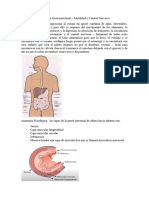
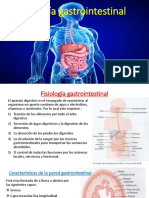
También podría gustarte
- Capitulos 63-64-65-66 PDFDocumento26 páginasCapitulos 63-64-65-66 PDFAdan Sánchez100% (1)
- MotilidadDocumento4 páginasMotilidadTALIA FERNANDA MUNIZ VASQUEZAún no hay calificaciones
- Fisiologia DigestivaDocumento54 páginasFisiologia DigestivaJosé MYAún no hay calificaciones
- Control Nervioso de La Función Gastrointestinal Sistema Nervioso EntéricoDocumento7 páginasControl Nervioso de La Función Gastrointestinal Sistema Nervioso EntéricoOscar Larios AyalaAún no hay calificaciones
- Fisiología: GastrointestinalDocumento8 páginasFisiología: GastrointestinalSarah Ddelgado100% (1)
- Resumen Guyton y Hall CAPITULO 62Documento8 páginasResumen Guyton y Hall CAPITULO 62Daniel Jaramillo Aponte100% (1)
- Cuestionario #04Documento9 páginasCuestionario #04Maria BautistaAún no hay calificaciones
- Principios Generales de La Función GastrointestinalDocumento8 páginasPrincipios Generales de La Función GastrointestinalFernando OrtizAún no hay calificaciones
- Capitulo 63Documento5 páginasCapitulo 63Thiellen Souza DiasAún no hay calificaciones
- Generalidades Del Si 266944 Downloadable 885932Documento9 páginasGeneralidades Del Si 266944 Downloadable 885932Jennifer NayelyAún no hay calificaciones
- Protocolo de Fisiolog DigestivaDocumento24 páginasProtocolo de Fisiolog DigestivaRuth Esther Nuñez MaldonadoAún no hay calificaciones
- Informe 1Documento14 páginasInforme 1Edwin Antonio Garcia VasquezAún no hay calificaciones
- Sistema Digestivo MonogástricosDocumento7 páginasSistema Digestivo MonogástricosFlavio CastañedaAún no hay calificaciones
- Plantilla para Artículos Normas APA Normasapa - Com 1Documento18 páginasPlantilla para Artículos Normas APA Normasapa - Com 1Vanesa NaranjoAún no hay calificaciones
- Principios Generales Función GastrointestinalDocumento6 páginasPrincipios Generales Función GastrointestinalAline Marian Moreira CordeiroAún no hay calificaciones
- Capitulo 62Documento7 páginasCapitulo 62Fernanda Maza100% (2)
- Principios Generales de La Motilidad GastrointestinalDocumento20 páginasPrincipios Generales de La Motilidad GastrointestinalElian ZoldyckAún no hay calificaciones
- Función GastrointestinalDocumento10 páginasFunción Gastrointestinalmaria alegriaAún no hay calificaciones
- Sistema DigestivoDocumento14 páginasSistema DigestivoDaniela Fernanda Ochoa SanchezAún no hay calificaciones
- Sistema Digestivo Material de LecturaDocumento23 páginasSistema Digestivo Material de Lecturacristhian AyalaAún no hay calificaciones
- Fisiologia Nutricion Clase 10 y 11Documento60 páginasFisiologia Nutricion Clase 10 y 11NAYELI LORENA HUARZAYA CUPIAún no hay calificaciones
- Aparato digestivo: funciones y estructuraDocumento57 páginasAparato digestivo: funciones y estructurageanniloAún no hay calificaciones
- Previo 6. ABSORCIÓN DE GLUCOSA Y PERISTALTISMODocumento15 páginasPrevio 6. ABSORCIÓN DE GLUCOSA Y PERISTALTISMOFernanda San MartinAún no hay calificaciones
- Microvasculatura Del Tubo DigestivoDocumento3 páginasMicrovasculatura Del Tubo DigestivoClaudioVillalobosAún no hay calificaciones
- Reflejos GastrointestinalesDocumento1 páginaReflejos GastrointestinalesValentina Arango100% (1)
- Fisiología Gastrointestinal .Documento25 páginasFisiología Gastrointestinal .wqzb4m9bwhAún no hay calificaciones
- Sistema Nervioso EntéricoDocumento9 páginasSistema Nervioso EntéricorosarioAún no hay calificaciones
- Fisio - Funcion Gastrointestinal-Resumen SPDocumento9 páginasFisio - Funcion Gastrointestinal-Resumen SPkkk jjjAún no hay calificaciones
- Fisiología GastrointestinalDocumento22 páginasFisiología GastrointestinalElías David Navarro ToscanoAún no hay calificaciones
- Apuntes Sistema Digestivo OKDocumento21 páginasApuntes Sistema Digestivo OKFernando LópezAún no hay calificaciones
- Cuestionario CaCuestionariop 62 GuytonDocumento4 páginasCuestionario CaCuestionariop 62 GuytonJosh GarridoAún no hay calificaciones
- Tubo Digestivo TextoDocumento46 páginasTubo Digestivo TextoRomero EspinozaAún no hay calificaciones
- Farmacos intestinales uterinosDocumento23 páginasFarmacos intestinales uterinosJimena Requejo BustamanteAún no hay calificaciones
- Digestivo FinalDocumento19 páginasDigestivo FinalJuan Martin Menes VegaAún no hay calificaciones
- Motilidad GastroIntestinal IDocumento6 páginasMotilidad GastroIntestinal IcopiastopoAún no hay calificaciones
- Fisiologiaaa EXPODocumento9 páginasFisiologiaaa EXPOnathalia.guamanAún no hay calificaciones
- CONCEPTOS GENERALES DijestivoDocumento9 páginasCONCEPTOS GENERALES DijestivoAlma GarciaAún no hay calificaciones
- MotilidadDocumento10 páginasMotilidadSabry El KokoAún no hay calificaciones
- Fisiología GIDocumento21 páginasFisiología GILenny E. Mejia JaquezAún no hay calificaciones
- Diapos FisioDocumento279 páginasDiapos FisioJohanna Cevallos100% (1)
- Sistema Nervioso en El Sistema DigestivoDocumento39 páginasSistema Nervioso en El Sistema DigestivoHarDmando LauraAún no hay calificaciones
- Digest IvoDocumento16 páginasDigest IvoCESAR DANIEL VELÁSQUEZ SANDOVALAún no hay calificaciones
- Fisiologia Gastrointestinal PDFDocumento14 páginasFisiologia Gastrointestinal PDFjaimusicAún no hay calificaciones
- Aparato digestivo: procesos y funcionesDocumento19 páginasAparato digestivo: procesos y funcioneskareli FloresAún no hay calificaciones
- Capitulo 63. PRINCIPIOS GENERALES DE LA FUNCION GASTROINTESTI0NAL. MOTILIDAD, CONTROL NERVIOSO Y CRICULACION SANGUINEADocumento11 páginasCapitulo 63. PRINCIPIOS GENERALES DE LA FUNCION GASTROINTESTI0NAL. MOTILIDAD, CONTROL NERVIOSO Y CRICULACION SANGUINEAFernandaAún no hay calificaciones
- Aparato DigestivoDocumento19 páginasAparato DigestivoAndreaAún no hay calificaciones
- CardiovascularDocumento10 páginasCardiovascularshavier009Aún no hay calificaciones
- Fisiologia GastrointestinalDocumento14 páginasFisiologia GastrointestinalPaola Carvajal QuinteroAún no hay calificaciones
- Principios Generales de La Funcion Gastroitestinal, Modalidad Control Nervioso y Circulacion SanguineaDocumento4 páginasPrincipios Generales de La Funcion Gastroitestinal, Modalidad Control Nervioso y Circulacion SanguineaYefferson Rafael huamani perezAún no hay calificaciones
- Apuntes de Fisiologia DigestivaDocumento45 páginasApuntes de Fisiologia DigestivaRonald Mamani Huañapaco50% (4)
- Sistema digestivo: funciones y componentesDocumento6 páginasSistema digestivo: funciones y componentesYorismel AlvarezAún no hay calificaciones
- UntitledDocumento232 páginasUntitledYuly medinaAún no hay calificaciones
- PERISTALTISMODocumento22 páginasPERISTALTISMOZeus LvAún no hay calificaciones
- Motilidad DigestivaDocumento16 páginasMotilidad DigestivatrumandemontesAún no hay calificaciones
- Sistema DigestivoDocumento38 páginasSistema DigestivoMarianela AscencioAún no hay calificaciones
- DigestivoDocumento26 páginasDigestivoAlberto Maldonado LarioAún no hay calificaciones
- Inervacion Del Tracto GiDocumento2 páginasInervacion Del Tracto Giacardona75Aún no hay calificaciones
- Documento Teórico Motilidad Del Intestino DelgadoDocumento11 páginasDocumento Teórico Motilidad Del Intestino DelgadoЗаира СуарезAún no hay calificaciones
- El Sistema Nervioso Entérico y NeurogastroenterologíaDocumento10 páginasEl Sistema Nervioso Entérico y NeurogastroenterologíaBrainArtVentasAún no hay calificaciones
- BIOSEGURIDADDocumento2 páginasBIOSEGURIDADDiego OrtegaAún no hay calificaciones
- InmunogenoDocumento1 páginaInmunogenoDiego OrtegaAún no hay calificaciones
- Alteraciones de TransmisionDocumento4 páginasAlteraciones de TransmisionDiego OrtegaAún no hay calificaciones
- Azul Marino Rojo Botiquín Desastre Natural PósterDocumento1 páginaAzul Marino Rojo Botiquín Desastre Natural PósterDiego OrtegaAún no hay calificaciones
- Presion ArterialDocumento2 páginasPresion ArterialDiego OrtegaAún no hay calificaciones
- Ascaris ToxocaraDocumento3 páginasAscaris ToxocaraDiego OrtegaAún no hay calificaciones
- ApuntesDocumento21 páginasApuntesDiego OrtegaAún no hay calificaciones
- ANENCEFALIADocumento5 páginasANENCEFALIADiego OrtegaAún no hay calificaciones
- Alteraciones de La PielDocumento1 páginaAlteraciones de La PielDiego OrtegaAún no hay calificaciones
- ANENCEFALIADocumento5 páginasANENCEFALIADiego OrtegaAún no hay calificaciones
- Diferenciar los cuatro tipos de huevos de Ascaris lumbricoides según su morfologíaDocumento4 páginasDiferenciar los cuatro tipos de huevos de Ascaris lumbricoides según su morfologíaLuisTorrezAún no hay calificaciones
- Ascaris Lumbricoides HelmintoDocumento4 páginasAscaris Lumbricoides HelmintoDiego OrtegaAún no hay calificaciones
- Ascaris Lumbricoides HelmintoDocumento4 páginasAscaris Lumbricoides HelmintoDiego OrtegaAún no hay calificaciones
- DefinicionDocumento2 páginasDefinicionDiego OrtegaAún no hay calificaciones
- TRIADASDocumento5 páginasTRIADASDiego OrtegaAún no hay calificaciones
- Tema Nro 3Documento7 páginasTema Nro 3Diego OrtegaAún no hay calificaciones
- Tema 13 ESTRUCTURA Y FUNCION DE LOS CROMOSOMASDocumento14 páginasTema 13 ESTRUCTURA Y FUNCION DE LOS CROMOSOMASDiego OrtegaAún no hay calificaciones
- ANENCEFALIADocumento5 páginasANENCEFALIADiego OrtegaAún no hay calificaciones
- CelulasDocumento1 páginaCelulasDiego OrtegaAún no hay calificaciones
- AsfixiaDocumento15 páginasAsfixiaSilvia MacorAún no hay calificaciones
- ANENCEFALIADocumento5 páginasANENCEFALIADiego OrtegaAún no hay calificaciones
- Anomalias Del Sistema DigestivoDocumento19 páginasAnomalias Del Sistema DigestivoDiego OrtegaAún no hay calificaciones
- Triada ecológica: definición, elementos y ejemplosDocumento20 páginasTriada ecológica: definición, elementos y ejemplosDiego OrtegaAún no hay calificaciones
- Metabolismo de Las ProteinasDocumento1 páginaMetabolismo de Las ProteinasDiego OrtegaAún no hay calificaciones
- Diferencias entre endemia, epidemia y pandemiaDocumento2 páginasDiferencias entre endemia, epidemia y pandemiaDiego OrtegaAún no hay calificaciones
- Alteraciones de La PielDocumento1 páginaAlteraciones de La PielDiego OrtegaAún no hay calificaciones
- EpidemiologiaDocumento1 páginaEpidemiologiaDiego OrtegaAún no hay calificaciones
- Anatomia ToraxDocumento6 páginasAnatomia ToraxSheila AbrahamAún no hay calificaciones
- Practica EcgDocumento7 páginasPractica EcgDiego OrtegaAún no hay calificaciones
- Manual botiquín: Guía para el uso y contenido de botiquines de primeros auxiliosDocumento20 páginasManual botiquín: Guía para el uso y contenido de botiquines de primeros auxiliosPilar Medrano MenaAún no hay calificaciones
- El Hombre Que Calculaba de Malba TahanDocumento5 páginasEl Hombre Que Calculaba de Malba TahanAngely Ana Bonfante HenaoAún no hay calificaciones
- Razas de Vacunos Con Aptitud para Producción de CarneDocumento1 páginaRazas de Vacunos Con Aptitud para Producción de Carneangel quispe tallaAún no hay calificaciones
- Higiene y Manipulacion de AlimentosDocumento7 páginasHigiene y Manipulacion de AlimentosGermanSepulvedaBolivarAún no hay calificaciones
- Pract. Lab. Analítico - BizcochueloDocumento7 páginasPract. Lab. Analítico - BizcochueloSofia GarciaAún no hay calificaciones
- Estrategia de Trade MarketingDocumento8 páginasEstrategia de Trade Marketingmatylokre LokreAún no hay calificaciones
- Caspar HauserDocumento295 páginasCaspar HauserclaribarbAún no hay calificaciones
- Manual de Gastronomia SaludableDocumento6 páginasManual de Gastronomia SaludableVIVIR MASAún no hay calificaciones
- Los refrescos en la cocteleríaDocumento57 páginasLos refrescos en la cocteleríaManuel EchëverryAún no hay calificaciones
- Bella y Bestia Sistema Internaciones y Sistema InglesDocumento7 páginasBella y Bestia Sistema Internaciones y Sistema InglesMaka OsAún no hay calificaciones
- Jugo de FrutasDocumento6 páginasJugo de Frutasvalen rojasAún no hay calificaciones
- PAN PAN PUM (1) FinallllDocumento2 páginasPAN PAN PUM (1) FinallllDaysi Duran FloresAún no hay calificaciones
- Del Arroyo.Documento225 páginasDel Arroyo.Adriana Dammiel Mora100% (3)
- Dieta KetoDocumento3 páginasDieta KetoPaola Alejandra Ortiz OregonAún no hay calificaciones
- Numeros RacionalesDocumento7 páginasNumeros RacionalesJuan Miguel Aquije IncaAún no hay calificaciones
- ACT 3.1 MetodologiaDocumento20 páginasACT 3.1 MetodologiaJimmy AyluardoAún no hay calificaciones
- 6184 File PRACTICA-04-RVDocumento3 páginas6184 File PRACTICA-04-RVItsel AlfaroAún no hay calificaciones
- Permeabilidad CelularDocumento31 páginasPermeabilidad CelularCamila AnahiAún no hay calificaciones
- Emulsiones cárnicas: factores de estabilidad y proceso de formaciónDocumento31 páginasEmulsiones cárnicas: factores de estabilidad y proceso de formaciónAnahí Auqui ArotomaAún no hay calificaciones
- Chefs ContemporáneosDocumento3 páginasChefs ContemporáneosfridaAún no hay calificaciones
- Coordinador 4to SecundariaDocumento16 páginasCoordinador 4to SecundariaElizabeth Cruzado RiosAún no hay calificaciones
- Arroz en Costa RicaDocumento22 páginasArroz en Costa RicaKheysi Nayelli Bello De los SantosAún no hay calificaciones
- Sleeping BeautyDocumento181 páginasSleeping BeautyJoss HeartAún no hay calificaciones
- Norma 204.007-2015Documento16 páginasNorma 204.007-2015Fernando GuevaraAún no hay calificaciones
- Manual de InstrucciónesDocumento12 páginasManual de Instrucciónesthegamer argAún no hay calificaciones
- Trabajo FinalDocumento21 páginasTrabajo FinalPaola SuarezAún no hay calificaciones
- Cartilla Abono OrganicoDocumento8 páginasCartilla Abono OrganicoNelsonBernalAlcalaAún no hay calificaciones
- Fetuccini A Lo AlfredoDocumento2 páginasFetuccini A Lo AlfredowalterAún no hay calificaciones
- Cerveceria Nacional Eeff Auditados 2019Documento115 páginasCerveceria Nacional Eeff Auditados 2019Alexander VasquezAún no hay calificaciones
- Soberania AlimentariaDocumento1 páginaSoberania AlimentariaAlexandra espinoza100% (1)
- El sentido de la lecheríaDocumento6 páginasEl sentido de la lecheríaValerie Juliana Luna Lopez0% (1)