurotransmisorDocumento52 páginasCap. 8 NeurotransmisorjesusAún no hay calificaciones
- Farmacos Estimulantes ColinergicosDocumento14 páginasFarmacos Estimulantes ColinergicosSthilaire BelloAún no hay calificaciones
- 1er Paper - Organophosphate Intoxication1.en - EsDocumento10 páginas1er Paper - Organophosphate Intoxication1.en - EsIbeth NossaAún no hay calificaciones
- Uabp 3 SGR 2021Documento422 páginasUabp 3 SGR 2021Alejandro JacquatAún no hay calificaciones
- Vademecum Ecuador.Documento32 páginasVademecum Ecuador.Allan GabrielAún no hay calificaciones
- Fisiopatología de La Intoxicación Por OrganofosforadosDocumento2 páginasFisiopatología de La Intoxicación Por OrganofosforadosGaston Reyes PalomequeAún no hay calificaciones
- Trabajo Teórico de Unidad IIDocumento22 páginasTrabajo Teórico de Unidad IIThalia Aponte100% (1)
- Odontologia 2013 III ClaseDocumento60 páginasOdontologia 2013 III ClaseYeshenya Verònika CastilloAún no hay calificaciones
- Cómo conversar con cualquiera: Descubre cómo ser la persona más interesante del lugar y a destruir la ansiedad social permanentementeDe EverandCómo conversar con cualquiera: Descubre cómo ser la persona más interesante del lugar y a destruir la ansiedad social permanentementeCalificación: 4 de 5 estrellas4/5 (81)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Terapia cognitiva del comportamiento: Descubre cómo despejar tu cerebro con la TCC. Aprender a superar el miedo y la ansiedad, la depresiónDe EverandTerapia cognitiva del comportamiento: Descubre cómo despejar tu cerebro con la TCC. Aprender a superar el miedo y la ansiedad, la depresiónCalificación: 5 de 5 estrellas5/5 (3)
- Terapia cognitiva: Conceptos básicos y profundizaciónDe EverandTerapia cognitiva: Conceptos básicos y profundizaciónCalificación: 5 de 5 estrellas5/5 (1)
- Psicología y trastornos de los niños en la edad evolutiva: Qué son y cómo funcionanDe EverandPsicología y trastornos de los niños en la edad evolutiva: Qué son y cómo funcionanCalificación: 5 de 5 estrellas5/5 (1)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Dinámicas familiares a través de la vida de José: La túnica del PadreDe EverandDinámicas familiares a través de la vida de José: La túnica del PadreCalificación: 5 de 5 estrellas5/5 (2)
- Mente Organizada Cómo Pensar con Claridad y Hacer Todas Las Decisiones de Vida CorrectasDe EverandMente Organizada Cómo Pensar con Claridad y Hacer Todas Las Decisiones de Vida CorrectasCalificación: 4 de 5 estrellas4/5 (11)
- Neurociencia para vencer la depresión: La esprial ascendenteDe EverandNeurociencia para vencer la depresión: La esprial ascendenteCalificación: 4.5 de 5 estrellas4.5/5 (10)
- Guía para superar los pensamientos atemorizantes, obsesivos o inquietantes: Libérate de los pensamientos negativos con la terapia cognitivo conductual (CBT)De EverandGuía para superar los pensamientos atemorizantes, obsesivos o inquietantes: Libérate de los pensamientos negativos con la terapia cognitivo conductual (CBT)Calificación: 4 de 5 estrellas4/5 (25)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)
- Cómo estimular el cerebro del niño: 100 ejercicios para potenciar la concentración, la memoria y otras funciones ejecutivasDe EverandCómo estimular el cerebro del niño: 100 ejercicios para potenciar la concentración, la memoria y otras funciones ejecutivasCalificación: 4.5 de 5 estrellas4.5/5 (12)
- Genética general: Libro de textoDe EverandGenética general: Libro de textoCalificación: 4.5 de 5 estrellas4.5/5 (11)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- DMT: La molécula del espíritu: Las revolucionarias investigaciones de un médico sobre la biología de las experiencias místicas y cercanas a la muerteDe EverandDMT: La molécula del espíritu: Las revolucionarias investigaciones de un médico sobre la biología de las experiencias místicas y cercanas a la muerteCalificación: 4 de 5 estrellas4/5 (135)
- Bioneuroemoción: Un método para el bienestar emocionalDe EverandBioneuroemoción: Un método para el bienestar emocionalCalificación: 5 de 5 estrellas5/5 (4)
- La conciencia en el cerebro: Descifrando el enigma de cómo el cerebro elabora nuestros pensamientosDe EverandLa conciencia en el cerebro: Descifrando el enigma de cómo el cerebro elabora nuestros pensamientosCalificación: 2.5 de 5 estrellas2.5/5 (3)
- Cerebros rotos: Pacientes asombrosos que me enseñaron a vivirDe EverandCerebros rotos: Pacientes asombrosos que me enseñaron a vivirCalificación: 5 de 5 estrellas5/5 (3)
- Abrázame fuerte: Siete conversaciones para un amor duraderoDe EverandAbrázame fuerte: Siete conversaciones para un amor duraderoManu BerásteguiCalificación: 4.5 de 5 estrellas4.5/5 (13)
- El arte del descanso: Descubre el método para dormir bien y descansar mejorDe EverandEl arte del descanso: Descubre el método para dormir bien y descansar mejorCalificación: 4.5 de 5 estrellas4.5/5 (4)