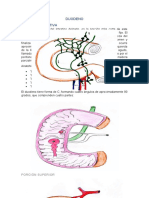
También podría gustarte
- Fisiología RenalDocumento6 páginasFisiología RenalMaria Fernanda julioAún no hay calificaciones
- Funciones Tubulos RenalesDocumento12 páginasFunciones Tubulos Renalesmaria riveroAún no hay calificaciones
- Función TubularDocumento17 páginasFunción TubularPilar Riquelme AguilarAún no hay calificaciones
- Tema 15, Fisiologia Renal - Funcion TubularDocumento18 páginasTema 15, Fisiologia Renal - Funcion TubularMaria Fernanda Pingo FloresAún no hay calificaciones
- Mecanismos de Reabsorción y Secreción TubularesDocumento10 páginasMecanismos de Reabsorción y Secreción TubularesMagdalena Astudillo RademacherAún no hay calificaciones
- Sustancias Sin Umbral y Función Del Epitelio TubularDocumento2 páginasSustancias Sin Umbral y Función Del Epitelio Tubulareduardo aguirreAún no hay calificaciones
- Fisiologìa de La Filtraciòn, Reabsorciòn y Secreciòn Tubular, QuizDocumento5 páginasFisiologìa de La Filtraciòn, Reabsorciòn y Secreciòn Tubular, QuizAndrew OspinaAún no hay calificaciones
- Manejo de Sales Por El Ri+ OnDocumento11 páginasManejo de Sales Por El Ri+ OnLenny Fanny Alvärädo100% (1)
- Mecanismos de La Formación de La Orin2Documento4 páginasMecanismos de La Formación de La Orin2majoAún no hay calificaciones
- Capitulo 28Documento11 páginasCapitulo 28Daniela Crystal GuillenAún no hay calificaciones
- Tema 13 FHDocumento9 páginasTema 13 FHMaria MingAún no hay calificaciones
- Paper RenalDocumento23 páginasPaper RenaljmvallerojasAún no hay calificaciones
- Manejo Renal de SodioDocumento12 páginasManejo Renal de SodioYvegly PaezAún no hay calificaciones
- Funcion TubularDocumento42 páginasFuncion TubularHector Marrufo75% (4)
- Mecanismo de La Formación de La OrinaDocumento3 páginasMecanismo de La Formación de La OrinaCarlos JaqueAún no hay calificaciones
- REABSORCIONDocumento9 páginasREABSORCIONEmmanuel Zótico Orellana JiménezAún no hay calificaciones
- Reabsorcion y Secrecion Tubular NuevoDocumento6 páginasReabsorcion y Secrecion Tubular NuevoLizbeth Paye ZamataAún no hay calificaciones
- Formación de La OrinaDocumento3 páginasFormación de La OrinaUYIAún no hay calificaciones
- GUIA 20 RiñonDocumento19 páginasGUIA 20 RiñonCamila GabrioneAún no hay calificaciones
- Función Renal IIDocumento8 páginasFunción Renal IILaura Carolina HerreraAún no hay calificaciones
- Mecanismos Que Regulan El Movimiento de Solutos.Documento3 páginasMecanismos Que Regulan El Movimiento de Solutos.Lisnep DelgadoAún no hay calificaciones
- Diureticos ResumenDocumento8 páginasDiureticos ResumenJulio100% (1)
- Clase IIDocumento18 páginasClase IIJose Ignacio Verdugo GutierrezAún no hay calificaciones
- Burton y RoseDocumento102 páginasBurton y RoseJem Severino VillacísAún no hay calificaciones
- Pruebas Alternativas en Examen General de OrinaDocumento8 páginasPruebas Alternativas en Examen General de OrinaYony HuancaAún no hay calificaciones
- Tema 8 Funciones TubularesDocumento14 páginasTema 8 Funciones TubularesvtedosoAún no hay calificaciones
- Tema 20 Reabsorcion y Secrecion Tubular PDFDocumento36 páginasTema 20 Reabsorcion y Secrecion Tubular PDFCielo Mar RiosAún no hay calificaciones
- Procesamiento Tubular Del Filtrado Glomerular Con Audio 22Documento26 páginasProcesamiento Tubular Del Filtrado Glomerular Con Audio 22ab21023Aún no hay calificaciones
- Fisiologia Renal 2Documento29 páginasFisiologia Renal 2ERWIN DEIVIS DIAZ BARRIOSAún no hay calificaciones
- Sistema Renal IIIDocumento13 páginasSistema Renal IIIVALENTINA CAMILA RIQUELME CLAROSAún no hay calificaciones
- Nefrona SamuelDocumento3 páginasNefrona SamuelSAMUEL ALEJANDRO MORENO MORENOAún no hay calificaciones
- Eq#2 Previo#5 IbarraCervantesMarianaDocumento7 páginasEq#2 Previo#5 IbarraCervantesMarianaMariana Ibarra CervantesAún no hay calificaciones
- Tema 15. Función Tubular. Mecanismos de Concentración y Dilución de Orina.Documento36 páginasTema 15. Función Tubular. Mecanismos de Concentración y Dilución de Orina.Pilar del Pino67% (3)
- Metabolismo de Compuestos Organicos PDFDocumento6 páginasMetabolismo de Compuestos Organicos PDFAgustin GamarnikAún no hay calificaciones
- CAPITULO 28 de GUYTONDocumento10 páginasCAPITULO 28 de GUYTONJhonier MaldonadoAún no hay calificaciones
- SEMINARIO 16 DiureticosDocumento28 páginasSEMINARIO 16 DiureticosyanettAún no hay calificaciones
- Fisiología de Producción de OrinaDocumento7 páginasFisiología de Producción de OrinaKarime LopezavilaAún no hay calificaciones
- Taller Fisiología Animal (Parcial 3)Documento20 páginasTaller Fisiología Animal (Parcial 3)LauraFernandezFrancoAún no hay calificaciones
- Reabsorción y Secreción TubularDocumento42 páginasReabsorción y Secreción TubularEvelyncita MendozaAún no hay calificaciones
- Practica 11 Fotosíntesis Agro 2022karenDocumento7 páginasPractica 11 Fotosíntesis Agro 2022karenKAREN ANABELLA VILCA MARTINEZAún no hay calificaciones
- Resumen DIURETICOSDocumento7 páginasResumen DIURETICOSDichany Gutierrez YepesAún no hay calificaciones
- Seminario Renal II - Sodio y PotasioDocumento65 páginasSeminario Renal II - Sodio y PotasioCayque OliveiraAún no hay calificaciones
- Orina LaboratorioDocumento12 páginasOrina LaboratorioDiego Cabanillas EspinozaAún no hay calificaciones
- Guyton Capitulo 28 Resumen CompendioDocumento21 páginasGuyton Capitulo 28 Resumen CompendioCristián Yax0% (1)
- Farmacos Diureticos 1 PracticaDocumento5 páginasFarmacos Diureticos 1 PracticaAlicia Benites Torres100% (2)
- Fisiopatologia de La DiarreaDocumento8 páginasFisiopatologia de La DiarreaGBlackmoreAún no hay calificaciones
- Anatomia y Fisiologia RenalDocumento6 páginasAnatomia y Fisiologia Renalvenustiano carranzaAún no hay calificaciones
- Cap 28 - REABSORCION Y SECRECION TUBULARDocumento19 páginasCap 28 - REABSORCION Y SECRECION TUBULARRodrigo LunaAún no hay calificaciones
- Practica de La Funcion RenalDocumento43 páginasPractica de La Funcion RenalEva Desireé Herrera RodriguezAún no hay calificaciones
- 10 - DiureticosDocumento6 páginas10 - DiureticosemiliorallAún no hay calificaciones
- Informe Sistema RenalDocumento8 páginasInforme Sistema RenalOscar PaulAún no hay calificaciones
- 12222222222222Documento6 páginas12222222222222Jesus Carrasco100% (1)
- Food Intake and Starvation Induces Metabolic Changes (1) EsDocumento37 páginasFood Intake and Starvation Induces Metabolic Changes (1) EscraffittoarAún no hay calificaciones
- Bioquímica de La Función RenalDocumento4 páginasBioquímica de La Función RenalaldoAún no hay calificaciones
- FUNCIÓN TUBULAR - CAPITULO 27 GuytonDocumento13 páginasFUNCIÓN TUBULAR - CAPITULO 27 GuytonSabrina SouzaAún no hay calificaciones
- 4.4 Reabsorcion y Secrecion TubularDocumento12 páginas4.4 Reabsorcion y Secrecion Tubularingrid gomezAún no hay calificaciones
- Guía de Estudios 7. Introducción Al Metabolismo. Leyva Vallejo Saúl Arturo-Grupo 1Documento9 páginasGuía de Estudios 7. Introducción Al Metabolismo. Leyva Vallejo Saúl Arturo-Grupo 1arturoleyva2001Aún no hay calificaciones
- Transporte Activo BombasDocumento12 páginasTransporte Activo BombasEleacith Johanán Mandon ArengasAún no hay calificaciones
- Fisiología Renal IIIDocumento10 páginasFisiología Renal IIIconstanzaalvarado0402Aún no hay calificaciones
- Triptico ApsDocumento2 páginasTriptico ApsJean Pierre Gomez ArotomaAún no hay calificaciones
- Jean SemioDocumento9 páginasJean SemioJean Pierre Gomez ArotomaAún no hay calificaciones
- Seminario SemioDocumento39 páginasSeminario SemioJean Pierre Gomez ArotomaAún no hay calificaciones
- Guia AnatomiaDocumento43 páginasGuia AnatomiaJean Pierre Gomez ArotomaAún no hay calificaciones
- Investigacion Formativa Inmunizacion Sarcov2Documento23 páginasInvestigacion Formativa Inmunizacion Sarcov2Jean Pierre Gomez ArotomaAún no hay calificaciones
- Identificación y Aislamiento de MicobacteriumDocumento4 páginasIdentificación y Aislamiento de MicobacteriumJean Pierre Gomez ArotomaAún no hay calificaciones
- Practica Numero°17Documento10 páginasPractica Numero°17Jean Pierre Gomez ArotomaAún no hay calificaciones
- Medicina Del BarrocoDocumento35 páginasMedicina Del BarrocoJean Pierre Gomez ArotomaAún no hay calificaciones
- Silabo MicroDocumento21 páginasSilabo MicroJean Pierre Gomez ArotomaAún no hay calificaciones
- Practica Numero°17Documento10 páginasPractica Numero°17Jean Pierre Gomez ArotomaAún no hay calificaciones
- Seram Recuerdo Anatómico Espacios Peritoneales y RetroperitonealesDocumento22 páginasSeram Recuerdo Anatómico Espacios Peritoneales y Retroperitonealesmikelww100% (3)
- Teoria 5Documento67 páginasTeoria 5Jean Pierre Gomez ArotomaAún no hay calificaciones
- PeritoneoDocumento18 páginasPeritoneoPaty MéndezAún no hay calificaciones
- Punto 4 ParasitologiaDocumento5 páginasPunto 4 ParasitologiaJean Pierre Gomez ArotomaAún no hay calificaciones
- Túbulo Contorneado ProximalDocumento11 páginasTúbulo Contorneado ProximalJean Pierre Gomez ArotomaAún no hay calificaciones
- Practica 15 - Identificacion y Aislamiento de Vibrio CholeraeDocumento8 páginasPractica 15 - Identificacion y Aislamiento de Vibrio CholeraeJean Pierre Gomez ArotomaAún no hay calificaciones
- Practica N°16 Resumen Del VideoDocumento6 páginasPractica N°16 Resumen Del VideoJean Pierre Gomez ArotomaAún no hay calificaciones
- RENANCENTISTADocumento30 páginasRENANCENTISTAJean Pierre Gomez ArotomaAún no hay calificaciones
- GiardialambliaDocumento16 páginasGiardialambliaJean Pierre Gomez ArotomaAún no hay calificaciones
- Practica 14Documento7 páginasPractica 14Jean Pierre Gomez ArotomaAún no hay calificaciones
- Universidad Privada San Juan Bautista Sac: Sílabo Por CompetenciaDocumento31 páginasUniversidad Privada San Juan Bautista Sac: Sílabo Por CompetenciaJean Pierre Gomez Arotoma100% (1)
- GiardialambliaDocumento16 páginasGiardialambliaJean Pierre Gomez ArotomaAún no hay calificaciones
- Prácticas N10Documento11 páginasPrácticas N10Jean Pierre Gomez ArotomaAún no hay calificaciones
- Resumen de Control de Riesgo Sanguineo Por Los Tejidos: Iv CicloDocumento2 páginasResumen de Control de Riesgo Sanguineo Por Los Tejidos: Iv CicloJean Pierre Gomez ArotomaAún no hay calificaciones
- Resumen de Control de Riesgo Sanguineo Por Los Tejidos: Iv CicloDocumento2 páginasResumen de Control de Riesgo Sanguineo Por Los Tejidos: Iv CicloJean Pierre Gomez ArotomaAún no hay calificaciones
- Practica 13Documento9 páginasPractica 13Jean Pierre Gomez ArotomaAún no hay calificaciones
- Resumen de Control de Riesgo Sanguineo Por Los Tejidos: Iv CicloDocumento2 páginasResumen de Control de Riesgo Sanguineo Por Los Tejidos: Iv CicloJean Pierre Gomez ArotomaAún no hay calificaciones
- Resumen de Control de Riesgo Sanguineo Por Los Tejidos: Iv CicloDocumento2 páginasResumen de Control de Riesgo Sanguineo Por Los Tejidos: Iv CicloJean Pierre Gomez ArotomaAún no hay calificaciones
- Resumen de Control de Riesgo Sanguineo Por Los Tejidos: Iv CicloDocumento2 páginasResumen de Control de Riesgo Sanguineo Por Los Tejidos: Iv CicloJean Pierre Gomez ArotomaAún no hay calificaciones
- Autoevaluable - Guias 11 y 12Documento7 páginasAutoevaluable - Guias 11 y 12valeAún no hay calificaciones
- Fisiologia GastroDocumento37 páginasFisiologia GastrothegaudiAún no hay calificaciones
- Nutrición Autótrofa y Nutrición HeterótrofaDocumento16 páginasNutrición Autótrofa y Nutrición HeterótrofaVimuZzaAún no hay calificaciones
- Tipos de ELISA, Elementos Que Lo ComponenDocumento4 páginasTipos de ELISA, Elementos Que Lo ComponenCarlos RondonAún no hay calificaciones
- Concept OsDocumento4 páginasConcept OsJuan Pablo RuizAún no hay calificaciones
- Vectores-Nociones Bibliotecas Parte 1 y 2-2021 PDFDocumento35 páginasVectores-Nociones Bibliotecas Parte 1 y 2-2021 PDFtito tatoAún no hay calificaciones
- Metabolismo Del Cáncer Mirando Hacia El Futuro (1) EspañolDocumento12 páginasMetabolismo Del Cáncer Mirando Hacia El Futuro (1) EspañolJOSE ANTONIO GOMEZ LA ROSAAún no hay calificaciones
- Protein AsDocumento30 páginasProtein AsJoselyn BaqueroAún no hay calificaciones
- BIOLOGIA-EXAMEN Junio 2012 PDFDocumento4 páginasBIOLOGIA-EXAMEN Junio 2012 PDFnereaabreuAún no hay calificaciones
- Guía de Actividad y Rubrica de Evaluación - Tarea 2 - Genetica y BiotecnologiaDocumento15 páginasGuía de Actividad y Rubrica de Evaluación - Tarea 2 - Genetica y BiotecnologiaLucia JaimesAún no hay calificaciones
- Articulo en InglesDocumento10 páginasArticulo en Inglesferney medinaAún no hay calificaciones
- Propiedades Catalíticas de La CatalasaDocumento2 páginasPropiedades Catalíticas de La CatalasaEduardo L. DíazAún no hay calificaciones
- Potencial de Acción en MesetaDocumento1 páginaPotencial de Acción en MesetaMaria CordovaAún no hay calificaciones
- Fructosa 6 FosfatoDocumento2 páginasFructosa 6 Fosfatoluis angel zegarra tuanamaAún no hay calificaciones
- Catalogue Amix Esp 2013 Web PDFDocumento56 páginasCatalogue Amix Esp 2013 Web PDFamixcolombiaAún no hay calificaciones
- Poe ProteinasDocumento22 páginasPoe Proteinasmanuel sosaAún no hay calificaciones
- Proliferacion CelularDocumento35 páginasProliferacion Celularapi-3777784100% (1)
- Guia Rutas MetabolicasDocumento3 páginasGuia Rutas MetabolicasRocioAún no hay calificaciones
- Folleto de Alimentación SaludableDocumento3 páginasFolleto de Alimentación SaludableOmar GomezAún no hay calificaciones
- Pap TesisDocumento155 páginasPap TesisDragan MalencicAún no hay calificaciones
- Ejercicio 16Documento3 páginasEjercicio 16KTAún no hay calificaciones
- Modo de Accion AntimicrobianaDocumento13 páginasModo de Accion AntimicrobianaRosarioFigueroaAún no hay calificaciones
- Virus HepatitisDocumento15 páginasVirus HepatitisLuis MartinezAún no hay calificaciones
- Clase de LipoproteinasDocumento26 páginasClase de LipoproteinasLuzvelia NúñezAún no hay calificaciones
- Infecciones Líticas ViroDocumento6 páginasInfecciones Líticas ViroLaura Cárdenas100% (1)
- TG Colesterol y Glucosa.Documento5 páginasTG Colesterol y Glucosa.doriz trejoAún no hay calificaciones
- Deteccion de OXIGENODocumento11 páginasDeteccion de OXIGENORenzo Ramos MolinaAún no hay calificaciones
- 2022.unidad 2 Lipidos - ActividadesDocumento4 páginas2022.unidad 2 Lipidos - ActividadesSofia CotrerasAún no hay calificaciones
- Deficiencias Del Metabolismo de Purinas y AsDocumento27 páginasDeficiencias Del Metabolismo de Purinas y Asmiguel angel100% (2)
- Los Acidos NucleicosDocumento9 páginasLos Acidos NucleicoscalosraAún no hay calificaciones
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- La revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaDe EverandLa revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaCalificación: 5 de 5 estrellas5/5 (203)
- Zensorialmente : Dejá que tu cuerpo sea tu cerebroDe EverandZensorialmente : Dejá que tu cuerpo sea tu cerebroAún no hay calificaciones
- Homo antecessor: El nacimiento de una especieDe EverandHomo antecessor: El nacimiento de una especieCalificación: 5 de 5 estrellas5/5 (1)
- Las Cinco Leyes Biológicas De La Naturaleza: La Nueva Medicina (Color Edition) SpanishDe EverandLas Cinco Leyes Biológicas De La Naturaleza: La Nueva Medicina (Color Edition) SpanishCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Disciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreDe EverandDisciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreCalificación: 5 de 5 estrellas5/5 (3)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- El péndulo de sanación: Péndulo hebreo. Investigación y sistematización de la técnicaDe EverandEl péndulo de sanación: Péndulo hebreo. Investigación y sistematización de la técnicaCalificación: 4.5 de 5 estrellas4.5/5 (27)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- La vibración de las ondas cerebrales: Recuperar el ritmo de una vida saludable y felizDe EverandLa vibración de las ondas cerebrales: Recuperar el ritmo de una vida saludable y felizCalificación: 5 de 5 estrellas5/5 (7)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Ikigai: Los secretos orientales para la longevidad explicados con el cine y la cultura popDe EverandIkigai: Los secretos orientales para la longevidad explicados con el cine y la cultura popCalificación: 4 de 5 estrellas4/5 (5)
- SNC - Soporte neurocrítico: De la urgencia a la terapia intensivaDe EverandSNC - Soporte neurocrítico: De la urgencia a la terapia intensivaAún no hay calificaciones
- Neurociencia en la escuela: Guía amigable (sin bla bla) para entender cómo funciona el cerebro durante el aprendizajeDe EverandNeurociencia en la escuela: Guía amigable (sin bla bla) para entender cómo funciona el cerebro durante el aprendizajeCalificación: 5 de 5 estrellas5/5 (5)
- Liberación del trauma: Perdón y temblor es el caminoDe EverandLiberación del trauma: Perdón y temblor es el caminoCalificación: 4 de 5 estrellas4/5 (4)
- Magia con Bicarbonato de Sodio: Decenas de Remedios y Usos Caseros que te Ahorrarán Dinero y Tiempo Utilizando el Bicarbonato de SodioDe EverandMagia con Bicarbonato de Sodio: Decenas de Remedios y Usos Caseros que te Ahorrarán Dinero y Tiempo Utilizando el Bicarbonato de SodioCalificación: 4.5 de 5 estrellas4.5/5 (7)
- Cerebro y silencio: Las claves de la creatividad y la serenidadDe EverandCerebro y silencio: Las claves de la creatividad y la serenidadCalificación: 5 de 5 estrellas5/5 (2)
- Dieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasDe EverandDieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasCalificación: 4 de 5 estrellas4/5 (9)
- La invasión de la pseudociencia: Claves para orientarse en un mundo en donde casi todo es posibleDe EverandLa invasión de la pseudociencia: Claves para orientarse en un mundo en donde casi todo es posibleCalificación: 4.5 de 5 estrellas4.5/5 (15)
- Anatomía & 100 estiramientos Esenciales (Color): Técnicas, beneficios, precauciones, consejos, tablas de series, dolenciasDe EverandAnatomía & 100 estiramientos Esenciales (Color): Técnicas, beneficios, precauciones, consejos, tablas de series, dolenciasCalificación: 4.5 de 5 estrellas4.5/5 (21)