También podría gustarte
- Sensaciones somáticas clasificaciónDocumento9 páginasSensaciones somáticas clasificaciónMARIANA AILEN FLORES FLORESAún no hay calificaciones
- Sistema SomatosensorialDocumento50 páginasSistema Somatosensorialmariobar17636Aún no hay calificaciones
- Sistema Somato Sensorial WordDocumento3 páginasSistema Somato Sensorial WordEdwin Quispe ToledoAún no hay calificaciones
- Sistema Nervioso Sensitivo Álvaro GazabónDocumento10 páginasSistema Nervioso Sensitivo Álvaro Gazabónivanacaico15Aún no hay calificaciones
- Vias Somatosensoriales Unidad 3Documento9 páginasVias Somatosensoriales Unidad 3Lo BM100% (2)
- Bases Fisiopatológicas Del DolorDocumento32 páginasBases Fisiopatológicas Del DolorCamila Alejandra Velazquez LopezAún no hay calificaciones
- Monografia TactoDocumento21 páginasMonografia TactoDeedee Revilla0% (1)
- Sistema SomatosensorialDocumento17 páginasSistema SomatosensorialEvelyn Rocio RomeroAún no hay calificaciones
- SENSIBILIDAD HUMANADocumento7 páginasSENSIBILIDAD HUMANAAndres Lopez Diago100% (1)
- Sistematización sensitiva y sensorialDocumento18 páginasSistematización sensitiva y sensorialNicole GarcíaAún no hay calificaciones
- Apunte14. Vías AferentesDocumento22 páginasApunte14. Vías AferentesThomas JohnsonAún no hay calificaciones
- Trabajo de Investigación Sobre Sistema SomatosensorialDocumento16 páginasTrabajo de Investigación Sobre Sistema SomatosensorialAntonio LopezAún no hay calificaciones
- TP 10Documento7 páginasTP 10Rafael AlvaradoAún no hay calificaciones
- Receptores SensorialesDocumento4 páginasReceptores SensorialesLiliana Henao.Aún no hay calificaciones
- Sistemas_Sensitivos_Generales_Apunte_Kine (1)Documento8 páginasSistemas_Sensitivos_Generales_Apunte_Kine (1)Alex AllueAún no hay calificaciones
- Sistema nervioso central y transmisión sinápticaDocumento4 páginasSistema nervioso central y transmisión sinápticaGabriela GutierrezAún no hay calificaciones
- SomatoviseralDocumento9 páginasSomatoviseralanaAún no hay calificaciones
- Funcion Somatosensitiva, Dolor, Cefalea y Regulacion de La TemperaturaDocumento13 páginasFuncion Somatosensitiva, Dolor, Cefalea y Regulacion de La TemperaturaJhonatan MChAún no hay calificaciones
- El DolorDocumento8 páginasEl Dolorlourdesmarquez231Aún no hay calificaciones
- Expo TactoDocumento41 páginasExpo TactoMaría Lourdes Cruz SalasAún no hay calificaciones
- Receptores cutáneos: Mecanorreceptores detectan deformacionesDocumento3 páginasReceptores cutáneos: Mecanorreceptores detectan deformacionesYahaira GutierrezAún no hay calificaciones
- Tema 6Documento9 páginasTema 6DAURA MEDINA HERNÁNDEZAún no hay calificaciones
- NEUROFISIOLOGIADocumento20 páginasNEUROFISIOLOGIABruno Masci MiralAún no hay calificaciones
- Unidad 1 - Parte 2Documento4 páginasUnidad 1 - Parte 2Ruth Esther Nuñez MaldonadoAún no hay calificaciones
- Unidad Ii NeuroDocumento9 páginasUnidad Ii NeuroSara SaucedoAún no hay calificaciones
- Notas de NocicepciónDocumento9 páginasNotas de NocicepciónCarolquiAún no hay calificaciones
- Vía espinotalámica: sensibilidad térmica y dolorDocumento6 páginasVía espinotalámica: sensibilidad térmica y dolorGerardo FloresAún no hay calificaciones
- SensibilidadDocumento111 páginasSensibilidadOlimpo RiveraAún no hay calificaciones
- PowerpointDocumento22 páginasPowerpointgabriela juana nahir castilloAún no hay calificaciones
- Resumen Sensibilidad SomáticaDocumento5 páginasResumen Sensibilidad SomáticaUPsublimados San RafaelAún no hay calificaciones
- Some S TesiaDocumento5 páginasSome S TesiaPILAR MOLINOLOAún no hay calificaciones
- Sensibilidad SomestesicaDocumento5 páginasSensibilidad SomestesicaHR100% (2)
- Dolor y SomestesiaDocumento18 páginasDolor y SomestesiaGustavoAún no hay calificaciones
- Receptores de DolorDocumento4 páginasReceptores de DolorEl fisiatraAún no hay calificaciones
- Sensibilidad Psicofisiologia 2023Documento6 páginasSensibilidad Psicofisiologia 2023Tati JuradoAún no hay calificaciones
- Ley de La ProyecciónDocumento11 páginasLey de La ProyecciónKaren Hermelinda0% (1)
- Arco Reflejo CuestionarioDocumento8 páginasArco Reflejo CuestionarioUam-Arturo MichelenaAún no hay calificaciones
- DolorDocumento18 páginasDolorVivi CardonaAún no hay calificaciones
- Vias NerviosasDocumento21 páginasVias NerviosasRoxanaSantillanAún no hay calificaciones
- Fisiología de Los ReflejosDocumento10 páginasFisiología de Los ReflejosmiiaestefaniiaAún no hay calificaciones
- Monografia Grupo 3 - FinalDocumento31 páginasMonografia Grupo 3 - FinalAngela Daniela Ortiz ArceAún no hay calificaciones
- Fisiologia Lab. 4Documento6 páginasFisiologia Lab. 4keyrisAún no hay calificaciones
- Tarea 3 (Psicofisiologia)Documento5 páginasTarea 3 (Psicofisiologia)andrixsonAún no hay calificaciones
- 48Documento5 páginas48Jhony PradelaAún no hay calificaciones
- Nervioso - 0 (Datos) 1-2Documento8 páginasNervioso - 0 (Datos) 1-2Agustina VallejosAún no hay calificaciones
- Taller TalamoDocumento9 páginasTaller TalamoEsteban Balcero ZuluagaAún no hay calificaciones
- Free-T2 - El Dolor-Gulag-FreeDocumento9 páginasFree-T2 - El Dolor-Gulag-FreeCristina PereaAún no hay calificaciones
- Sistema SomatosensoriaDocumento19 páginasSistema SomatosensoriaLupita GutierrezAún no hay calificaciones
- Sensibilidad Somatica, Percepcion.Documento3 páginasSensibilidad Somatica, Percepcion.Sol MayoralAguilarAún no hay calificaciones
- Fisio Sistemas Tema 2Documento3 páginasFisio Sistemas Tema 2abrahamAún no hay calificaciones
- Tipos de Dolor. El Dolor Rápido Se Siente en Cuestión de 0,1 S Después de Haber Aplicado El Estímulo, Se DescribeDocumento6 páginasTipos de Dolor. El Dolor Rápido Se Siente en Cuestión de 0,1 S Después de Haber Aplicado El Estímulo, Se DescribeJhony PradelaAún no hay calificaciones
- Vias de ConducciónDocumento20 páginasVias de ConducciónRocio CandásAún no hay calificaciones
- SNC I - MecanorreceptoresDocumento14 páginasSNC I - Mecanorreceptoresfisiorazetti09100% (3)
- Sistema Nervioso II - A.Documento30 páginasSistema Nervioso II - A.̿T̿e̿o̿ M̿o̿n̿t̿a̿n̿o̿ ̿G̿a̿m̿e̿z̿Aún no hay calificaciones
- Neurofisiologia Ii y Iii 2022 1 CicloDocumento56 páginasNeurofisiologia Ii y Iii 2022 1 CicloEdgardo RamirezAún no hay calificaciones
- Fisiología Humana 1 - ReporteDocumento28 páginasFisiología Humana 1 - ReporteNallely DiazAún no hay calificaciones
- $RUVQVEUDocumento11 páginas$RUVQVEUKevin AlanísAún no hay calificaciones
- Tema 4 SENSIBILIDAD SOMATICADocumento59 páginasTema 4 SENSIBILIDAD SOMATICANarda RetamosoAún no hay calificaciones
- Vias Centrales Del DolorDocumento5 páginasVias Centrales Del DolorEdher Lima GarciaAún no hay calificaciones
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2De EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Aún no hay calificaciones
- Regulación de La Expresión Génica A Nivel de La TraducciónDocumento3 páginasRegulación de La Expresión Génica A Nivel de La TraducciónAfrica ManeiroAún no hay calificaciones
- Material Final I 126Documento126 páginasMaterial Final I 126Africa ManeiroAún no hay calificaciones
- Resumen Mega Piola Salud Bibliotk AlequintanaDocumento36 páginasResumen Mega Piola Salud Bibliotk AlequintanaArmando NoelAún no hay calificaciones
- Guia Tabaco 2020 PDFDocumento118 páginasGuia Tabaco 2020 PDFLucila López MoristAún no hay calificaciones
- Notas sobre palabras clave y resumen de textoDocumento2 páginasNotas sobre palabras clave y resumen de textoAfrica ManeiroAún no hay calificaciones
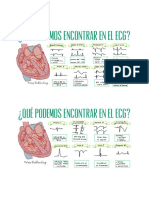
- Cambios ElectrocardiográficosDocumento1 páginaCambios ElectrocardiográficosAfrica ManeiroAún no hay calificaciones
- Mecanismos de Reparación Del ADNDocumento7 páginasMecanismos de Reparación Del ADNAfrica ManeiroAún no hay calificaciones
- Obesidad y Diabetes Fisiológia ResuDocumento5 páginasObesidad y Diabetes Fisiológia ResuAfrica ManeiroAún no hay calificaciones
- Regulación de la expresión génica en eucariotas: Cromatina y factores de transcripciónDocumento8 páginasRegulación de la expresión génica en eucariotas: Cromatina y factores de transcripciónAfrica ManeiroAún no hay calificaciones
- Resumen Ciclo Cardíaco, Acoplamiento Excito-Contráctil y Regulación Del Volumen SistólicoDocumento11 páginasResumen Ciclo Cardíaco, Acoplamiento Excito-Contráctil y Regulación Del Volumen SistólicoAfrica ManeiroAún no hay calificaciones
- Resumen de PatologíaBAQ - BhastaLinfomaNoHodgkinDocumento52 páginasResumen de PatologíaBAQ - BhastaLinfomaNoHodgkinAfrica ManeiroAún no hay calificaciones
- Fisiología RenalDocumento6 páginasFisiología RenalAfrica Maneiro0% (1)
- Fisiología respiratoria: intercambio gases, volúmenes pulmonaresDocumento6 páginasFisiología respiratoria: intercambio gases, volúmenes pulmonaresAfrica ManeiroAún no hay calificaciones
- Resumen Fisiología Celular Del Músculo Esquelético, Cardíaco y LisoDocumento15 páginasResumen Fisiología Celular Del Músculo Esquelético, Cardíaco y LisoAfrica ManeiroAún no hay calificaciones
- BIOESTADÍSTICADocumento2 páginasBIOESTADÍSTICAAfrica ManeiroAún no hay calificaciones
- Resumen Organización Del Sistema CardiovascularDocumento20 páginasResumen Organización Del Sistema CardiovascularAfrica ManeiroAún no hay calificaciones
- Radiaciones IonizantesDocumento1 páginaRadiaciones IonizantesAfrica ManeiroAún no hay calificaciones
- SueñoDocumento8 páginasSueñoAnonymous 5RDwUOxkmxAún no hay calificaciones
- Resumen HemodinámicaDocumento6 páginasResumen HemodinámicaAfrica ManeiroAún no hay calificaciones
- Zen y Artes Marciales - Taisen DeshimaruDocumento80 páginasZen y Artes Marciales - Taisen DeshimaruAfrica ManeiroAún no hay calificaciones
- La Historia Clinica Criminal PDFDocumento86 páginasLa Historia Clinica Criminal PDFEmily Nuñez0% (1)
- Ventilación Pulmonar y Difusión de GasesDocumento2 páginasVentilación Pulmonar y Difusión de GasesAfrica ManeiroAún no hay calificaciones
- Orexina y SueñoDocumento2 páginasOrexina y SueñoAfrica ManeiroAún no hay calificaciones
- Mi Religion - M. K. GandhiDocumento109 páginasMi Religion - M. K. GandhiRodrigo Gonzales CalleAún no hay calificaciones
- Mecanismos y Funcion Del SueñoDocumento6 páginasMecanismos y Funcion Del SueñoMaría Brú BeneditoAún no hay calificaciones
- Plan de Clase Secundaria 2014Documento45 páginasPlan de Clase Secundaria 2014cepc1312Aún no hay calificaciones
- P1 Morfo I 3era Edición 2009Documento10 páginasP1 Morfo I 3era Edición 2009JERISAIDAAún no hay calificaciones
- Esqueleto y Musculos 1º y 2º Eso 3ºev 10-11 PDFDocumento6 páginasEsqueleto y Musculos 1º y 2º Eso 3ºev 10-11 PDFsAún no hay calificaciones
- Prueba Ciencias Sistema LocomotorDocumento3 páginasPrueba Ciencias Sistema LocomotorCristina Palma Martinez100% (2)
- Calambres musculares: causas y minerales claveDocumento5 páginasCalambres musculares: causas y minerales claveIván Max PerezAún no hay calificaciones
- Desarrollo MotorDocumento62 páginasDesarrollo MotorLuz Bernal Romero100% (1)
- TEG Eduardo - Ricardo, FinalDocumento44 páginasTEG Eduardo - Ricardo, FinalRicardo HernandezAún no hay calificaciones
- Sistema MuscularDocumento9 páginasSistema MuscularSoudre MauroAún no hay calificaciones
- TEMA 26 FlexibilidadDocumento31 páginasTEMA 26 Flexibilidadalex_gil_9210Aún no hay calificaciones
- BIOLOGÍA - HISTOLOGÍA-Multimedia - Preparados-Para Alumnos y Prof.Documento158 páginasBIOLOGÍA - HISTOLOGÍA-Multimedia - Preparados-Para Alumnos y Prof.Cecilia MorenoAún no hay calificaciones
- 10 - ElectroestimulaciónDocumento23 páginas10 - ElectroestimulaciónEmi RiosAún no hay calificaciones
- LATARJETDocumento40 páginasLATARJETNoe MilyAún no hay calificaciones
- Exploración Neurológica CompletaDocumento13 páginasExploración Neurológica CompletaAlán Belman AndradeAún no hay calificaciones
- Educacion FisicaDocumento2 páginasEducacion FisicaAnderson taco alvisAún no hay calificaciones
- Traduccion Ultrasonido 6 en 1Documento14 páginasTraduccion Ultrasonido 6 en 1Alexa RM56% (18)
- Tecnicas de Enfermeria 45454 PDFDocumento18 páginasTecnicas de Enfermeria 45454 PDFLaura BecerraAún no hay calificaciones
- Resumen de TriptofanitoDocumento19 páginasResumen de TriptofanitoRicardo Blancas Castro100% (1)
- Hipotonía MuscularDocumento3 páginasHipotonía MuscularAdriana FlflAún no hay calificaciones
- AnatomiaDocumento296 páginasAnatomiaVitam NoelAún no hay calificaciones
- Morfología Del DiencefaloDocumento26 páginasMorfología Del DiencefaloJose TalaveraAún no hay calificaciones
- El Ultimo Plan para Perder Grasa - Daniel RomanDocumento19 páginasEl Ultimo Plan para Perder Grasa - Daniel RomanAfon AfonAún no hay calificaciones
- Practica NroDocumento4 páginasPractica NroAlex Mendoza PalominoAún no hay calificaciones
- BiofísicaDocumento17 páginasBiofísicaRuthAún no hay calificaciones
- Guía Generalidades AnatomiaDocumento16 páginasGuía Generalidades AnatomiaCarlos LlanosAún no hay calificaciones
- Contenido 3Documento20 páginasContenido 3Rodrigo EspinozaAún no hay calificaciones
- Posicionamiento Del en El Mercado Del Agua Mineral San MateoDocumento4 páginasPosicionamiento Del en El Mercado Del Agua Mineral San MateoKatherine CabreraAún no hay calificaciones
- Programa de Mejora de La Condición Física 3º EsoDocumento5 páginasPrograma de Mejora de La Condición Física 3º Esotrivas60Aún no hay calificaciones
- Aparato LocomotorDocumento11 páginasAparato Locomotoranatoniayfisilogia100% (1)
- Glosario de Terminos de Educacion FisicaDocumento10 páginasGlosario de Terminos de Educacion FisicaBiagna GrossoAún no hay calificaciones
- CerdoDocumento18 páginasCerdoJuan Linares PerezAún no hay calificaciones